VSIG4 Induces Epithelial-Mesenchymal Transition of Renal Tubular Cells under High-Glucose Conditions

 , and
, and 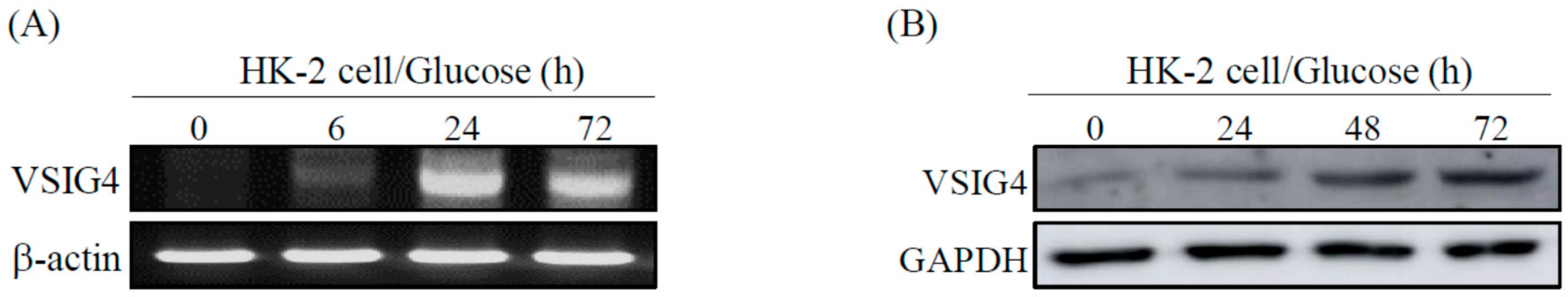{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. VSIG4 Upregulation by HG Stimulation in Renal Tubular Epithelial HK-2 Cells
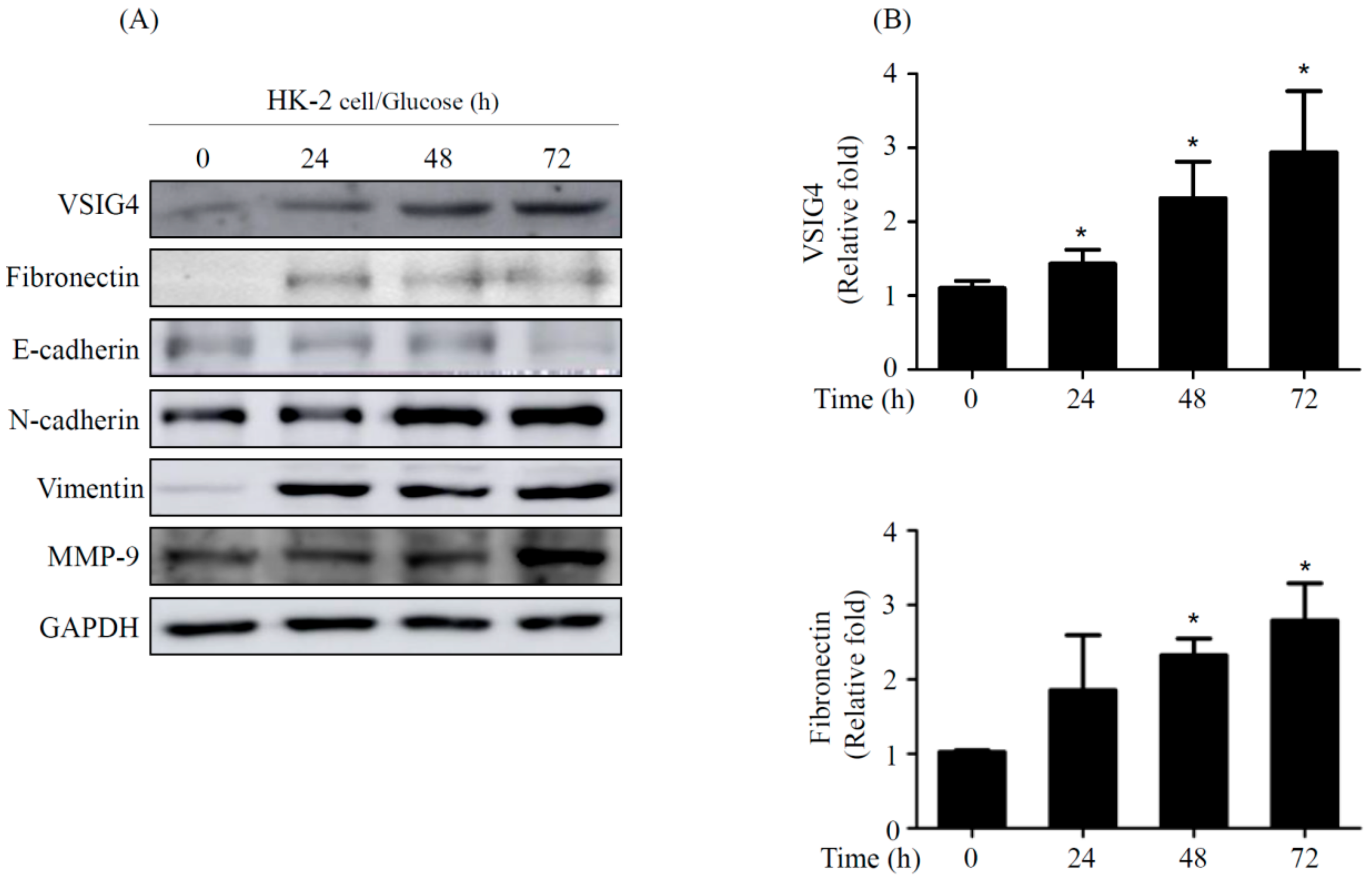
2.2. EMT by VSIG4 Upregulation in Renal Tubular Cells Stimulated by HG
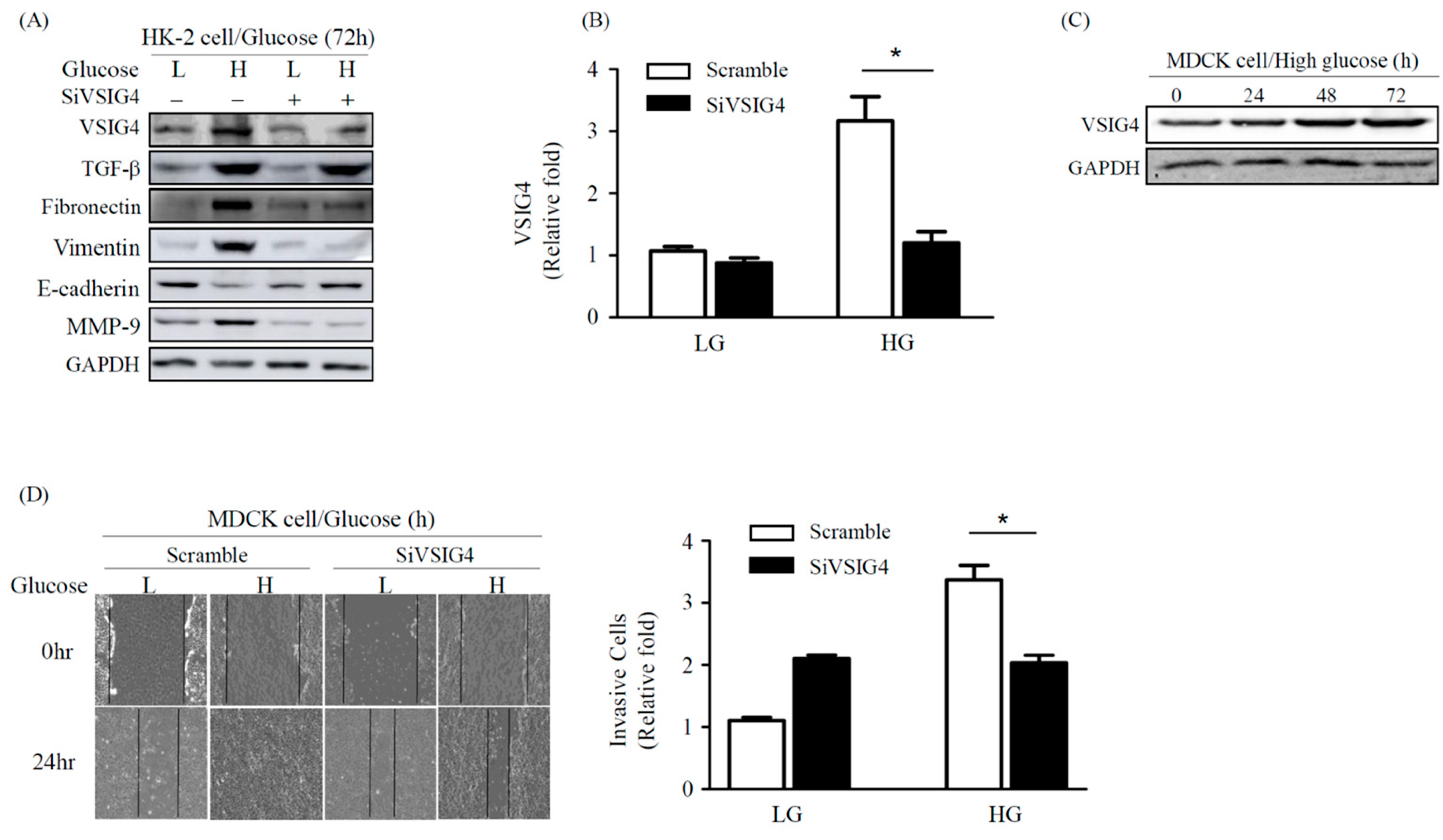
2.3. Effect of VSIG4 on EMT and Proliferation of Renal Epithelial MDCK Cells
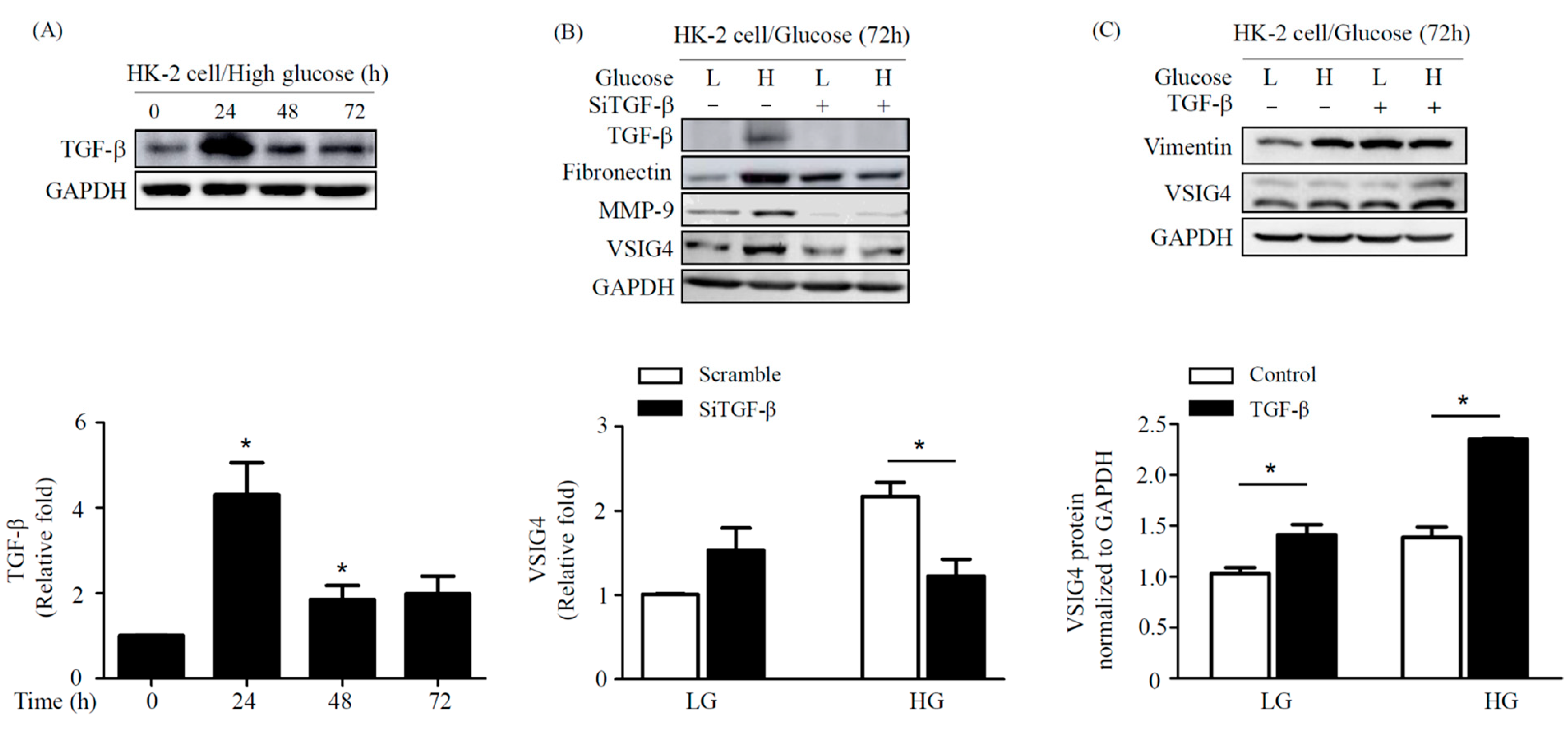
2.4. Effect of TGF-β on Fibrosis Marker in HG Conditions in HK-2 Cells
3. Discussion
4. Materials and Methods
4.1. Cells Culture and Reagents
4.2. RNA Interference
4.3. Western Blot Analysis
4.4. Reverse Transcription Polymerase Chain Reaction (RT-PCR) Analysis
4.5. Cell Motility and Invasion Assay
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| HG | high glucose |
| EMT | epithelial-mesenchymal transition |
| VSIG4 | V-set immunoglobulin-domain-containing 4 |
| TGF | transforming growth factor |
| MDCK | Madin Darby Canine Kidney |
References
- Burns, W.C.; Twigg, S.M.; Forbes, J.M.; Pete, J.; Tikellis, C.; Thallas-Bonke, V.; Thomas, M.C.; Cooper, M.E.; Kantharidis, P. Connective tissue growth factor plays an important role in advanced glycation end product–induced tubular epithelial-to-mesenchymal transition: Implications for diabetic renal disease. J. Am. Soc. Nephrol. 2006, 17, 2484–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonson, M. Phenotypic transitions and fibrosis in diabetic nephropathy. Kidney Int. 2007, 71, 846–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hills, C.E.; Squires, P.E. TGF-β1-induced epithelial-to-mesenchymal transition and therapeutic intervention in diabetic nephropathy. Am. J. Nephrol. 2010, 31, 68–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutariya, B.; Jhonsa, D.; Saraf, M.N. TGF-β: The connecting link between nephropathy and fibrosis. Immunopharmacol. Immunotoxicol. 2016, 38, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Vogt, L.; Schmitz, N.; Kurrer, M.O.; Bauer, M.; Hinton, H.I.; Behnke, S.; Gatto, D.; Sebbel, P.; Beerli, R.R.; Sonderegger, I.; et al. VSIG4, a B7 family–related protein, is a negative regulator of T cell activation. J. Clin. Invest. 2006, 116, 2817–2826. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Q.; Pang, X.; Zhang, Z.; Wei, Y.; Hong, J.; Chen, H. Microglial V-set and immunoglobulin domain-containing 4 protects against ischemic stroke in mice by suppressing TLR4-regulated inflammatory response. Biochem. Biophys. Res. Commun. 2020, 522, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Guo, S.; Chen, Y.; Ca, D.; Xu, H.; Yang, C.; Fei, L.; Ni, B.; Ruan, Z. VSIG4 expression on macrophages facilitates lung cancer development. Lab. Investig. 2014, 94, 706–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.H.; Qian, Y.; Li, Z.; Zhang, N.N.; Xie, Y.J. Let-7g-5p inhibits epithelial-mesenchymal transition consistent with reduction of glioma stem cell phenotypes by targeting VSIG4 in glioblastoma. Oncol. Rep. 2016, 36, 2967–2975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.M.; Oh, S.W.; Park, S.H.; Hur, D.Y.; Hong, S.W.; Han, S.Y. Epstein-Barr virus-encoded latent membrane protein 1 induces epithelial to mesenchymal transition by inducing V-set Ig domain containing 4 (VSIG4) expression via NF-kB in renal tubular epithelial HK-2 cells. Biochem. Biophys. Res. Commun. 2017, 492, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Geng, X.D.; Wang, W.W.; Feng, Z.; Cheng, X.L.; Shen, W.J.; Dng, Z.Y.; Cai, G.Y.; Chen, X.N.; Hong, Q. Identification of key genes and pathways in diabetic nephropathy by bioinformatics analysis. J. Diab. Investig. 2019, 10, 972–984. [Google Scholar] [CrossRef] [PubMed]
- Rao, D.A.; Arazi, A.; Wofsy, D.; Diamond, B. Design and application of single-cell RNA sequencing to study kidney immune cells in lupus nephritis. Nat. Rev. Nephrol. 2019, 16, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Hodgin, J.B.; Berthier, C.C.; John, R.; Grone, E.; Porubsky, S.; Grone, H.J.; Herzenberg, A.M.; Scholey, J.W.; Haldunewich, M.; Cattran, D.C.; et al. The molecular phenotype of endocapillary proliferation: Novel therapeutic targets for IgA nephropathy. PLoS ONE 2014, 9, e103413. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.L.; Wang, F.S.; Kuo, Y.R.; Huang, Y.T.; Huang, H.C.; Sun, Y.C.; Kuo, Y.H. Ras modulation of superoxide activates ERK-dependent fibronectin expression in diabetes-induced renal injuries. Kidney Int. 2006, 69, 1593–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, T.K.; Zheng, G.; Hsu, T.T.; Wng, Y.; Lee, V.W.S.; Tian, Z.; Wang, Y.; Cao, Q.; Wang, Y.; Harris, D.C. Macrophage matrix metalloproteinase-9 mediates epithelial-mesenchymal transition in vitro in murine renal tubular cells. Am. J. Pathol. 2010, 176, 1256–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, E.-Y.; Jo, H.A.; Park, S.H.; Cha, D.R.; Hur, D.Y.; Han, S.Y. VSIG4 Induces Epithelial-Mesenchymal Transition of Renal Tubular Cells under High-Glucose Conditions. Life 2020, 10, 354. https://doi.org/10.3390/life10120354
Gong E-Y, Jo HA, Park SH, Cha DR, Hur DY, Han SY. VSIG4 Induces Epithelial-Mesenchymal Transition of Renal Tubular Cells under High-Glucose Conditions. Life. 2020; 10(12):354. https://doi.org/10.3390/life10120354
Chicago/Turabian StyleGong, Eun-Yeung, Hyung Ah Jo, Sang Hyun Park, Dae Ryong Cha, Dae Young Hur, and Sang Youb Han. 2020. "VSIG4 Induces Epithelial-Mesenchymal Transition of Renal Tubular Cells under High-Glucose Conditions" Life 10, no. 12: 354. https://doi.org/10.3390/life10120354