The Model System Saccharomyces cerevisiae Versus Emerging Non-Model Yeasts for the Production of Biofuels


Abstract
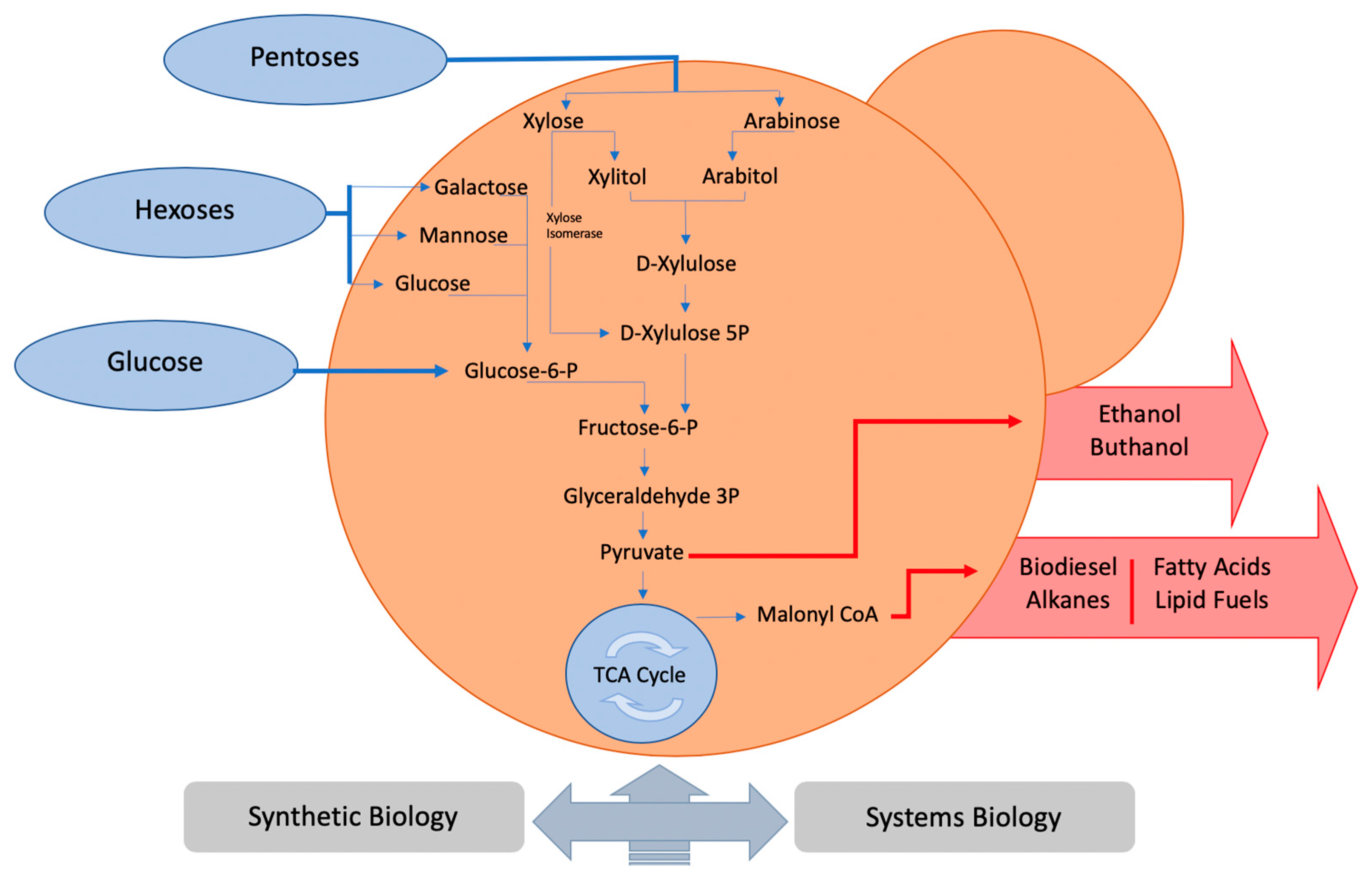
:1. Introduction
2. Conventional and Non-Conventional Yeast Bioproduction Systems
2.1. S. Cerevisiae for the Production of Chemicals Beyond Ethanol
2.2. Y. lipolytica, K. marxianus, I. orientalis and P. pastoris Cell Factories for the Production of Chemicals
2.2.1. Yarrowia Lipolytica
2.2.2. Kluyveromyces Marxianus
2.2.3. Issatchenkia Orientalis
2.2.4. Pichia Pastoris
3. Advancing Technologies for Optimization of Production Pathways and Host
3.1. Classical Genetics Tools and Emerging Technologies
3.2. Systems Biology in Engineering Yeasts for Bioproduction
4. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dellomonaco, C.; Fava, F.; Gonzalez, R. The path to next generation biofuels: Successes and challenges in the era of synthetic biology. Microb. Cell Factories 2010, 9, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, J. Yeast cell factories on the horizon. Science 2015, 349, 1050–1051. [Google Scholar] [CrossRef] [PubMed]
- Madhavan, A.; Jose, A.A.; Binod, P.; Sindhu, R.; Sukumaran, R.K.; Pandey, A.; Castro, G.E. Synthetic Biology and Metabolic Engineering Approaches and Its Impact on Non-Conventional Yeast and Biofuel Production. Front. Energy Res. 2017, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Hansen, A.S.L.; Lennen, R.M.; Sonnenschein, N.; Herrgård, M.J. Systems biology solutions for biochemical production challenges. Curr. Opin. Biotechnol. 2017, 45, 85–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, J. Yeast Systems Biology: Model Organism and Cell Factory. Biotechnol. J. 2019, 14, e1800421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.-S.; Kwak, S.; Turner, T.L.; Jin, Y.-S. Yeast synthetic biology toolbox and applications for biofuel production. FEMS Yeast Res. 2014, 15, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radecka, D.; Mukherjee, V.; Mateo, R.Q.; Stojiljkovic, M.; Foulquié-Moreno, M.R.; Thevelein, J.M. Looking beyond Saccharomyces: The potential of non-conventional yeast species for desirable traits in bioethanol fermentation. FEMS Yeast Res. 2015, 15. [Google Scholar] [CrossRef] [Green Version]
- Deparis, Q.; Claes, A.; Foulquié-Moreno, M.R.; Thevelein, J.M. Engineering tolerance to industrially relevant stress factors in yeast cell factories. FEMS Yeast Res. 2017, 17. [Google Scholar] [CrossRef]
- Thomas, K.C.; Hynes, S.H.; Ingledew, W.M. Influence of Medium Buffering Capacity on Inhibition of Saccharomyces cerevisiae Growth by Acetic and Lactic Acids. Appl. Environ. Microbiol. 2002, 68, 1616–1623. [Google Scholar] [CrossRef] [Green Version]
- Martien, J.; Amador-Noguez, D. Recent applications of metabolomics to advance microbial biofuel production. Curr. Opin. Biotechnol. 2017, 43, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Cai, P.; Gao, J.; Zhou, Y. CRISPR-mediated genome editing in non-conventional yeasts for biotechnological applications. Microb. Cell Factories 2019, 18, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, S.; Ngo, H.H.; Wu, Y.-R. Advanced CRISPR/Cas-based genome editing tools for microbial biofuels production: A review. Renew. Energy 2020, 149, 1107–1119. [Google Scholar] [CrossRef]
- Yang, Z.; Blenner, M. Genome editing systems across yeast species. Curr. Opin. Biotechnol. 2020, 66, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Lian, J.; Mishra, S.; Zhao, H. Recent advances in metabolic engineering of Saccharomyces cerevisiae: New tools and their applications. Metab. Eng. 2018, 50, 85–108. [Google Scholar] [CrossRef] [PubMed]
- Besada-Lombana, P.B.; McTaggart, T.L.; Da Silva, N.A. Molecular tools for pathway engineering in Saccharomyces cerevisiae. Curr. Opin. Biotechnol. 2018, 53, 39–49. [Google Scholar] [CrossRef]
- Deaner, M.; Alper, H.S. Enhanced scale and scope of genome engineering and regulation using CRISPR/Cas in Saccharomyces cerevisiae. FEMS Yeast Res. 2019, 19, 76. [Google Scholar] [CrossRef]
- Wagner, J.M.; Alper, H.S. Synthetic biology and molecular genetics in non-conventional yeasts: Current tools and future advances. Fungal Genet. Biol. 2016, 89, 126–136. [Google Scholar] [CrossRef] [Green Version]
- Löbs, A.-K.; Schwartz, C.; Wheeldon, I. Genome and metabolic engineering in non-conventional yeasts: Current advances and applications. Synth. Syst. Biotechnol. 2017, 2, 198–207. [Google Scholar] [CrossRef]
- Passoth, V. Conventional and Non-Conventional Yeasts for the Production of Biofuels; Springer Science and Business Media LLC: Berlin, Germany, 2017; pp. 385–416. [Google Scholar]
- Goffeau, A.; Phillippsen, P.; Botstein, D. Yeast ferments genome-wide functional analyses. Nat. Genet. 1996, 14, 1–2. [Google Scholar]
- Dikicioglu, D.; Pir, P.; Oliver, S.G. Predicting complex phenotype–genotype interactions to enable yeast engineering: Saccharomyces cerevisiae as a model organism and a cell factory. Biotechnol. J. 2013, 8, 1017–1034. [Google Scholar] [CrossRef] [Green Version]
- Kavšček, M.; Stražar, M.; Curk, T.; Natter, K.; Petrovič, U. Yeast as a cell factory: Current state and perspectives. Microb. Cell Factories 2015, 14, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, K.-K.; Nielsen, J. Metabolic engineering of Saccharomyces cerevisiae: A key cell factory platform for future biorefineries. Cell. Mol. Life Sci. 2012, 69, 2671–2690. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.-K.; Roldão, A.; Siewers, V.; Nielsen, J. A systems-level approach for metabolic engineering of yeast cell factories. FEMS Yeast Res. 2012, 12, 228–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christie, K.R.; Weng, S.; Balakrishnan, R.; Costanzo, M.C.; Dolinski, K.; Dwight, S.S.; Engel, S.R.; Feierbach, B.; Fisk, D.G.; Hirschman, J.E.; et al. Saccharomyces Genome Database (SGD) provides tools to identify and analyze sequences from Saccharomyces cerevisiae and related sequences from other organisms. Nucleic Acids Res. 2004, 32, D311–D314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krink-Koutsoubelis, N.; Loechner, A.C.; Lechner, A.; Link, H.; Denby, C.M.; Voegeli, B.; Erb, T.J.; Yuzawa, S.; Jakociunas, T.; Katz, L.; et al. Engineered Production of Short-Chain Acyl-Coenzyme A Esters inSaccharomyces cerevisiae. ACS Synth. Biol. 2018, 7, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Banerjee, D.; Mukhopadhyay, A.; Petzold, C.J. Systems and synthetic biology tools for advanced bioproduction hosts. Curr. Opin. Biotechnol. 2020, 64, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Srirangan, K.; Bruder, M.; Akawi, L.; Miscevic, D.; Kilpatrick, S.; Moo-Young, M.; Chou, C.P. Recent advances in engineering propionyl-CoA metabolism for microbial production of value-added chemicals and biofuels. Crit. Rev. Biotechnol. 2016, 37, 701–722. [Google Scholar] [CrossRef]
- Valle-Rodríguez, J.O.; Shi, S.; Siewers, V.; Nielsen, J. Metabolic engineering of Saccharomyces cerevisiae for production of fatty acid ethyl esters, an advanced biofuel, by eliminating non-essential fatty acid utilization pathways. Appl. Energy 2014, 115, 226–232. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.J.; Buijs, N.A.; Zhu, Z.; Qin, J.; Siewers, V.; Nielsen, J. Production of fatty acid-derived oleochemicals and biofuels by synthetic yeast cell factories. Nat. Commun. 2016, 7, 11709. [Google Scholar] [CrossRef] [Green Version]
- Foo, J.L.; Susanto, A.V.; Keasling, J.D.; Leong, S.S.J.; Chang, M.W. Whole-cell biocatalytic and de novo production of alkanes from free fatty acids in Saccharomyces cerevisiae. Biotechnol. Bioeng. 2017, 114, 232–237. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.-W.; Theisen, M.K.; Liao, J.C. Metabolic systems modeling for cell factories improvement. Curr. Opin. Biotechnol. 2017, 46, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Crespo, J.L.; Daicho, K.; Ushimaru, T.; Hall, M.N. The GATA Transcription Factors GLN3 and GAT1 Link TOR to Salt Stress in Saccharomyces cerevisiae. J. Biol. Chem. 2001, 276, 34441–34444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahsavarani, H.; Hasegawa, D.; Yokota, D.; Sugiyama, M.; Kaneko, Y.; Boonchird, C.; Harashima, S. Enhanced bio-ethanol production from cellulosic materials by semi-simultaneous saccharification and fermentation using high temperature resistant Saccharomyces cerevisiae TJ14. J. Biosci. Bioeng. 2013, 115, 20–23. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Jiang, R. Improving Saccharomyces cerevisiae ethanol production and tolerance via RNA polymerase II subunit Rpb7. Biotechnol. Biofuels 2017, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.K.; Kumar, M.; Kumari, S.; Kumari, P.; Yusuf, F.; Jakeer, S.; Naz, S.; Chandna, P.; Bhatnagar, I.; Gaur, N.A. Evaluation of divergent yeast genera for fermentation-associated stresses and identification of a robust sugarcane distillery waste isolate Saccharomyces cerevisiae NGY10 for lignocellulosic ethanol production in SHF and SSF. Biotechnol. Biofuels 2019, 12, 40. [Google Scholar] [CrossRef] [Green Version]
- Matsushika, A.; Negi, K.; Suzuki, T.; Goshima, T.; Hoshino, T. Identification and Characterization of a Novel Issatchenkia orientalis GPI-Anchored Protein, IoGas1, Required for Resistance to Low pH and Salt Stress. PLoS ONE 2016, 11, e0161888. [Google Scholar] [CrossRef]
- Qiao, K.; Abidi, S.H.I.; Liu, H.; Zhang, H.; Chakraborty, S.; Watson, N.; Ajikumar, P.K.; Stephanopoulos, G. Engineering lipid overproduction in the oleaginous yeast Yarrowia lipolytica. Metab. Eng. 2015, 29, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Papouskova, K.; Sychrova, H. Yarrowia lipolytica possesses two plasma membrane alkali metal cation/H+ antiporters with different functions in cell physiology. FEBS Lett. 2006, 580, 1971–1976. [Google Scholar] [CrossRef] [Green Version]
- Hackenschmidt, S.; Bracharz, F.; Daniel, R.; Thürmer, A.; Bruder, S.; Kabisch, J. Effects of a high-cultivation temperature on the physiology of three different Yarrowia lipolytica strains. FEMS Yeast Res. 2019, 19, 068. [Google Scholar] [CrossRef]
- Biryukova, E.N.; Arinbasarova, A.Y.; Suzina, N.E.; Sorokin, V.V.; Medentsev, A.G. Ultrastructural changes in Yarrowia lipolytica cells under stress conditions. Microbiol. 2011, 80, 350–354. [Google Scholar] [CrossRef]
- Cavka, A.; Jönsson, L.J. Comparison of the growth of filamentous fungi and yeasts in lignocellulose-derived media. Biocatal. Agric. Biotechnol. 2014, 3, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Cui, Z.; Gao, C.; Li, J.; Hou, J.; Lin, C.S.K.; Qi, Q. Engineering of unconventional yeast Yarrowia lipolytica for efficient succinic acid production from glycerol at low pH. Metab. Eng. 2017, 42, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Wang, J.; Zhu, Y.; Zhang, B.; Kong, X.; Li, W.; Wang, D.; Hong, J. Release of glucose repression on xylose utilization in Kluyveromyces marxianus to enhance glucose-xylose co-utilization and xylitol production from corncob hydrolysate. Microb. Cell Factories 2019, 18, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Erdei, É.; Molnár, M.; Gyémánt, G.; Antal, K.; Emri, T.; Pócsi, I.; Nagy, J. Trehalose overproduction affects the stress tolerance of Kluyveromyces marxianus ambiguously. Bioresour. Technol. 2011, 102, 7232–7235. [Google Scholar] [CrossRef] [PubMed]
- Diniz, R.H.S.; Villada, J.C.; Alvim, M.C.T.; Vidigal, P.M.P.; Vieira, N.M.; Lamas-Maceiras, M.; Cerdán, M.-E.; González-Siso, M.-I.; Lahtvee, P.-J.; Da Silveira, W.B. Transcriptome analysis of the thermotolerant yeast Kluyveromyces marxianus CCT 7735 under ethanol stress. Appl. Microbiol. Biotechnol. 2017, 101, 6969–6980. [Google Scholar] [CrossRef]
- Rugthaworn, P.; Murata, Y.; Machida, M.; Apiwatanapiwat, W.; Hirooka, A.; Thanapase, W.; Dangjarean, H.; Ushiwaka, S.; Morimitsu, K.; Kosugi, A.; et al. Growth Inhibition of Thermotolerant Yeast, Kluyveromyces marxianus, in Hydrolysates from Cassava Pulp. Appl. Biochem. Biotechnol. 2014, 173, 1197–1208. [Google Scholar] [CrossRef]
- Yadav, J.; Bezawada, J.; Ajila, C.; Yan, S.; Tyagi, R.; Surampalli, R. Mixed culture of Kluyveromyces marxianus and Candida krusei for single-cell protein production and organic load removal from whey. Bioresour. Technol. 2014, 164, 119–127. [Google Scholar] [CrossRef]
- Cheng, H.; Lv, J.; Wang, H.; Wang, B.; Li, Z.; Deng, Z. Genetically engineered Pichia pastoris yeast for conversion of glucose to xylitol by a single-fermentation process. Appl. Microbiol. Biotechnol. 2014, 98, 3539–3552. [Google Scholar] [CrossRef]
- Dragosits, M.; Mattanovich, D.; Gasser, B. Induction and Measurement of UPR and Osmotic Stress in the Yeast Pichia Pastoris; Elsevier: Amsterdam, The Netherlands, 2011; Volume 489, pp. 165–188. [Google Scholar]
- Sakaki, T.; Zähringer, U.; Warnecke, D.C.; Fahl, A.; Knogge, W.; Heinz, E. Sterol glycosides and cerebrosides accumulate inPichia pastoris, Rhynchosporium secalisand other fungi under normal conditions or under heat shock and ethanol stress. Yeast 2001, 18, 679–695. [Google Scholar] [CrossRef]
- Seman, W.W.; Bakar, S.; Bukhari, N.; Gaspar, S.; Othman, R.; Nathan, S.; Mahadi, N.; Jahim, J.; Murad, A.; Abu Bakar, F.D. High level expression of Glomerella cingulata cutinase in dense cultures of Pichia pastoris grown under fed-batch conditions. J. Biotechnol. 2014, 184, 219–228. [Google Scholar] [CrossRef]
- Miao, Y.; Xiong, G.; Li, R.; Wu, Z.; Zhang, X.; Weng, P. Transcriptome profiling of Issatchenkia orientalis under ethanol stress. AMB Express 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kampranis, S.C.; Makris, A.M. Developing a yeast cell factory for the production of terpenoids. Comput. Struct. Biotechnol. J. 2012, 3, e201210006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghildiyal, R.; Bisht, G.S.; Shrivastava, R. Engineering Yeast as Cellular Factory. Metab. Eng. Bioact. Compounds 2017, 173–208. [Google Scholar] [CrossRef]
- Lane, S.; Zhang, Y.; Yun, E.J.; Ziolkowski, L.; Zhang, G.; Jin, Y.-S.; Avalos, J.L. Xylose assimilation enhances the production of isobutanol in engineered Saccharomyces cerevisiae. Biotechnol. Bioeng. 2019, 117, 372–381. [Google Scholar] [CrossRef]
- Wess, J.; Brinek, M.; Boles, E. Improving isobutanol production with the yeast Saccharomyces cerevisiae by successively blocking competing metabolic pathways as well as ethanol and glycerol formation. Biotechnol. Biofuels 2019, 12, 1–15. [Google Scholar] [CrossRef]
- Pronk, J.T.; Yde Steensma, H.; Van Dijken, J.P. Pyruvate metabolism in Saccharomyces cerevisiae. Yeast 1996, 12, 1607–1633. [Google Scholar] [CrossRef]
- Sakihama, Y.; Hidese, R.; Hasunuma, T.; Kondo, A. Increased flux in acetyl-CoA synthetic pathway and TCA cycle of Kluyveromyces marxianus under respiratory conditions. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Quintelas, C.; Braga, A.; Mesquita, D.P.; Amaral, A.L.; Ferreira, E.C.; Belo, I. NIR spectroscopy applied to the determination of 2-phenylethanol and l -phenylalanine concentrations in culture medium of Yarrowia lipolytica. J. Chem. Technol. Biotechnol. 2018, 94, 812–818. [Google Scholar] [CrossRef]
- Ata, Ö.; Rebnegger, C.; Tatto, N.E.; Valli, M.; Mairinger, T.; Hann, S.; Steiger, M.G.; Çalık, P.; Mattanovich, D. A single Gal4-like transcription factor activates the Crabtree effect in Komagataella phaffii. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Qiao, K.; Wasylenko, T.M.; Zhou, K.; Xu, P.; Stephanopoulos, G. Lipid production in Yarrowia lipolytica is maximized by engineering cytosolic redox metabolism. Nat. Biotechnol. 2017, 35, 173–177. [Google Scholar] [CrossRef]
- Hanko, E.K.; Denby, C.M.; I Nogué, V.S.; Lin, W.; Ramirez, K.J.; Singer, C.A.; Beckham, G.T.; Keasling, J.D. Engineering β-oxidation in Yarrowia lipolytica for methyl ketone production. Metab. Eng. 2018, 48, 52–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darvishi, F.; Fathi, Z.; Ariana, M.; Moradi, H. Yarrowia lipolytica as a workhorse for biofuel production. Biochem. Eng. J. 2017, 127, 87–96. [Google Scholar] [CrossRef]
- Blazeck, J.; Reed, B.; Garg, R.; Gerstner, R.; Pan, A.; Agarwala, V.; Alper, H.S. Generalizing a hybrid synthetic promoter approach in Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2012, 97, 3037–3052. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Nambou, K.; Wei, L.-J.; Hua, Q. Heterologous production of α-farnesene in metabolically engineered strains of Yarrowia lipolytica. Bioresour. Technol. 2016, 216, 1040–1048. [Google Scholar] [CrossRef] [PubMed]
- Bellou, S.; Makri, A.; Triantaphyllidou, I.-E.; Papanikolaou, S.; Aggelis, G. Morphological and metabolic shifts of Yarrowia lipolytica induced by alteration of the dissolved oxygen concentration in the growth environment. Microbiology 2014, 160, 807–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beopoulos, A.; Nicaud, J.-M.; Gaillardin, C. An overview of lipid metabolism in yeasts and its impact on biotechnological processes. Appl. Microbiol. Biotechnol. 2011, 90, 1193–1206. [Google Scholar] [CrossRef]
- Niehus, X.; Coq, A.-M.C.-L.; Sandoval, G.; Nicaud, J.-M.; Ledesma-Amaro, R. Engineering Yarrowia lipolytica to enhance lipid production from lignocellulosic materials. Biotechnol. Biofuels 2018, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lazar, Z.; Liu, N.; Stephanopoulos, G. Holistic Approaches in Lipid Production by Yarrowia lipolytica. Trends Biotechnol. 2018, 36, 1157–1170. [Google Scholar] [CrossRef]
- Madeira, J.V., Jr.; Gombert, A.K. Towards high-temperature fuel ethanol production using Kluyveromyces marxianus: On the search for plug-in strains for the Brazilian sugarcane-based biorefinery. Biomass Bioenergy 2018, 119, 217–228. [Google Scholar] [CrossRef]
- Dasgupta, D.; Ghosh, D.; Bandhu, S.; Adhikari, D.K. Lignocellulosic sugar management for xylitol and ethanol fermentation with multiple cell recycling by Kluyveromyces marxianus IIPE453. Microbiol. Res. 2017, 200, 64–72. [Google Scholar] [CrossRef]
- Raimondi, S.; Zanni, E.; Amaretti, A.; Palleschi, C.; Uccelletti, D.; Rossi, M. Thermal adaptability of Kluyveromyces marxianus in recombinant protein production. Microb. Cell Factories 2013, 12, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Signori, L.; Passolunghi, S.; Ruohonen, L.; Porro, D.; Branduardi, P. Effect of oxygenation and temperature on glucose-xylose fermentation in Kluyveromyces marxianus CBS712 strain. Microb. Cell Factories 2014, 13, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoval-Nuñez, D.; Arellano-Plaza, M.; Gschaedler, A.; Arrizon, J.; Amaya-Delgado, L. A comparative study of lignocellulosic ethanol productivities by Kluyveromyces marxianus and Saccharomyces cerevisiae. Clean Technol. Environ. Policy 2017, 20, 1491–1499. [Google Scholar] [CrossRef]
- Pentjuss, A.; Stalidzans, E.; Liepins, J.; Kokina, A.; Martynova, J.; Zikmanis, P.; Mozga, I.; Scherbaka, R.; Hartman, H.; Poolman, M.G.; et al. Model-based biotechnological potential analysis of Kluyveromyces marxianus central metabolism. J. Ind. Microbiol. Biotechnol. 2017, 44, 1177–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Yuan, W.; Li, Y.; Xiang, R.; Hou, S.; Zhong, S.; Bai, F. Transcriptional analysis of Kluyveromyces marxianus for ethanol production from inulin using consolidated bioprocessing technology. Biotechnol. Biofuels 2015, 8, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varela, J.A.; Gethins, L.; Stanton, C.; Ross, P.; Morrissey, J.P. Applications of Kluyveromyces marxianus in Biotechnology; Springer Science and Business Media LLC: Berlin, Germany, 2017; pp. 439–453. [Google Scholar]
- Zhang, B.; Li, L.; Zhang, J.; Gao, X.; Wang, D.; Hong, J. Improving ethanol and xylitol fermentation at elevated temperature through substitution of xylose reductase in Kluyveromyces marxianus. J. Ind. Microbiol. Biotechnol. 2013, 40, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhang, L.; Wang, D.; Gao, X.; Hong, J. Identification of a xylose reductase gene in the xylose metabolic pathway of Kluyveromyces marxianus NBRC1777. J. Ind. Microbiol. Biotechnol. 2011, 38, 2001–2010. [Google Scholar] [CrossRef]
- Murata, Y.; Danjarean, H.; Fujimoto, K.; Kosugi, A.; Arai, T.; Ibrahim, W.A.; Suliman, O.; Hashim, R.; Mori, Y. Ethanol fermentation by the thermotolerant yeast, Kluyveromyces marxianus TISTR5925, of extracted sap from old oil palm trunk. Aims Energy 2015, 3, 201–213. [Google Scholar] [CrossRef]
- Tavares, B.; Felipe, M.D.G.D.A.; Dos Santos, J.C.; Pereira, F.M.; Gomes, S.D.; Sene, L. An experimental and modeling approach for ethanol production by Kluyveromyces marxianus in stirred tank bioreactor using vacuum extraction as a strategy to overcome product inhibition. Renew. Energy 2019, 131, 261–267. [Google Scholar] [CrossRef]
- Chi, Z.-M.; Zhang, T.; Cao, T.-S.; Liu, X.-Y.; Cui, W.; Zhao, C.-H. Biotechnological potential of inulin for bioprocesses. Bioresour. Technol. 2011, 102, 4295–4303. [Google Scholar] [CrossRef]
- Costa, D.A.; De Souza, C.J.A.; Costa, P.S.; Rodrigues, M.Q.R.B.; Dos Santos, A.F.; Lopes, M.R.; Geniêr, H.L.A.; Silveira, W.B.; Fietto, L.G. Physiological characterization of thermotolerant yeast for cellulosic ethanol production. Appl. Microbiol. Biotechnol. 2014, 98, 3829–3840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, W.; Wang, M.; Zhan, R.; Yu, Y.; He, Y.; Lv, H. Kluyveromyces marxianus developing ethanol tolerance during adaptive evolution with significant improvements of multiple pathways. Biotechnol. Biofuels 2019, 12, 63. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Li, P.; Zhang, L.; Li, S. Understanding the stress responses of Kluyveromyces marxianus after an arrest during high-temperature ethanol fermentation based on integration of RNA-Seq and metabolite data. Appl. Microbiol. Biotechnol. 2019, 103, 2715–2729. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, A.S.; Varela, J.A.; Juergens, H.; Daran, J.-M.G.; Morrissey, J.P. Biological Parts for Kluyveromyces marxianus Synthetic Biology. Front. Bioeng. Biotechnol. 2019, 7, 97. [Google Scholar] [CrossRef] [Green Version]
- Kurtzman, C.P.; Robnett, C.J.; Basehoar-Powers, E. Phylogenetic relationships among species of Pichia, Issatchenkia and Williopsis determined from multigene sequence analysis, and the proposal of Barnettozyma gen. nov., Lindnera gen. nov and Wickerhamomyces gen. nov. Fems Yeast Res. 2008, 8, 939–954. [Google Scholar] [CrossRef] [Green Version]
- Xiao, H.; Shao, Z.; Jiang, Y.; Dole, S.; Zhao, H. Exploiting Issatchenkia orientalis SD108 for succinic acid production. Microb. Cell Factories 2014, 13, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Koutinas, M.; Patsalou, M.; Stavrinou, S.; Vyrides, I. High temperature alcoholic fermentation of orange peel by the newly isolated thermotolerantPichia kudriavzeviiKVMP10. Lett. Appl. Microbiol. 2015, 62, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Kwon, Y.-J.; Ma, A.-Z.; Li, Q.; Wang, F.; Zhuang, G.; Liu, C.-Z. Effect of lignocellulosic inhibitory compounds on growth and ethanol fermentation of newly-isolated thermotolerant Issatchenkia orientalis. Bioresour. Technol. 2011, 102, 8099–8104. [Google Scholar] [CrossRef]
- Isono, N.; Hayakawa, H.; Usami, A.; Mishima, T.; Hisamatsu, M. A comparative study of ethanol production by Issatchenkia orientalis strains under stress conditions. J. Biosci. Bioeng. 2012, 113, 76–78. [Google Scholar] [CrossRef]
- Toivari, M.H.; Vehkomäki, M.-L.; Nygård, Y.; Penttilä, M.; Ruohonen, L.; Wiebe, M.G. Low pH d-xylonate production with Pichia kudriavzevii. Bioresour. Technol. 2013, 133, 555–562. [Google Scholar] [CrossRef] [Green Version]
- Boura, K.; Kandylis, P.; Bekatorou, A.; Dima, A.; Kanellaki, M.; Koutinas, A.A. Ester-Based Biofuels from Wastes. Sustain. Biotechnol. Enzymatic Resour. Renew. Energy 2018, 307–324. [Google Scholar] [CrossRef]
- Ekas, H.; Deaner, M.; Alper, H.S. Recent advancements in fungal-derived fuel and chemical production and commercialization. Curr. Opin. Biotechnol. 2019, 57, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Fatma, Z.; Song, X.; Hsieh, P.-H.; Tran, V.G.; Lyon, W.L.; Sayadi, M.; Shao, Z.; Yoshikuni, Y.; Zhao, H. A genetic toolbox for metabolic engineering of Issatchenkia orientalis. Metab. Eng. 2020, 59, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Tran, V.G.; Cao, M.; Fatma, Z.; Song, X.; Zhao, H. Development of a CRISPR/Cas9-Based Tool for Gene Deletion in Issatchenkia orientalis. mSphere 2019, 4, e00345-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siripong, W.; Wolf, P.; Kusumoputri, T.P.; Downes, J.J.; Kocharin, K.; Tanapongpipat, S.; Runguphan, W. Metabolic engineering of Pichia pastoris for production of isobutanol and isobutyl acetate. Biotechnol. Biofuels 2018, 11, 1. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Zhang, Z. Engineering strategies for enhanced production of protein and bio-products in Pichia pastoris: A review. Biotechnol. Adv. 2018, 36, 182–195. [Google Scholar] [CrossRef]
- Yang, Z.; Meng, X. Production of (2R, 3R)-2,3-butanediol using engineered Pichia pastoris: Strain construction, characterization and fermentation. Biotechnol. Biofuels 2018, 11, 35. [Google Scholar] [CrossRef]
- Hu, S.; Luo, X.; Wan, C.; Li, Y. Characterization of Crude Glycerol from Biodiesel Plants. J. Agric. Food Chem. 2012, 60, 5915–5921. [Google Scholar] [CrossRef]
- Luo, Z.; Miao, J.; Li, G.; Du, Y.; Yu, X. A Recombinant Highly Thermostable beta-Mannanase (ReTMan26) from Thermophilic Bacillus subtilis (TBS2) Expressed in Pichia pastoris and Its pH and Temperature Stability. Appl. Biochem. Biotechnol. 2017, 182, 1259–1275. [Google Scholar] [CrossRef]
- Hama, S.; Kondo, A. Enzymatic biodiesel production: An overview of potential feedstocks and process development. Bioresour. Technol. 2013, 135, 386–395. [Google Scholar] [CrossRef]
- Christopher, L.P.; Kumar, H.; Zambare, V.P. Enzymatic biodiesel: Challenges and opportunities. Appl. Energy 2014, 119, 497–520. [Google Scholar] [CrossRef]
- Hama, S.; Noda, H.; Kondo, A. How lipase technology contributes to evolution of biodiesel production using multiple feedstocks. Curr. Opin. Biotechnol. 2018, 50, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Huang, L.; Zheng, N.; Fu, Y.; Shan, M.; Xu, Z.; Ma, J.; Lu, F. Development of a Pichia pastoris whole-cell biocatalyst with overexpression of mutant lipase I PCLG47I from Penicillium cyclopium for biodiesel production. RSC Adv. 2018, 8, 26161–26168. [Google Scholar] [CrossRef] [Green Version]
- Hüttner, S.; Klaubauf, S.; De Vries, R.P.; Olsson, L. Characterisation of three fungal glucuronoyl esterases on glucuronic acid ester model compounds. Appl. Microbiol. Biotechnol. 2017, 101, 5301–5311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conacher, C.G.; García-Aparicio, M.P.; Coetzee, G.; Van Zyl, W.H.; Görgens, J.F. Scalable methanol-free production of recombinant glucuronoyl esterase in Pichia pastoris. BMC Res. Notes 2019, 12, 1–6. [Google Scholar] [CrossRef]
- Li, Y.-X.; Yi, P.; Liu, J.; Yan, Q.; Jiang, Z. High-level expression of an engineered β-mannanase (mRmMan5A) in Pichia pastoris for manno-oligosaccharide production using steam explosion pretreated palm kernel cake. Bioresour. Technol. 2018, 256, 30–37. [Google Scholar] [CrossRef]
- Fischer, J.; Glieder, A. Current advances in engineering tools for Pichia pastoris. Curr. Opin. Biotechnol. 2019, 59, 175–181. [Google Scholar] [CrossRef]
- Peng, B.; Plan, M.R.; Carpenter, A.; Nielsen, L.K.; Vickers, C.E. Coupling gene regulatory patterns to bioprocess conditions to optimize synthetic metabolic modules for improved sesquiterpene production in yeast. Biotechnol. Biofuels 2017, 10, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Carroll, D. Genome Engineering with Zinc-Finger Nucleases. Genetics 2011, 188, 773–782. [Google Scholar] [CrossRef] [Green Version]
- Reyon, D.; Khayter, C.; Regan, M.R.; Joung, J.K.; Sander, J.D. Engineering Designer Transcription Activator—Like Effector Nucleases (TALENs) by REAL or REAL-Fast Assembly. Curr. Protoc. Mol. Biol. 2012, 100, 12.15.1–12.15.14. [Google Scholar] [CrossRef]
- Aouida, M.; Li, L.; Mahjoub, A.; Alshareef, S.; Ali, Z.; Piatek, A.; Mahfouz, M.M. Transcription activator-like effector nucleases mediated metabolic engineering for enhanced fatty acids production in Saccharomyces cerevisiae. J. Biosci. Bioeng. 2015, 120, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Rigouin, C.; Gueroult, M.; Croux, C.; Dubois, G.; Borsenberger, V.; Barbe, S.; Marty, A.; Daboussi, F.; André, I.; Bordes, F. Production of Medium Chain Fatty Acids by Yarrowia lipolytica: Combining Molecular Design and TALEN to Engineer the Fatty Acid Synthase. ACS Synth. Biol. 2017, 6, 1870–1879. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liwei, M.; Park, J.-B.; Jeong, S.-H.; Wei, G.; Wang, Y.; Kim, S.-W. Microbial Platform for Terpenoid Production: Escherichia coli and Yeast. Front. Microbiol. 2018, 9, 2460. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Liang, L.; Choudhury, A.; Garst, A.D.; Eckert, C.A.; Oh, E.J.; Winkler, J.; Gill, R.T. Multiplex navigation of global regulatory networks (MINR) in yeast for improved ethanol tolerance and production. Metab. Eng. 2019, 51, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Zhao, E.M.; Zhang, Y.; Mehl, J.; Park, H.; Lalwani, M.A.; Toettcher, J.E.; Avalos, J.L. Optogenetic regulation of engineered cellular metabolism for microbial chemical production. Nat. Cell Biol. 2018, 555, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Vervoort, Y.; Linares, A.G.; Roncoroni, M.; Liu, C.; Steensels, J.; Verstrepen, K.J. High-throughput system-wide engineering and screening for microbial biotechnology. Curr. Opin. Biotechnol. 2017, 46, 120–125. [Google Scholar] [CrossRef]
- Gao, S.; Tong, Y.; Wen, Z.; Zhu, L.; Ge, M.; Chen, D.; Jiang, Y.; Yang, S. Multiplex gene editing of the Yarrowia lipolytica genome using the CRISPR-Cas9 system. J. Ind. Microbiol. Biotechnol. 2016, 43, 1085–1093. [Google Scholar] [CrossRef]
- Jacobs, J.Z.; Ciccaglione, K.M.; Tournier, V.; Zaratiegui, M. Implementation of the CRISPR-Cas9 system in fission yeast. Nat. Commun. 2014, 5, 1–5. [Google Scholar] [CrossRef]
- Schwartz, C.M.; Hussain, M.S.; Blenner, M.; Wheeldon, I. Synthetic RNA Polymerase III Promoters Facilitate High-Efficiency CRISPR–Cas9-Mediated Genome Editing in Yarrowia lipolytica. ACS Synth. Biol. 2016, 5, 356–359. [Google Scholar] [CrossRef]
- Löbs, A.-K.; Engel, R.; Schwartz, C.; Flores, A.; Wheeldon, I. CRISPR–Cas9-enabled genetic disruptions for understanding ethanol and ethyl acetate biosynthesis in Kluyveromyces marxianus. Biotechnol. Biofuels 2017, 10, 1–14. [Google Scholar] [CrossRef]
- Weninger, A.; Hatzl, A.-M.; Schmid, C.; Vogl, T.; Glieder, A. Combinatorial optimization of CRISPR/Cas9 expression enables precision genome engineering in the methylotrophic yeast Pichia pastoris. J. Biotechnol. 2016, 235, 139–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wehrs, M.; Tanjore, D.; Eng, T.; Lievense, J.; Pray, T.R.; Mukhopadhyay, A. Engineering Robust Production Microbes for Large-Scale Cultivation. Trends Microbiol. 2019, 27, 524–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeniran, A.; Sherer, M.; Tyo, K.E.J. Yeast-based biosensors: Design and applications. FEMS Yeast Res. 2014, 15, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumann, L.; Rajkumar, A.S.; Morrissey, J.P.; Boles, E.; Oreb, M. A Yeast-Based Biosensor for Screening of Short- and Medium-Chain Fatty Acid Production. ACS Synth. Biol. 2018, 7, 2640–2646. [Google Scholar] [CrossRef]
- Zhang, Y.; Hammer, S.K.; Carrasco-López, C.; Echauri, S.A.G.; Avalos, J.L. Genetically encoded biosensors for branched-chain amino acid metabolism to monitor mitochondrial and cytosolic production of isobutanol and isopentanol in yeast. BioRxiv 2020. [Google Scholar] [CrossRef]
- Thorwall, S.; Schwartz, C.; Chartron, J.W.; Wheeldon, I. Stress-tolerant non-conventional microbes enable next-generation chemical biosynthesis. Nat. Chem. Biol. 2020, 16, 113–121. [Google Scholar] [CrossRef]
- Vogl, T.; Hartner, F.S.; Glieder, A. New opportunities by synthetic biology for biopharmaceutical production in Pichia pastoris. Curr. Opin. Biotechnol. 2013, 24, 1094–1101. [Google Scholar] [CrossRef] [Green Version]
- Löbs, A.-K.; Schwartz, C.; Thorwall, S.; Wheeldon, I. Highly Multiplexed CRISPRi Repression of Respiratory Functions Enhances Mitochondrial Localized Ethyl Acetate Biosynthesis in Kluyveromyces marxianus. ACS Synth. Biol. 2018, 7, 2647–2655. [Google Scholar] [CrossRef]
- Schwartz, C.; Curtis, N.; Löbs, A.-K.; Wheeldon, I. Multiplexed CRISPR Activation of Cryptic Sugar Metabolism Enables Yarrowia Lipolytica Growth on Cellobiose. Biotechnol. J. 2018, 13, e1700584. [Google Scholar] [CrossRef]
- Zhang, X.-H.; Tee, L.Y.; Wang, X.-G.; Huang, Q.-S.; Yang, S.-H. Off-target Effects in CRISPR/Cas9-mediated Genome Engineering. Mol. Ther. Nucleic Acids 2015, 4, e264. [Google Scholar] [CrossRef]
- Stovicek, V.; Holkenbrink, C.; Borodina, I. CRISPR/Cas system for yeast genome engineering: Advances and applications. FEMS Yeast Res. 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Labun, K.; Montague, T.G.; Gagnon, J.A.; Thyme, S.B.; Valen, E. CHOPCHOP v2: A web tool for the next generation of CRISPR genome engineering. Nucleic Acids Res. 2016, 44, W272–W276. [Google Scholar] [CrossRef] [PubMed]
- Heigwer, F.; Kerr, G.; Boutros, M. E-CRISP: Fast CRISPR target site identification. Nat. Methods 2014, 11, 122–123. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.M.; Liang, L.L.; Freed, E.; Chang, H.; Oh, E.; Liu, Z.Y.; Garst, A.; Eckert, C.A.; Gill, R.T. Synthetic chimeric nucleases function for efficient genome editing. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Slaymaker, I.M.; Gao, L.; Zetsche, B.; Scott, D.A.; Yan, W.X.; Zhang, F. Faculty Opinions recommendation of Rationally engineered Cas9 nucleases with improved specificity. Faculty Opinions Post Publ. Peer Rev. Biomed. Lit. 2016, 351, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Martins-Santana, L.; Nora, L.C.; Sanches-Medeiros, A.; Lovate, G.L.; Cassiano, M.H.A.; Silva-Rocha, R. Systems and Synthetic Biology Approaches to Engineer Fungi for Fine Chemical Production. Front. Bioeng. Biotechnol. 2018, 6, 117. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, J. Systems Biology of Metabolism. Annu. Rev. Biochem. 2017, 86, 245–275. [Google Scholar] [CrossRef]
- Campbell, K.; Xia, J.; Nielsen, J. The Impact of Systems Biology on Bioprocessing. Trends Biotechnol. 2017, 35, 1156–1168. [Google Scholar] [CrossRef]
- Lu, H.; Li, F.; Sánchez, B.J.; Zhu, Z.; Li, G.; Domenzain, I.; Marcišauskas, S.; Anton, P.M.; Lappa, D.; Lieven, C.; et al. A consensus S. cerevisiae metabolic model Yeast8 and its ecosystem for comprehensively probing cellular metabolism. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lachance, J.-C.; Lloyd, C.J.; Monk, J.M.; Yang, L.; Sastry, A.V.; Seif, Y.; Palsson, B.O.; Rodrigue, S.; Feist, A.M.; King, Z.A.; et al. BOFdat: Generating biomass objective functions for genome-scale metabolic models from experimental data. PLoS Comput. Biol. 2019, 15, e1006971. [Google Scholar] [CrossRef]
- Chae, T.U.; Choi, S.Y.; Kim, J.W.; Ko, Y.-S.; Lee, S.Y. Recent advances in systems metabolic engineering tools and strategies. Curr. Opin. Biotechnol. 2017, 47, 67–82. [Google Scholar] [CrossRef] [PubMed]
- Loira, N.; Dulermo, T.; Nicaud, J.-M.; Sherman, D.J. A genome-scale metabolic model of the lipid-accumulating yeast Yarrowia lipolytica. BMC Syst. Biol. 2012, 6, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcišauskas, S.; Ji, B.; Nielsen, J. Reconstruction and analysis of a Kluyveromyces marxianus genome-scale metabolic model. BMC Bioinform. 2019, 20, 551–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caspeta, L.; Shoaie, S.; Agren, R.; Nookaew, I.; Nielsen, J. Genome-scale metabolic reconstructions of Pichia stipitis and Pichia pastoris and in-silico evaluation of their potentials. BMC Syst. Biol. 2012, 6, 24. [Google Scholar] [CrossRef] [Green Version]
- Pharkya, P.; Maranas, C.D. An optimization framework for identifying reaction activation/inhibition or elimination candidates for overproduction in microbial systems. Metab. Eng. 2006, 8, 1–13. [Google Scholar] [CrossRef]
- Ranganathan, R.; Lockless, S.W. Statistical Methods for Analyzing Biological Sequences. U.S. Patent 7,016,786, 21 March 2006. [Google Scholar]
- Castillo, S.; Patil, K.R.; Jouhten, P. Yeast Genome-Scale Metabolic Models for Simulating Genotype–Phenotype Relations. In Process Integration in Biochemical Engineering; Springer Science and Business Media LLC: Berlin, Germany, 2019; Volume 58, pp. 111–133. [Google Scholar]
- Choi, H.S.; Lee, S.Y.; Kim, T.Y.; Woo, H.M. In Silico Identification of Gene Amplification Targets for Improvement of Lycopene Production. Appl. Environ. Microbiol. 2010, 76, 3097–3105. [Google Scholar] [CrossRef] [Green Version]
- Medema, M.H.; Van Raaphorst, R.; Takano, E.; Breitling, R. Computational tools for the synthetic design of biochemical pathways. Nat. Rev. Genet. 2012, 10, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Fischer, C.R.; Klein-Marcuschamer, D.; Stephanopoulos, G. Selection and optimization of microbial hosts for biofuels production. Metab. Eng. 2008, 10, 295–304. [Google Scholar] [CrossRef]
- Young, E.; Lee, S.-M.; Alper, H. Optimizing pentose utilization in yeast: The need for novel tools and approaches. Biotechnol. Biofuels 2010, 3, 24. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.-C.; Kong, I.I.; Wei, N.; Peng, D.; Turner, T.L.; Sung, B.H.; Sohn, J.-H.; Jin, Y.-S. Optimization of an acetate reduction pathway for producing cellulosic ethanol by engineered yeast. Biotechnol. Bioeng. 2016, 113, 2587–2596. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Sahara, T.; Ohgiya, S.; Kamagata, Y.; Fujimori, K.E. Systematic optimization of gene expression of pentose phosphate pathway enhances ethanol production from a glucose/xylose mixed medium in a recombinant Saccharomyces cerevisiae. AMB Express 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jeschek, M.; Gerngross, D.; Panke, S. Combinatorial pathway optimization for streamlined metabolic engineering. Curr. Opin. Biotechnol. 2017, 47, 142–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mccarty, N.S.; Ledesma-Amaro, R. Synthetic Biology Tools to Engineer Microbial Communities for Biotechnology. Trends Biotechnol. 2019, 37, 181–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, K.R.; Jang, W.D.; Yang, D.; Cho, J.S.; Park, D.; Lee, S.Y. Systems Metabolic Engineering Strategies: Integrating Systems and Synthetic Biology with Metabolic Engineering. Trends Biotechnol. 2019, 37, 817–837. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Organism | Glucose (w/v) | Salt | Temp | Ethanol (v/v) | 5-HMF * (g/L) | pH |
|---|---|---|---|---|---|---|
| S. cerevisiae | 40% [32] | 0.4 M NaCl [33] | 39 °C [34] | 10% [35] | 3.0 g [36] | 3 [37] |
| Y. lipolytica | 30% [38] | 2 M NaCl [39] | 35 °C [40] | 5% [41] | 2.0 g [42] | 3.5 [43] |
| K. marxianus | 40% [44] | 1.5 M NaCl [45] | 52 °C [7] | 6% [46] | 3.84 g [47] | 3.5 [48] |
| P. pastoris | 40% [49] | 1.6 M NaCl [50] | 41 °C [51] | 6% [51] | - | 3 [52] |
| I. orientalis | 48% [7] | 0.85 M NaCl [7] | 45 °C [7] | 10% [53] | 7 g [7] | 2 [37] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lacerda, M.P.; Oh, E.J.; Eckert, C. The Model System Saccharomyces cerevisiae Versus Emerging Non-Model Yeasts for the Production of Biofuels. Life 2020, 10, 299. https://doi.org/10.3390/life10110299
Lacerda MP, Oh EJ, Eckert C. The Model System Saccharomyces cerevisiae Versus Emerging Non-Model Yeasts for the Production of Biofuels. Life. 2020; 10(11):299. https://doi.org/10.3390/life10110299
Chicago/Turabian StyleLacerda, Maria Priscila, Eun Joong Oh, and Carrie Eckert. 2020. "The Model System Saccharomyces cerevisiae Versus Emerging Non-Model Yeasts for the Production of Biofuels" Life 10, no. 11: 299. https://doi.org/10.3390/life10110299