The Circadian Clock—A Molecular Tool for Survival in Cyanobacteria


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
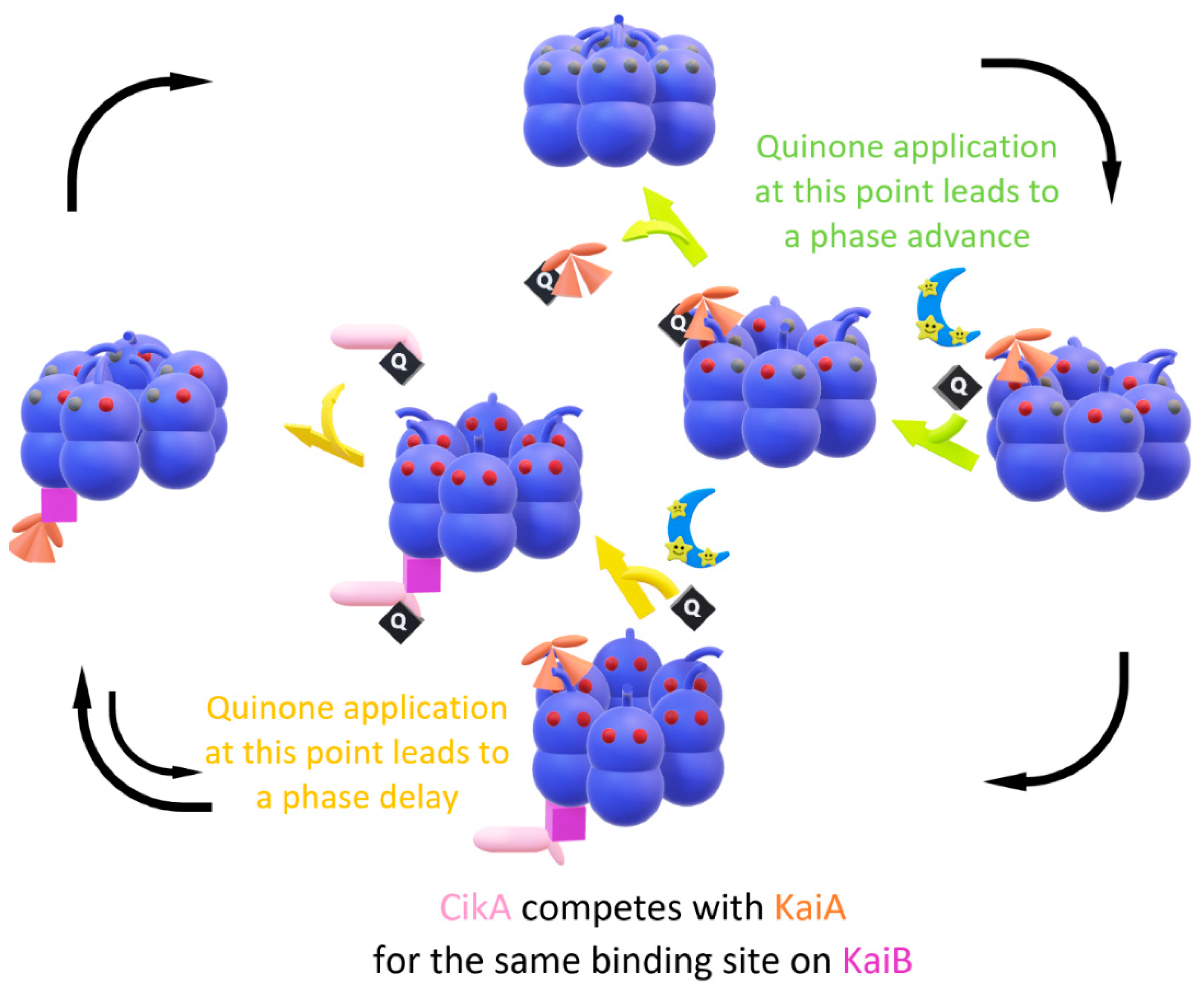{kind=link}
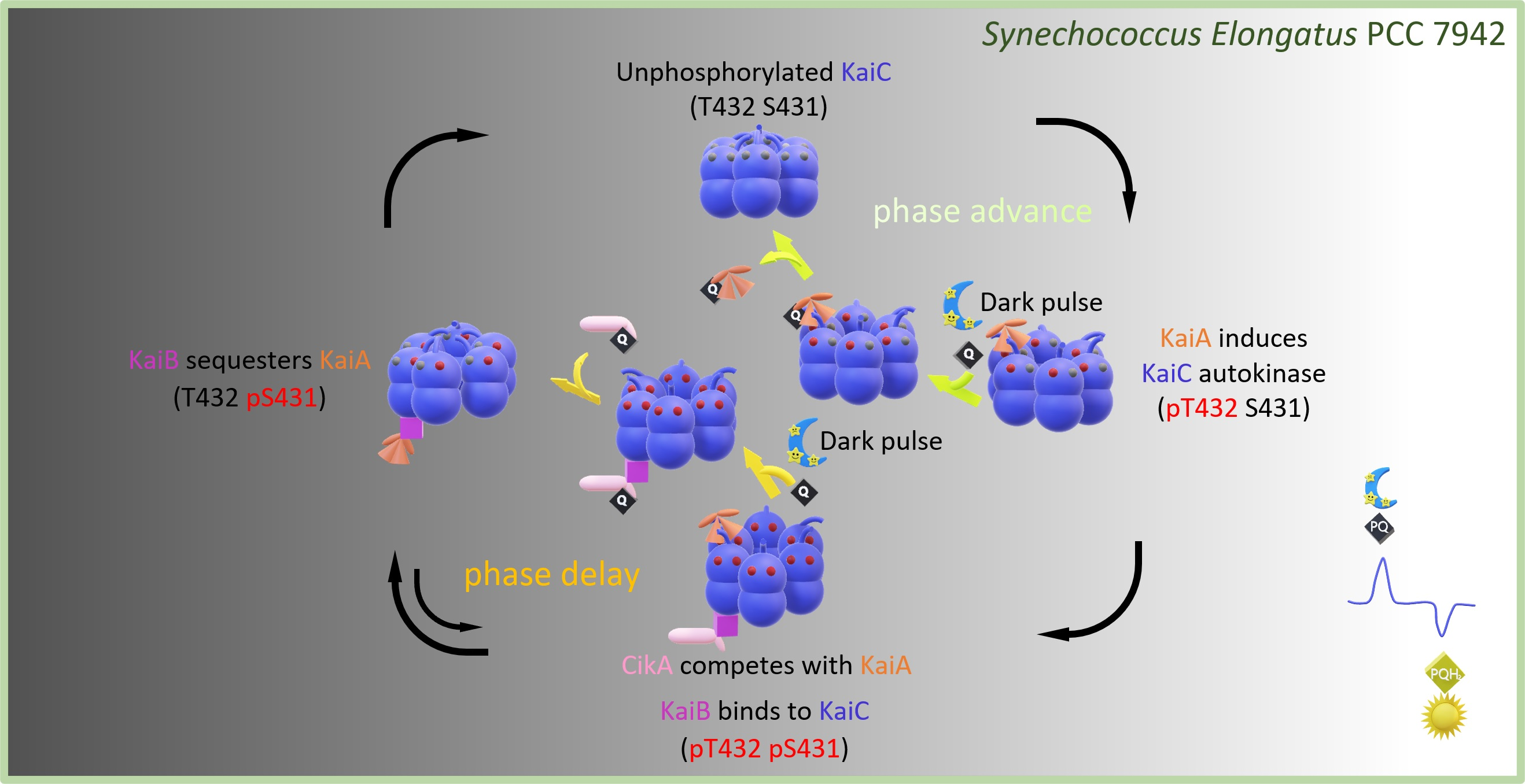
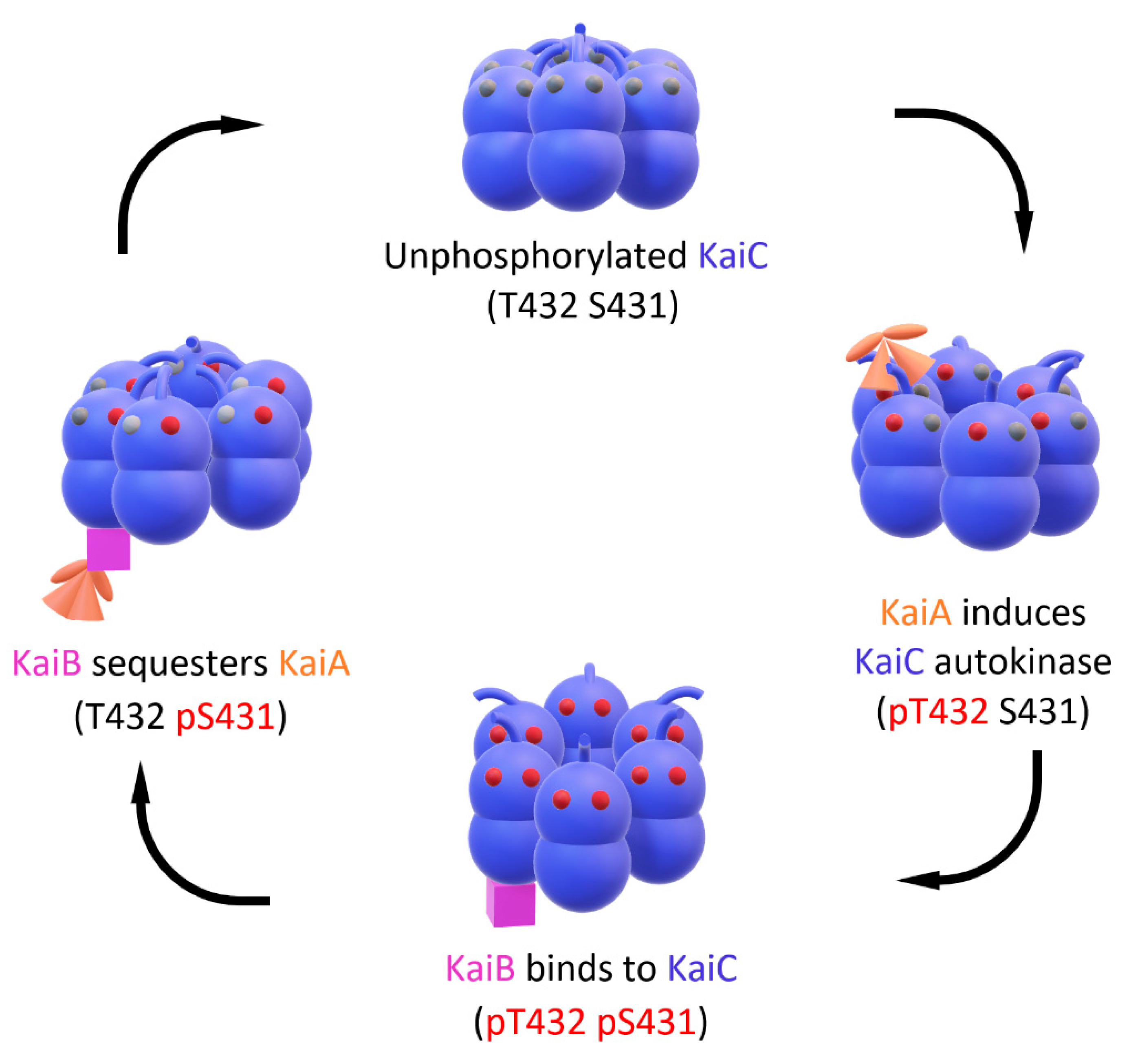
1. Introduction—The Free-Running Cyanobacterial Central Oscillator
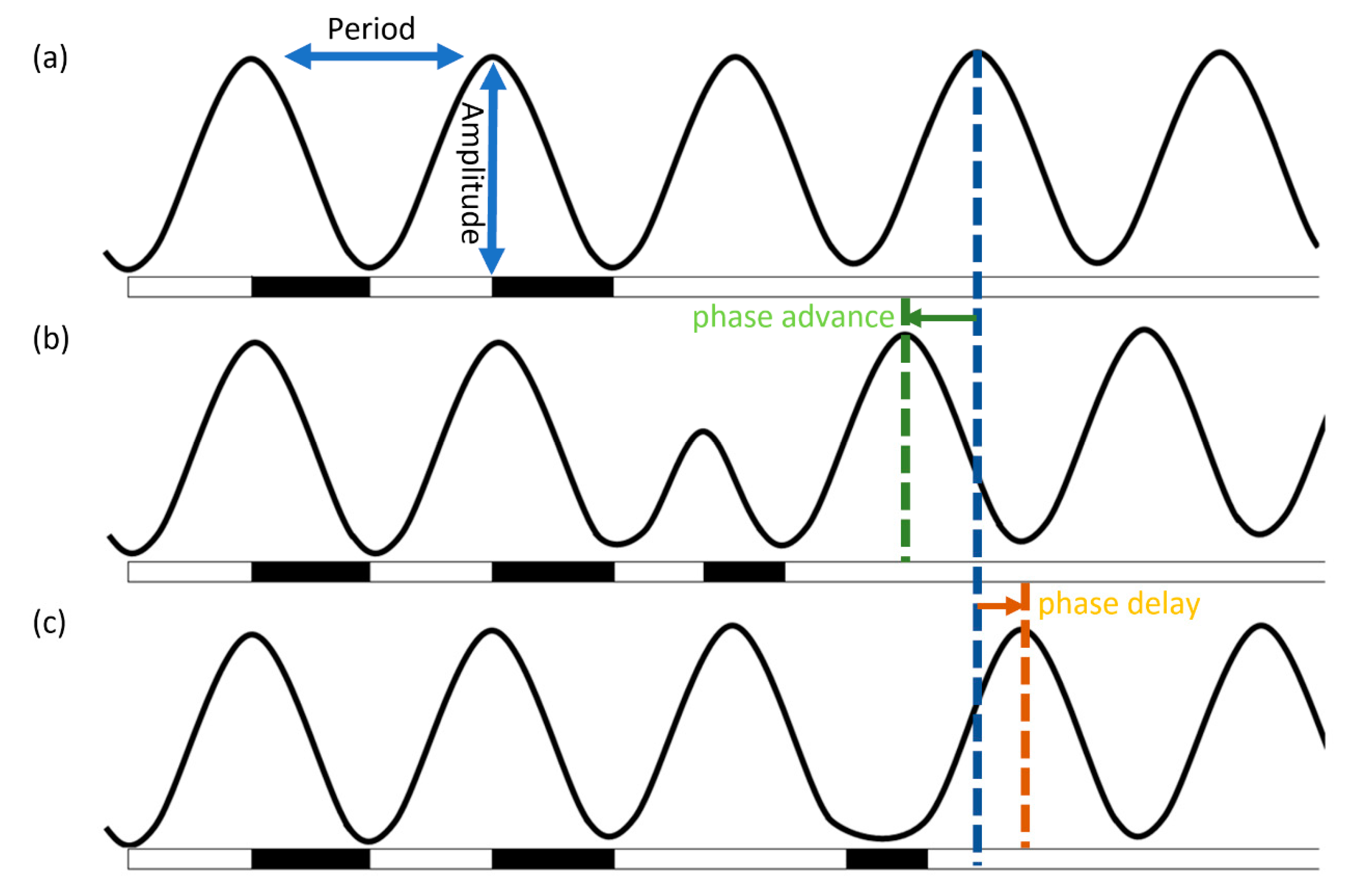
2. Entrainment through the Input Pathway
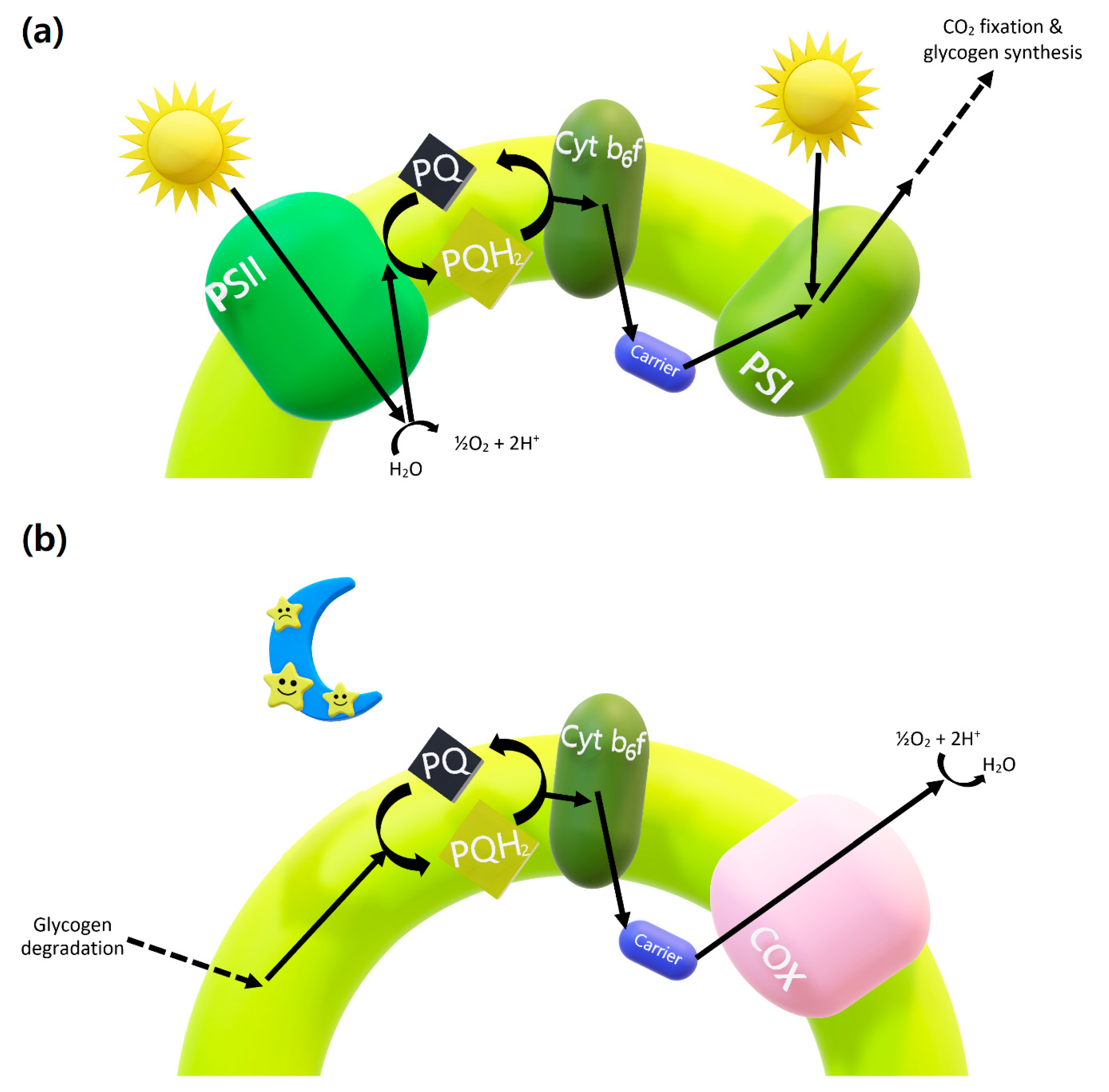
3. Physiological Rhythm through the Output Pathway
4. Timekeepers in Other Species of Cyanobacteria
5. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mullineaux, C.W. Co-existence of photosynthetic and respiratory activities in cyanobacterial thylakoid membranes. Biochim. Biophys. Acta 2014, 1837, 503–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puszynska, A.M.; O’Shea, E.K. Switching of metabolic programs in response to light availability is an essential function of the cyanobacterial circadian output pathway. eLife 2017, 6, e23210. [Google Scholar] [CrossRef] [PubMed]
- Markson, J.S.; Piechura, J.R.; Puszynska, A.M.; O’Shea, E.K. Circadian control of global gene expression by the cyanobacterial master regulator RpaA. Cell 2013, 155, 1396–1408. [Google Scholar] [CrossRef] [Green Version]
- Pattanayak, G.K.; Phong, C.; Rust, M.J. Rhythms in energy storage control the ability of the cyanobacterial circadian clock to reset. Curr. Biol. 2014, 24, 1934–1938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piechura, J.R.; Amarnath, K.; O’Shea, E.K. Natural changes in light interact with circadian regulation at promoters to control gene expression in cyanobacteria. eLife 2017, 6, e32032. [Google Scholar] [CrossRef] [PubMed]
- Schuurmans, R.M.; Schuurmans, J.M.; Bekker, M.; Kromkamp, J.C.; Matthijs, H.C.; Hellingwerf, K.J. The redox potential of the plastoquinone pool of the cyanobacterium Synechocystis species strain PCC 6803 is under strict homeostatic control. Plant Physiol. 2014, 165, 463–475. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.S.; Green, E.W.; Zhao, Y.; van Ooijen, G.; Olmedo, M.; Qin, X.; Xu, Y.; Pan, M.; Valekunja, U.K.; Feeney, K.A.; et al. Peroxiredoxins are conserved markers of circadian rhythms. Nature 2012, 485, 459–464. [Google Scholar] [CrossRef] [Green Version]
- Kondo, T.; Strayer, C.A.; Kulkarni, R.D.; Taylor, W.; Ishiura, M.; Golden, S.S.; Johnson, C.H. Circadian rhythms in prokaryotes: Luciferase as a reporter of circadian gene expression in cyanobacteria. Proc. Natl. Acad. Sci. USA 1993, 90, 5672–5676. [Google Scholar] [CrossRef] [Green Version]
- Ishiura, M.; Kutsuna, S.; Aoki, S.; Iwasaki, H.; Andersson, C.R.; Tanabe, A.; Golden, S.S.; Johnson, C.H.; Kondo, T. Expression of a gene cluster kaiABC as a circadian feedback process in cyanobacteria. Science 1998, 281, 1519–1523. [Google Scholar] [CrossRef]
- Pattanayek, R.; Wang, J.; Mori, T.; Xu, Y.; Johnson, C.H.; Egli, M. Visualizing a circadian clock protein: Crystal structure of KaiC and functional insights. Mol. Cell 2004, 15, 375–388. [Google Scholar] [CrossRef]
- Terauchi, K.; Kitayama, Y.; Nishiwaki, T.; Miwa, K.; Murayama, Y.; Oyama, T.; Kondo, T. ATPase activity of KaiC determines the basic timing for circadian clock of cyanobacteria. Proc. Natl. Acad. Sci. USA 2007, 104, 16377–16381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomita, J.; Nakajima, M.; Kondo, T.; Iwasaki, H. No transcription-translation feedback in circadian rhythm of KaiC phosphorylation. Science 2005, 307, 251–254. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, M.; Imai, K.; Ito, H.; Nishiwaki, T.; Murayama, Y.; Iwasaki, H.; Oyama, T.; Kondo, T. Reconstitution of circadian oscillation of cyanobacterial KaiC phosphorylation in vitro. Science 2005, 308, 414–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, Y.M.; Dias, C.; Diekman, C.; Brochon, H.; Kim, P.; Kaur, M.; Kim, Y.-S.; Jang, H.-I.; Kim, Y.-I. Magnesium regulates the circadian oscillator in cyanobacteria. J. Biol. Rhythm. 2019, 34, 380–390. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.I.; Dong, G.; Carruthers, C.W., Jr.; Golden, S.S.; LiWang, A. The day/night switch in KaiC, a central oscillator component of the circadian clock of cyanobacteria. Proc. Natl. Acad. Sci. USA 2008, 105, 12825–12830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vakonakis, I.; LiWang, A.C. Structure of the C-terminal domain of the clock protein KaiA in complex with a KaiC-derived peptide: Implications for KaiC regulation. Proc. Natl. Acad. Sci. USA 2004, 101, 10925–10930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitayama, Y.; Iwasaki, H.; Nishiwaki, T.; Kondo, T. KaiB functions as an attenuator of KaiC phosphorylation in the cyanobacterial circadian clock system. EMBO 2003, 22, 2127–2134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.-G.; Cohen, S.E.; Phong, C.; Myers, W.K.; Kim, Y.-I.; Tseng, R.; Lin, J.; Zhang, L.; Boyd, J.S.; Lee, Y. A protein fold switch joins the circadian oscillator to clock output in cyanobacteria. Science 2015, 349, 324–328. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.G.; Kuo, N.W.; Tseng, R.; LiWang, A. Flexibility of the C-terminal, or CII, ring of KaiC governs the rhythm of the circadian clock of cyanobacteria. Proc. Natl. Acad. Sci. USA 2011, 108, 14431–14436. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.G.; Tseng, R.; Kuo, N.W.; LiWang, A. Rhythmic ring-ring stacking drives the circadian oscillator clockwise. Proc. Natl. Acad. Sci. USA 2012, 109, 16847–16851. [Google Scholar] [CrossRef] [Green Version]
- Rust, M.J.; Markson, J.S.; Lane, W.S.; Fisher, D.S.; O’Shea, E.K. Ordered phosphorylation governs oscillation of a three-protein circadian clock. Science 2007, 318, 809–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishiwaki, T.; Satomi, Y.; Kitayama, Y.; Terauchi, K.; Kiyohara, R.; Takao, T.; Kondo, T. A sequential program of dual phosphorylation of KaiC as a basis for circadian rhythm in cyanobacteria. EMBO. 2007, 26, 4029–4037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijayan, V.; Zuzow, R.; O’Shea, E.K. Oscillations in supercoiling drive circadian gene expression in cyanobacteria. Proc. Natl. Acad. Sci. USA 2009, 106, 22564–22568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihalcescu, I.; Hsing, W.; Leibler, S. Resilient circadian oscillator revealed in individual cyanobacteria. Nature 2004, 430, 81–85. [Google Scholar] [CrossRef]
- Ito, H.; Kageyama, H.; Mutsuda, M.; Nakajima, M.; Oyama, T.; Kondo, T. Autonomous synchronization of the circadian KaiC phosphorylation rhythm. Nat. Struct. Mol. Biol. 2007, 14, 1084–1088. [Google Scholar] [CrossRef]
- Ouyang, Y.; Andersson, C.R.; Kondo, T.; Golden, S.S.; Johnson, C.H. Resonating circadian clocks enhance fitness in cyanobacteria. Proc. Natl. Acad. Sci. USA 1998, 95, 8660–8664. [Google Scholar] [CrossRef] [Green Version]
- Woelfle, M.A.; Ouyang, Y.; Phanvijhitsiri, K.; Johnson, C.H. The adaptive value of circadian clocks: An experimental assessment in cyanobacteria. Curr. Biol. 2004, 14, 1481–1486. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.I.; Boyd, J.S.; Espinosa, J.; Golden, S.S. Detecting KaiC phosphorylation rhythms of the cyanobacterial circadian oscillator in vitro and in vivo. Methods Enzymol. 2015, 551, 153–173. [Google Scholar] [CrossRef] [Green Version]
- Kim, P.; Kaszuba, A.; Jang, H.-I.; Kim, Y.-I. Purification of GST-Fused Cyanobacterial Central Oscillator Protein KaiC. Appl. Biochem. Microbiol. 2020, 56, 395–399. [Google Scholar] [CrossRef]
- Taton, A.; Erikson, C.; Yang, Y.; Rubin, B.E.; Rifkin, S.A.; Golden, J.W.; Golden, S.S. The circadian clock and darkness control natural competence in cyanobacteria. Nat. Commun. 2020, 11, 1688. [Google Scholar] [CrossRef] [Green Version]
- Kiyohara, Y.B.; Katayama, M.; Kondo, T. A novel mutation in kaiC affects resetting of the cyanobacterial circadian clock. J. Bacteriol. 2005, 187, 2559–2564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutsuda, M.; Michel, K.P.; Zhang, X.; Montgomery, B.L.; Golden, S.S. Biochemical properties of CikA, an unusual phytochrome-like histidine protein kinase that resets the circadian clock in Synechococcus elongatus PCC 7942. J. Biol. Chem. 2003, 278, 19102–19110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rust, M.J.; Golden, S.S.; O’Shea, E.K. Light-driven changes in energy metabolism directly entrain the cyanobacterial circadian oscillator. Science 2011, 331, 220–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.I.; Vinyard, D.J.; Ananyev, G.M.; Dismukes, G.C.; Golden, S.S. Oxidized quinones signal onset of darkness directly to the cyanobacterial circadian oscillator. Proc. Natl. Acad. Sci. USA 2012, 109, 17765–17769. [Google Scholar] [CrossRef] [Green Version]
- Ivleva, N.B.; Bramlett, M.R.; Lindahl, P.A.; Golden, S.S. LdpA: A component of the circadian clock senses redox state of the cell. EMBO J. 2005, 24, 1202–1210. [Google Scholar] [CrossRef] [Green Version]
- Katayama, M.; Kondo, T.; Xiong, J.; Golden, S.S. ldpA encodes an iron-sulfur protein involved in light-dependent modulation of the circadian period in the cyanobacterium Synechococcus elongatus PCC 7942. J. Bacteriol. 2003, 185, 1415–1422. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, O.; Katayama, M.; Williams, S.B.; Kondo, T.; Golden, S.S. CikA, a bacteriophytochrome that resets the cyanobacterial circadian clock. Science 2000, 289, 765–768. [Google Scholar] [CrossRef] [Green Version]
- Wood, T.L.; Bridwell-Rabb, J.; Kim, Y.-I.; Gao, T.; Chang, Y.-G.; LiWang, A.; Barondeau, D.P.; Golden, S.S. The KaiA protein of the cyanobacterial circadian oscillator is modulated by a redox-active cofactor. Proc. Natl. Acad. Sci. USA 2010, 107, 5804–5809. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y. Molecular mechanisms of entrainment in the Neurospora circadian clock. J. Biol. Rhythm. 2003, 18, 195–205. [Google Scholar] [CrossRef]
- Millar, A.J. A suite of photoreceptors entrains the plant circadian clock. J. Biol. Rhythm. 2003, 18, 217–226. [Google Scholar] [CrossRef]
- Tseng, R.; Goularte, N.F.; Chavan, A.; Luu, J.; Cohen, S.E.; Chang, Y.G.; Heisler, J.; Li, S.; Michael, A.K.; Tripathi, S.; et al. Structural basis of the day-night transition in a bacterial circadian clock. Science 2017, 355, 1174–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, M.; Ng, A.; Kim, P.; Diekman, C.; Kim, Y.I. CikA Modulates the Effect of KaiA on the Period of the Circadian Oscillation in KaiC Phosphorylation. J. Biol. Rhythm. 2019, 34, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, M.; Ito, H.; Kondo, T. In vitro regulation of circadian phosphorylation rhythm of cyanobacterial clock protein KaiC by KaiA and KaiB. FEBS Lett. 2010, 584, 898–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Dong, G.; Golden, S.S. The pseudo-receiver domain of CikA regulates the cyanobacterial circadian input pathway. Mol. Microbiol. 2006, 60, 658–668. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Zhang, X.; Ivleva, N.B.; Golden, S.S.; LiWang, A. NMR structure of the pseudo-receiver domain of CikA. Protein Sci. 2007, 16, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.; Porr, B.; Mori, T.; Kim, Y.S.; Johnson, C.H.; Diekman, C.O.; Kim, Y.I. CikA, an Input Pathway Component, Senses the Oxidized Quinone Signal to Generate Phase Delays in the Cyanobacterial Circadian Clock. J. Biol. Rhythms 2020, 35, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, G.-K.; Song, Z.-G. Entrainment mechanism of the cyanobacterial circadian clock induced by oxidized quinone. Chin. Phys. B 2020, 29, 098703. [Google Scholar] [CrossRef]
- Diamond, S.; Rubin, B.E.; Shultzaberger, R.K.; Chen, Y.; Barber, C.D.; Golden, S.S. Redox crisis underlies conditional light-dark lethality in cyanobacterial mutants that lack the circadian regulator, RpaA. Proc. Natl. Acad. Sci. USA 2017, 114, E580–E589. [Google Scholar] [CrossRef] [Green Version]
- Ito, H.; Mutsuda, M.; Murayama, Y.; Tomita, J.; Hosokawa, N.; Terauchi, K.; Sugita, C.; Sugita, M.; Kondo, T.; Iwasaki, H. Cyanobacterial daily life with Kai-based circadian and diurnal genome-wide transcriptional control in Synechococcus elongatus. Proc. Natl. Acad. Sci. USA 2009, 106, 14168–14173. [Google Scholar] [CrossRef] [Green Version]
- Takano, S.; Tomita, J.; Sonoike, K.; Iwasaki, H. The initiation of nocturnal dormancy in Synechococcus as an active process. BMC Biol. 2015, 13, 36. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, H.; Williams, S.B.; Kitayama, Y.; Ishiura, M.; Golden, S.S.; Kondo, T. A kaiC-interacting sensory histidine kinase, SasA, necessary to sustain robust circadian oscillation in cyanobacteria. Cell 2000, 101, 223–233. [Google Scholar] [CrossRef] [Green Version]
- Vakonakis, I.; Klewer, D.A.; Williams, S.B.; Golden, S.S.; LiWang, A.C. Structure of the N-terminal domain of the circadian clock-associated histidine kinase SasA. J. Mol. Biol. 2004, 342, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Pattanayek, R.; Williams, D.R.; Rossi, G.; Weigand, S.; Mori, T.; Johnson, C.H.; Stewart, P.L.; Egli, M. Combined SAXS/EM based models of the S. elongatus post-translational circadian oscillator and its interactions with the output His-kinase SasA. PLoS ONE 2011, 6, e23697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heisler, J.; Swan, J.A.; Palacios, J.G.; Sancar, C.; Ernst, D.C.; Spangler, R.K.; Bagshaw, C.R.; Tripathi, S.; Crosby, P.; Golden, S.S.; et al. Structural mimicry confers robustness in the cyanobacterial circadian clock. bioRxiv 2020. [Google Scholar] [CrossRef]
- Gutu, A.; O’Shea, E.K. Two antagonistic clock-regulated histidine kinases time the activation of circadian gene expression. Mol. Cell 2013, 50, 288–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takai, N.; Nakajima, M.; Oyama, T.; Kito, R.; Sugita, C.; Sugita, M.; Kondo, T.; Iwasaki, H. A KaiC-associating SasA-RpaA two-component regulatory system as a major circadian timing mediator in cyanobacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 12109–12114. [Google Scholar] [CrossRef] [Green Version]
- Fleming, K.E.; O’Shea, E.K. An RpaA-Dependent Sigma Factor Cascade Sets the Timing of Circadian Transcriptional Rhythms in Synechococcus elongatus. Cell Rep. 2018, 25, 2937–2945.e2933. [Google Scholar] [CrossRef] [Green Version]
- Teng, S.W.; Mukherji, S.; Moffitt, J.R.; de Buyl, S.; O’Shea, E.K. Robust circadian oscillations in growing cyanobacteria require transcriptional feedback. Science 2013, 340, 737–740. [Google Scholar] [CrossRef] [Green Version]
- Dong, G.; Yang, Q.; Wang, Q.; Kim, Y.I.; Wood, T.L.; Osteryoung, K.W.; van Oudenaarden, A.; Golden, S.S. Elevated ATPase activity of KaiC applies a circadian checkpoint on cell division in Synechococcus elongatus. Cell 2010, 140, 529–539. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.M.; Williams, S.B. Circadian rhythms in gene transcription imparted by chromosome compaction in the cyanobacterium Synechococcus elongatus. Proc. Natl. Acad. Sci. USA 2006, 103, 8564–8569. [Google Scholar] [CrossRef] [Green Version]
- Woelfle, M.A.; Xu, Y.; Qin, X.; Johnson, C.H. Circadian rhythms of superhelical status of DNA in cyanobacteria. Proc. Natl. Acad. Sci. USA 2007, 104, 18819–18824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavan, A.G.; Ernst, D.C.; Fang, M.; Sancar, C.; Partch, C.L.; Golden, S.S.; LiWang, A. Reconstitution of an intact clock that generates circadian DNA binding in vitro. bioRxiv 2020. [Google Scholar] [CrossRef]
- Holtzendorff, J.; Partensky, F.; Mella, D.; Lennon, J.F.; Hess, W.R.; Garczarek, L. Genome streamlining results in loss of robustness of the circadian clock in the marine cyanobacterium Prochlorococcus marinus PCC 9511. J. Biol. Rhythm. 2008, 23, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Axmann, I.M.; Duhring, U.; Seeliger, L.; Arnold, A.; Vanselow, J.T.; Kramer, A.; Wilde, A. Biochemical evidence for a timing mechanism in prochlorococcus. J. Bacteriol. 2009, 191, 5342–5347. [Google Scholar] [CrossRef] [Green Version]
- Baca, I.; Sprockett, D.; Dvornyk, V. Circadian Input Kinases and Their Homologs in Cyanobacteria: Evolutionary Constraints Versus Architectural Diversification. J. Mol. Evol. 2010, 70, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Wiegard, A.; Dorrich, A.K.; Deinzer, H.T.; Beck, C.; Wilde, A.; Holtzendorff, J.; Axmann, I.M. Biochemical analysis of three putative KaiC clock proteins from Synechocystis sp. PCC 6803 suggests their functional divergence. Microbiol. Read. 2013, 159, 948–958. [Google Scholar] [CrossRef] [Green Version]
- Anderson, S.L.; McIntosh, L. Light-activated heterotrophic growth of the cyanobacterium Synechocystis sp. strain PCC 6803: A blue-light-requiring process. J. Bacteriol. 1991, 173, 2761–2767. [Google Scholar] [CrossRef] [Green Version]
- Aoki, S.; Kondo, T.; Wada, H.; Ishiura, M. Circadian rhythm of the cyanobacterium Synechocystis sp. strain PCC 6803 in the dark. J. Bacteriol. 1997, 179, 5751–5755. [Google Scholar] [CrossRef] [Green Version]
- Yamaoka, T.; Satoh, K.; Katoh, S. Photosynthetic activities of a thermophilic blue-green alga. Plant Cell Physiol. 1978, 19, 943–954. [Google Scholar] [CrossRef]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic Assignments, Strain Histories and Properties of Pure Cultures of Cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Vakonakis, I.; Sun, J.; Wu, T.; Holzenburg, A.; Golden, S.S.; LiWang, A.C. NMR structure of the KaiC-interacting C-terminal domain of KaiA, a circadian clock protein: Implications for KaiA-KaiC interaction. Proc. Natl. Acad. Sci. USA 2004, 101, 1479–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pattanayek, R.; Williams, D.R.; Pattanayek, S.; Mori, T.; Johnson, C.H.; Stewart, P.L.; Egli, M. Structural model of the circadian clock KaiB-KaiC complex and mechanism for modulation of KaiC phosphorylation. EMBO J. 2008, 27, 1767–1778. [Google Scholar] [CrossRef] [PubMed]
- Pattanayek, R.; Williams, D.R.; Pattanayek, S.; Xu, Y.; Mori, T.; Johnson, C.H.; Stewart, P.L.; Egli, M. Analysis of KaiA-KaiC protein interactions in the cyano-bacterial circadian clock using hybrid structural methods. EMBO J. 2006, 25, 2017–2028. [Google Scholar] [CrossRef] [PubMed]
- Onai, K.; Morishita, M.; Itoh, S.; Okamoto, K.; Ishiura, M. Circadian rhythms in the thermophilic cyanobacterium Thermosynechococcus elongatus: Compensation of period length over a wide temperature range. J. Bacteriol. 2004, 186, 4972–4977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hood, R.D.; Higgins, S.A.; Flamholz, A.; Nichols, R.J.; Savage, D.F. The stringent response regulates adaptation to darkness in the cyanobacterium Synechococcus elongatus. Proc. Natl. Acad. Sci. USA 2016, 113, E4867–E4876. [Google Scholar] [CrossRef] [Green Version]
- Puszynska, A.M.; O’Shea, E.K. ppGpp Controls Global Gene Expression in Light and in Darkness in S. elongatus. Cell Rep. 2017, 21, 3155–3165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinosa, J.; Boyd, J.S.; Cantos, R.; Salinas, P.; Golden, S.S.; Contreras, A. Cross-talk and regulatory interactions between the essential response regulator RpaB and cyanobacterial circadian clock output. Proc. Natl. Acad. Sci. USA 2015, 112, 2198–2203. [Google Scholar] [CrossRef] [Green Version]
- Kasting, J.F.; Siefert, J.L. Life and the evolution of Earth’s atmosphere. Science 2002, 296, 1066–1068. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, P.; Kaur, M.; Jang, H.-I.; Kim, Y.-I. The Circadian Clock—A Molecular Tool for Survival in Cyanobacteria. Life 2020, 10, 365. https://doi.org/10.3390/life10120365
Kim P, Kaur M, Jang H-I, Kim Y-I. The Circadian Clock—A Molecular Tool for Survival in Cyanobacteria. Life. 2020; 10(12):365. https://doi.org/10.3390/life10120365
Chicago/Turabian StyleKim, Pyonghwa, Manpreet Kaur, Hye-In Jang, and Yong-Ick Kim. 2020. "The Circadian Clock—A Molecular Tool for Survival in Cyanobacteria" Life 10, no. 12: 365. https://doi.org/10.3390/life10120365