Electroacupuncture on the Scalp over the Motor Cortex Ameliorates Behavioral Deficits Following Neonatal Hypoxia-Ischemia in Rats via the Activation of Neural Stem Cells

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Neonatal Hypoxia–Ischemia Model
2.3. Electroacupuncture Treatment
2.4. Brdu Labeling
2.5. Open Field Test
2.6. Cylinder Test
2.7. Rotarod Test
2.8. Passive Avoidance Test
2.9. Immunofluorescence Staining
2.10. Nissl Staining
2.11. Data Analyses
3. Results
3.1. Effects of EA Treatment on Motor and Memory Functions Following Hypoxia-Ischemia
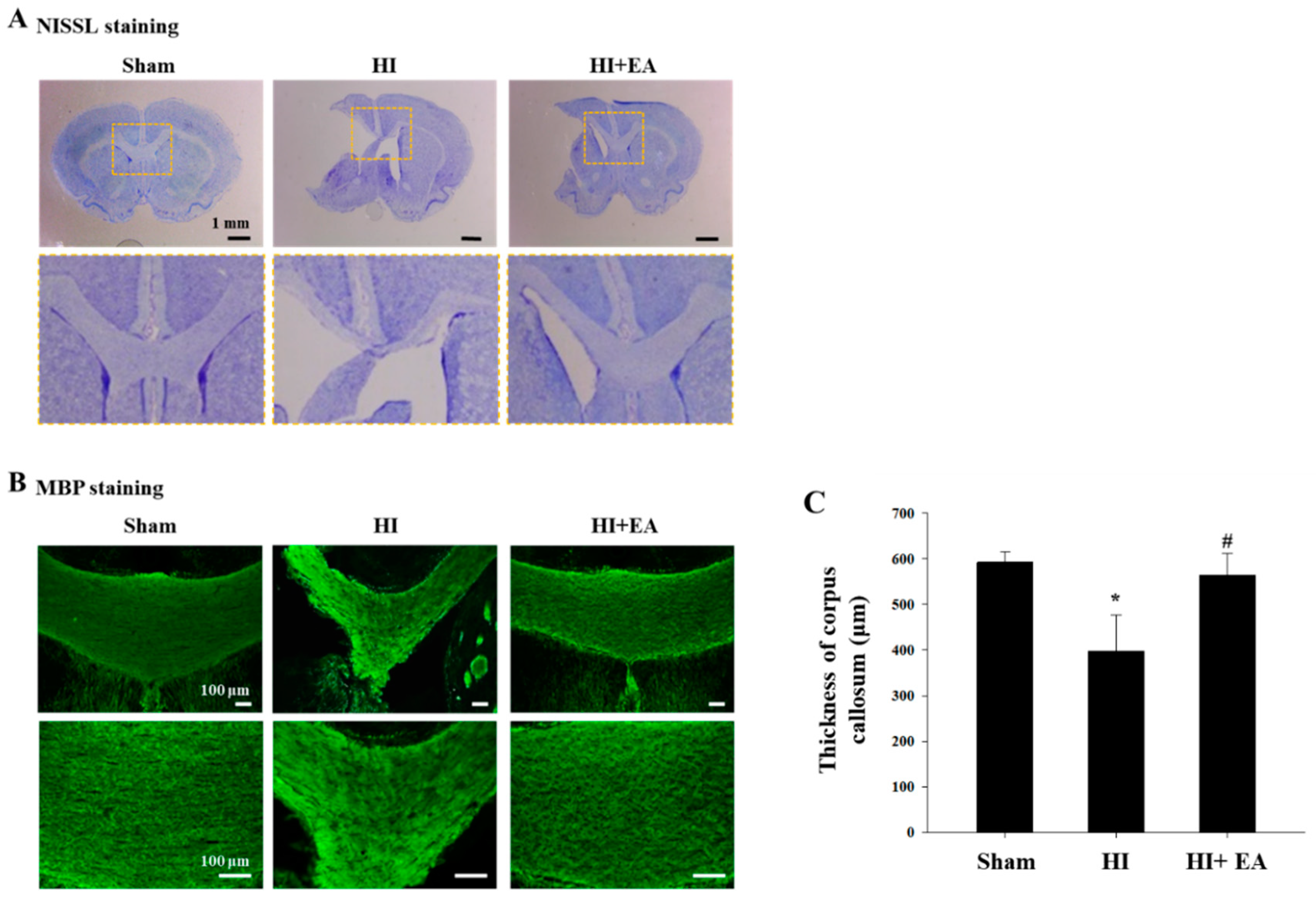
3.2. Effects of EA Treatment on the Corpus Callosum Thickness, Oligodendrocytes, and Astrocytes
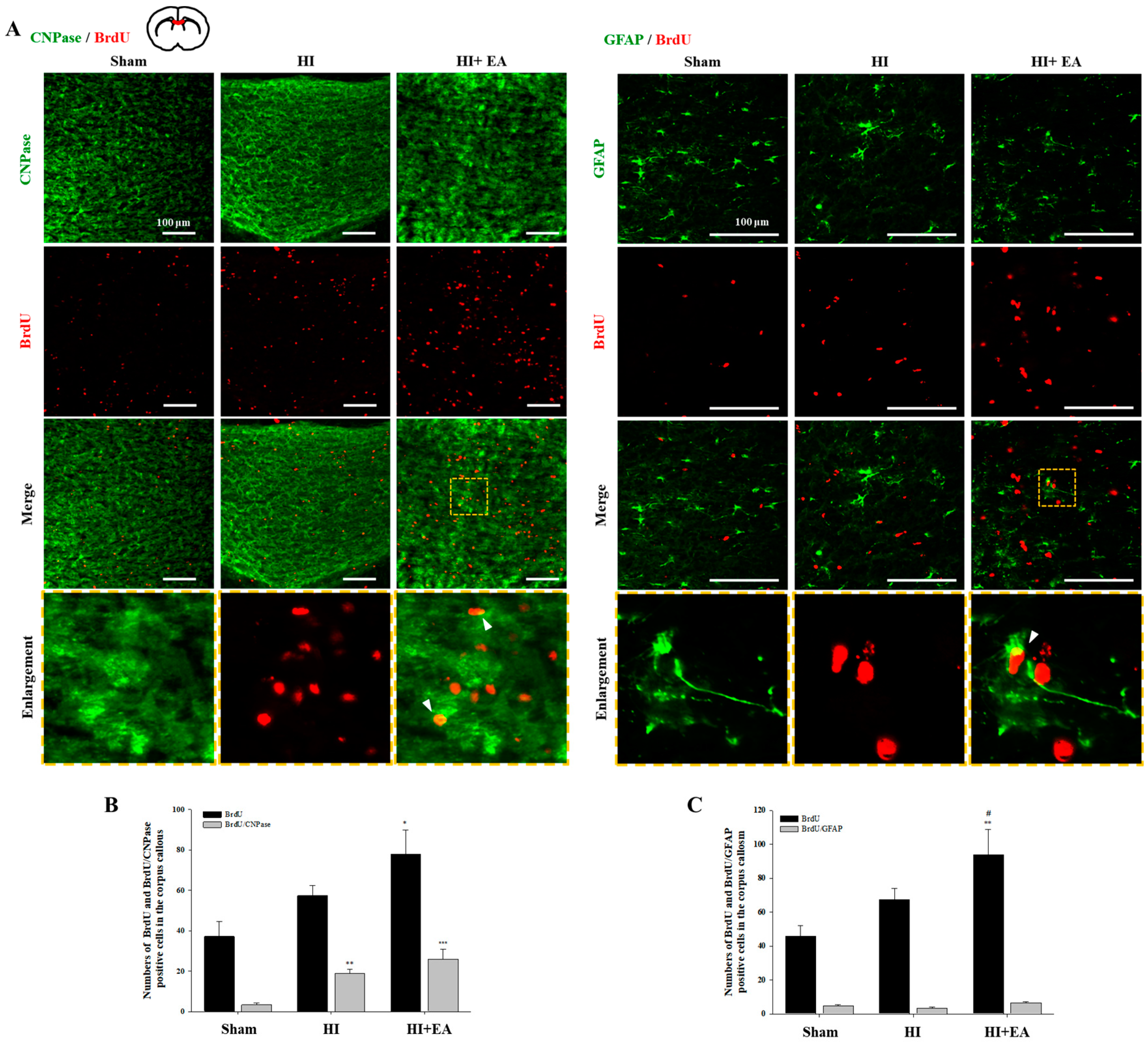
3.3. Effects of EA Treatment on Neurogenesis in the Subventricular Zone and Hippocampus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lawn, J.E.; Cousens, S.; Zupan, J. Lancet Neonatal Survival Steering, T. 4 million neonatal deaths: When? Where? Why? Lancet 2005, 365, 891–900. [Google Scholar] [CrossRef]
- Millar, L.J.; Shi, L.; Hoerder-Suabedissen, A.; Molnar, Z. Neonatal Hypoxia Ischaemia: Mechanisms, Models, and Therapeutic Challenges. Front. Cell. Neurosci. 2017, 11, 78. [Google Scholar] [CrossRef] [Green Version]
- Wyatt, J.S.; Gluckman, P.D.; Liu, P.Y.; Azzopardi, D.; Ballard, R.; Edwards, A.D.; Ferriero, D.M.; Polin, R.A.; Robertson, C.M.; Thoresen, M.; et al. Determinants of outcomes after head cooling for neonatal encephalopathy. Pediatrics 2007, 119, 912–921. [Google Scholar] [CrossRef]
- Follett, P.L.; Deng, W.; Dai, W.; Talos, D.M.; Massillon, L.J.; Rosenberg, P.A.; Volpe, J.J.; Jensen, F.E. Glutamate receptor-mediated oligodendrocyte toxicity in periventricular leukomalacia: A protective role for topiramate. J. Neurosci. 2004, 24, 4412–4420. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Lopez, D.; Natarajan, N.; Ashwal, S.; Vexler, Z.S. Mechanisms of perinatal arterial ischemic stroke. J. Cereb Blood Flow Metab. 2014, 34, 921–932. [Google Scholar] [CrossRef]
- Locatelli, A.; Incerti, M.; Ghidini, A.; Greco, M.; Villa, E.; Paterlini, G. Factors associated with umbilical artery acidemia in term infants with low Apgar scores at 5 min. Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 139, 146–150. [Google Scholar] [CrossRef]
- Spandou, E.; Papadopoulou, Z.; Soubasi, V.; Karkavelas, G.; Simeonidou, C.; Pazaiti, A.; Guiba-Tziampiri, O. Erythropoietin prevents long-term sensorimotor deficits and brain injury following neonatal hypoxia-ischemia in rats. Brain Res. 2005, 1045, 22–30. [Google Scholar] [CrossRef]
- Jones, M.W.; Morgan, E.; Shelton, J.E.; Thorogood, C. Cerebral palsy: Introduction and diagnosis (part I). J. Pediatr. Health Care 2007, 21, 146–152. [Google Scholar] [CrossRef]
- Miyamoto, N.; Maki, T.; Pham, L.D.; Hayakawa, K.; Seo, J.H.; Mandeville, E.T.; Mandeville, J.B.; Kim, K.W.; Lo, E.H.; Arai, K. Oxidative stress interferes with white matter renewal after prolonged cerebral hypoperfusion in mice. Stroke 2013, 44, 3516–3521. [Google Scholar] [CrossRef] [Green Version]
- Lindvall, O.; Kokaia, Z. Stem cell research in stroke: How far from the clinic? Stroke 2011, 42, 2369–2375. [Google Scholar] [CrossRef] [Green Version]
- Abe, K.; Yamashita, T.; Takizawa, S.; Kuroda, S.; Kinouchi, H.; Kawahara, N. Stem cell therapy for cerebral ischemia: From basic science to clinical applications. J. Cereb. Blood Flow Metab. 2012, 32, 1317–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arvidsson, A.; Collin, T.; Kirik, D.; Kokaia, Z.; Lindvall, O. Neuronal replacement from endogenous precursors in the adult brain after stroke. Nat. Med. 2002, 8, 963–970. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.M.; Li, D.; Zhang, S.H. Acupuncture for acute stroke. Cochrane Db Syst. Rev. 2018. [Google Scholar] [CrossRef] [PubMed]
- Pak, M.E.; Jung, D.H.; Lee, H.J.; Shin, M.J.; Kim, S.Y.; Shin, Y.B.; Yun, Y.J.; Shin, H.K.; Choi, B.T. Combined therapy involving electroacupuncture and treadmill exercise attenuates demyelination in the corpus callosum by stimulating oligodendrogenesis in a rat model of neonatal hypoxia-ischemia. Exp. Neurol. 2018, 300, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.R.; Kim, H.N.; Ahn, S.M.; Choi, Y.H.; Shin, H.K.; Choi, B.T. Electroacupuncture promotes post-stroke functional recovery via enhancing endogenous neurogenesis in mouse focal cerebral ischemia. PLoS ONE 2014, 9, e90000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Tao, J.; Lin, Y.; Lin, R.; Liu, W.; Chen, L. Electro-acupuncture exerts beneficial effects against cerebral ischemia and promotes the proliferation of neural progenitor cells in the cortical peri-infarct area through the Wnt/beta-catenin signaling pathway. Int. J. Mol. Med. 2015, 36, 1215–1222. [Google Scholar] [CrossRef] [Green Version]
- Ahn, S.M.; Kim, Y.R.; Kim, H.N.; Shin, Y.I.; Shin, H.K.; Choi, B.T. Electroacupuncture ameliorates memory impairments by enhancing oligodendrocyte regeneration in a mouse model of prolonged cerebral hypoperfusion. Sci. Rep. 2016, 6, 28646. [Google Scholar] [CrossRef]
- Kim, H.; Koo, Y.S.; Shin, M.J.; Kim, S.Y.; Shin, Y.B.; Choi, B.T.; Yun, Y.J.; Lee, S.Y.; Shin, H.K. Combination of Constraint-Induced Movement Therapy with Electroacupuncture Improves Functional Recovery following Neonatal Hypoxic-Ischemic Brain Injury in Rats. Biomed. Res. Int. 2018, 2018, 8638294. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Shen, J.; Wang, X.M.; Fu, D.L.; Chen, C.Y.; Lu, L.Y.; Lu, L.; Xie, C.L.; Fang, J.Q.; Zheng, G.Q. Scalp acupuncture for acute ischemic stroke: A meta-analysis of randomized controlled trials. Evid. Based Complement. Alternat. Med. 2012, 2012, 480950. [Google Scholar] [CrossRef] [Green Version]
- Han, J.Y.; Kim, J.H.; Park, J.H.; Song, M.Y.; Song, M.K.; Kim, D.J.; You, Y.N.; Park, G.C.; Choi, J.B.; Cho, M.R.; et al. Scalp acupuncture and electromagnetic convergence stimulation for patients with cerebral infarction: Study protocol for a randomized controlled trial. Trials 2016, 17, 490. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, S.J.; Cicchetti, F. Cellular and molecular mechanisms of action of transcranial direct current stimulation: Evidence from in vitro and in vivo models. Int. J. Neuropsychopharmacol. 2014, 18. [Google Scholar] [CrossRef] [PubMed]
- Rice, J.E., 3rd; Vannucci, R.C.; Brierley, J.B. The influence of immaturity on hypoxic-ischemic brain damage in the rat. Ann. Neurol. 1981, 9, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Radtke, C.; Sasaki, M.; Lankford, K.L.; Gallo, V.; Kocsis, J.D. CNPase expression in olfactory ensheathing cells. J. Biomed. Biotechnol. 2011, 2011, 608496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Sane, H.; Kulkarni, P.; D’sa, M.; Gokulchandran, N.; Badhe, P. Improved Quality of Life in A Case of Cerebral Palsy after Bone Marrow Mononuclear Cell Transplantation. Cell. J. 2015, 17, 389–394. [Google Scholar]
- Yager, J.Y. Animal models of hypoxic-ischemic brain damage in the newborn. Semin Pediatr. Neurol. 2004, 11, 31–46. [Google Scholar] [CrossRef]
- Aarnoudse-Moens, C.S.; Weisglas-Kuperus, N.; van Goudoever, J.B.; Oosterlaan, J. Meta-analysis of neurobehavioral outcomes in very preterm and/or very low birth weight children. Pediatrics 2009, 124, 717–728. [Google Scholar] [CrossRef]
- Silbereis, J.C.; Huang, E.J.; Back, S.A.; Rowitch, D.H. Towards improved animal models of neonatal white matter injury associated with cerebral palsy. Dis. Model. Mech. 2010, 3, 678–688. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Dietz, K.; DeLoyht, J.M.; Pedre, X.; Kelkar, D.; Kaur, J.; Vialou, V.; Lobo, M.K.; Dietz, D.M.; Nestler, E.J.; et al. Impaired adult myelination in the prefrontal cortex of socially isolated mice. Nat. Neurosci. 2012, 15, 1621–1623. [Google Scholar] [CrossRef] [Green Version]
- Palazuelos, J.; Crawford, H.C.; Klingener, M.; Sun, B.; Karelis, J.; Raines, E.W.; Aguirre, A. TACE/ADAM17 is essential for oligodendrocyte development and CNS myelination. J. Neurosci. 2014, 34, 11884–11896. [Google Scholar] [CrossRef]
- Verney, C.; Pogledic, I.; Biran, V.; Adle-Biassette, H.; Fallet-Bianco, C.; Gressens, P. Microglial reaction in axonal crossroads is a hallmark of noncystic periventricular white matter injury in very preterm infants. J. Neuropathol. Exp. Neurol. 2012, 71, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Nave, K.A. Myelination and support of axonal integrity by glia. Nature 2010, 468, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Zhang, J.; Jiang, H.; Wakana, S.; Poetscher, L.; Miller, M.I.; van Zijl, P.C.; Hillis, A.E.; Wytik, R.; Mori, S. DTI tractography based parcellation of white matter: Application to the mid-sagittal morphology of corpus callosum. Neuroimage 2005, 26, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Laporta-Hoyos, O.; Pannek, K.; Ballester-Plane, J.; Reid, L.B.; Vazquez, E.; Delgado, I.; Zubiaurre-Elorza, L.; Macaya, A.; Poo, P.; Melendez-Plumed, M.; et al. White matter integrity in dyskinetic cerebral palsy: Relationship with intelligence quotient and executive function. Neuroimage Clin. 2017, 15, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Bechler, M.E.; Ffrench-Constant, C. Neuroscience. A new wrap for neuronal activity? Science 2014, 344, 480–481. [Google Scholar] [CrossRef]
- Gibson, E.M.; Purger, D.; Mount, C.W.; Goldstein, A.K.; Lin, G.L.; Wood, L.S.; Inema, I.; Miller, S.E.; Bieri, G.; Zuchero, J.B.; et al. Neuronal activity promotes oligodendrogenesis and adaptive myelination in the mammalian brain. Science 2014, 344, 1252304. [Google Scholar] [CrossRef] [Green Version]
- Menn, B.; Garcia-Verdugo, J.M.; Yaschine, C.; Gonzalez-Perez, O.; Rowitch, D.; Alvarez-Buylla, A. Origin of oligodendrocytes in the subventricular zone of the adult brain. J. Neurosci. 2006, 26, 7907–7918. [Google Scholar] [CrossRef]
- Gage, F.H. Mammalian neural stem cells. Science 2000, 287, 1433–1438. [Google Scholar] [CrossRef]
- Barkho, B.Z.; Song, H.; Aimone, J.B.; Smrt, R.D.; Kuwabara, T.; Nakashima, K.; Gage, F.H.; Zhao, X. Identification of astrocyte-expressed factors that modulate neural stem/progenitor cell differentiation. Stem Cells Dev. 2006, 15, 407–421. [Google Scholar] [CrossRef] [Green Version]
- Rusznak, Z.; Henskens, W.; Schofield, E.; Kim, W.S.; Fu, Y. Adult Neurogenesis and Gliogenesis: Possible Mechanisms for Neurorestoration. Exp. Neurobiol. 2016, 25, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.W.; Xie, C.L.; Lu, L.; Zheng, G.Q. A systematic review and meta-analysis of Baihui (GV20)-based scalp acupuncture in experimental ischemic stroke. Sci. Rep. 2014, 4, 3981. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.R.; Kim, H.N.; Jang, J.Y.; Park, C.; Lee, J.H.; Shin, H.K.; Choi, Y.H.; Choi, B.T. Effects of electroacupuncture on apoptotic pathways in a rat model of focal cerebral ischemia. Int. J. Mol. Med. 2013, 32, 1303–1310. [Google Scholar] [CrossRef]
- Zhu, K.; Sun, J.; Kang, Z.; Zou, Z.; Wu, G.; Wang, J. Electroacupuncture Promotes Remyelination after Cuprizone Treatment by Enhancing Myelin Debris Clearance. Front. Neurosci. 2016, 10, 613. [Google Scholar] [CrossRef]
- Braun, R.; Klein, R.; Walter, H.L.; Ohren, M.; Freudenmacher, L.; Getachew, K.; Ladwig, A.; Luelling, J.; Neumaier, B.; Endepols, H.; et al. Transcranial direct current stimulation accelerates recovery of function, induces neurogenesis and recruits oligodendrocyte precursors in a rat model of stroke. Exp. Neurol. 2016, 279, 127–136. [Google Scholar] [CrossRef]
- Li, L.; El-Hayek, Y.H.; Liu, B.; Chen, Y.; Gomez, E.; Wu, X.; Ning, K.; Li, L.; Chang, N.; Zhang, L.; et al. Direct-current electrical field guides neuronal stem/progenitor cell migration. Stem Cells 2008, 26, 2193–2200. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, B.; Zhang, G.; Wang, J.; Tian, W.; Ju, G.; Wei, X.; Song, B. Electric signals regulate directional migration of ventral midbrain derived dopaminergic neural progenitor cells via Wnt/GSK3beta signaling. Exp. Neurol. 2015, 263, 113–121. [Google Scholar] [CrossRef]
- Kobelt, L.J.; Wilkinson, A.E.; McCormick, A.M.; Willits, R.K.; Leipzig, N.D. Short duration electrical stimulation to enhance neurite outgrowth and maturation of adult neural stem progenitor cells. Ann. Biomed. Eng. 2014, 42, 2164–2176. [Google Scholar] [CrossRef]
- Pikhovych, A.; Stolberg, N.P.; Jessica Flitsch, L.; Walter, H.L.; Graf, R.; Fink, G.R.; Schroeter, M.; Rueger, M.A. Transcranial Direct Current Stimulation Modulates Neurogenesis and Microglia Activation in the Mouse Brain. Stem Cells Int. 2016, 2016, 2715196. [Google Scholar] [CrossRef] [Green Version]
- Liebetanz, D.; Nitsche, M.A.; Tergau, F.; Paulus, W. Pharmacological approach to the mechanisms of transcranial DC-stimulation-induced after-effects of human motor cortex excitability. Brain 2002, 125, 2238–2247. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.K.; Lee, S.W.; Choi, B.T. Modulation of neurogenesis via neurotrophic factors in acupuncture treatments for neurological diseases. Biochem. Pharmacol. 2017, 141, 132–142. [Google Scholar] [CrossRef]
- Alam, M.A.; Subramanyam Rallabandi, V.P.; Roy, P.K. Systems Biology of Immunomodulation for Post-Stroke Neuroplasticity: Multimodal Implications of Pharmacotherapy and Neurorehabilitation. Front. Neurol. 2016, 7, 94. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.R.; Ahn, S.M.; Pak, M.E.; Lee, H.J.; Jung, D.H.; Shin, Y.I.; Shin, H.K.; Choi, B.T. Potential benefits of mesenchymal stem cells and electroacupuncture on the trophic factors associated with neurogenesis in mice with ischemic stroke. Sci. Rep. 2018, 8, 2044. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, D.H.; Pak, M.E.; Lee, H.J.; Ahn, S.M.; Yun, Y.J.; Shin, Y.-I.; Shin, H.K.; Lee, S.-Y.; Choi, B.T. Electroacupuncture on the Scalp over the Motor Cortex Ameliorates Behavioral Deficits Following Neonatal Hypoxia-Ischemia in Rats via the Activation of Neural Stem Cells. Life 2020, 10, 240. https://doi.org/10.3390/life10100240
Jung DH, Pak ME, Lee HJ, Ahn SM, Yun YJ, Shin Y-I, Shin HK, Lee S-Y, Choi BT. Electroacupuncture on the Scalp over the Motor Cortex Ameliorates Behavioral Deficits Following Neonatal Hypoxia-Ischemia in Rats via the Activation of Neural Stem Cells. Life. 2020; 10(10):240. https://doi.org/10.3390/life10100240
Chicago/Turabian StyleJung, Da Hee, Malk Eun Pak, Hong Ju Lee, Sung Min Ahn, Young Ju Yun, Yong-Il Shin, Hwa Kyoung Shin, Seo-Yeon Lee, and Byung Tae Choi. 2020. "Electroacupuncture on the Scalp over the Motor Cortex Ameliorates Behavioral Deficits Following Neonatal Hypoxia-Ischemia in Rats via the Activation of Neural Stem Cells" Life 10, no. 10: 240. https://doi.org/10.3390/life10100240