The Impact of a New Interleukin-2-Based Immunotherapy Candidate on Urothelial Cells to Support Use for Intravesical Drug Delivery

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Interleukin-2 and Its Diverse Potential
1.2. IL-2 Influencing Tissue Cells
1.3. IL-2 in Cancer Therapy
1.4. Bladder Cancer
1.5. Rational to Develop an Innovative Interleukin-2 Candidate for Intravesical Use
2. Materials and Methods
2.1. Isolation and Cultivation of Cells
2.2. Used Interleukin-2 Types
2.3. Cell Proliferation Assays (WST-Assay)
2.4. Apoptosis and Necrosis Analysis
2.5. CD25 Analysis
3. Results
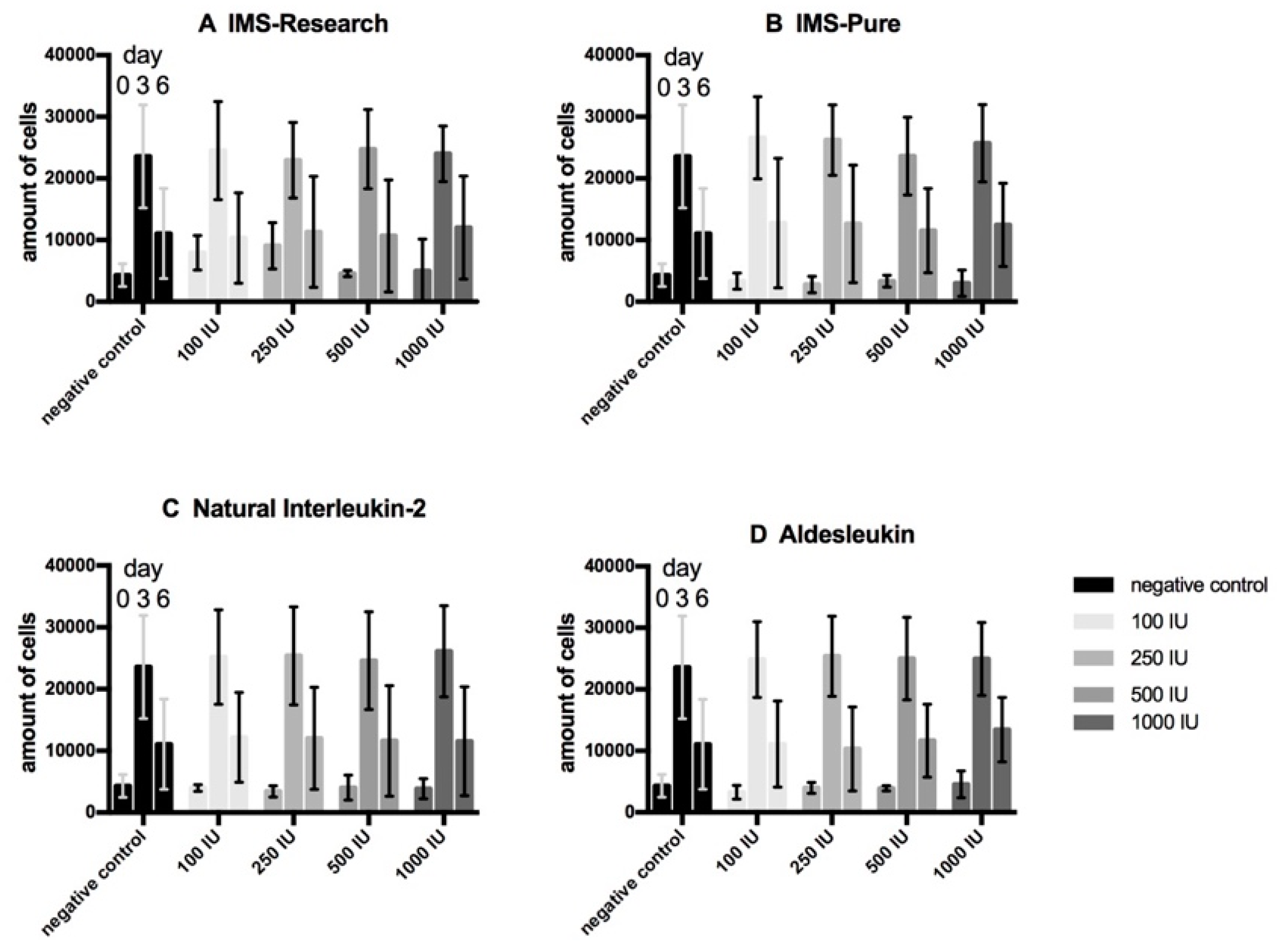
3.1. Cell Proliferation Assays (WST-Assay)
3.2. Detection of Apoptotic and Necrotic Processes
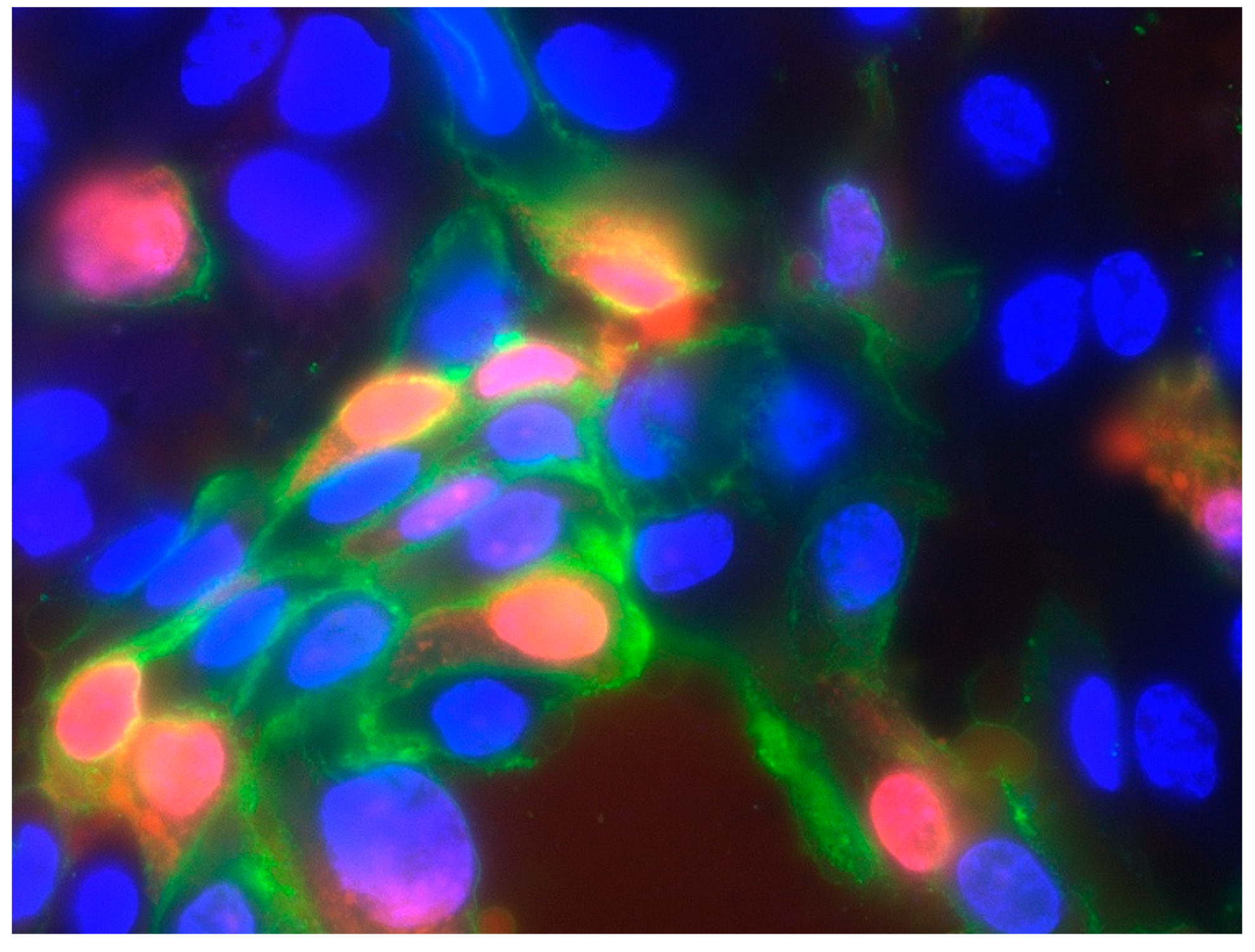
3.3. Detection of the Alpha Subunit of the IL-2 Receptor (CD 25)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Ethics Approval
Abbreviations
| BCG | Bacillus Calmette-Guerin |
| CHO | Chinese hamster ovarian |
| CO2 | Carbon dioxide |
| CTLL | Cytotoxic T lymphocyte |
| DMEM | Dulbeccos modified Eagles medium |
| E.coliFCS | Escherichia coli |
| IFN | Fetal calf serum |
| IL-2 | Interferon |
| Interleukin-2 | |
| IL-2R | Interleukin-2 receptor |
| IU | International units |
| MAPK | Mitogen-activated protein kinase |
| NMIBC | Non muscle-invasive bladder cancer |
| Pi3K | Phosphatidylinositol-3-kinase |
| P/S | Penicillin/streptomycin |
| PBS | Phosphate buffered saline |
| REM | Renal epithelial medium |
| RPMI | Roswell park memorial institute |
| SmPC | Summary of Product Characteristics |
| TNFa | Tumor necrosis factor alpha |
| WST | Water soluble tetrazolium |
References
- Morgan, D.A.; Ruscetti, F.W.; Gallo, R. Selective in vitro growth of T lymphocytes from normal human bone marrows. Science 1976, 193, 1007–1008. [Google Scholar] [CrossRef] [PubMed]
- Olejniczak, K.; Kasprzak, A. Biological properties of interleukin 2 and its role in pathogenesis of selected diseases—A review. Med. Sci. Monit. 2008, 14, RA179–RA189. [Google Scholar]
- Gaffen, S.L.; Liu, K.D. Overview of interleukin-2 function, production and clinical applications. Cytokine 2004, 28, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Lorre, K.; Fransen, L.; Ceuppens, J.L. Interleukin-2 induces tumor necrosis factor-alpha production by activated human T cells via a cyclosporin-sensitive pathway. Eur. Cytokine Netw. 1992, 3, 321–330. [Google Scholar] [PubMed]
- Bachmann, M.F.; Oxenius, A. Interleukin 2: From immunostimulation to immunoregulation and back again. EMBO Rep. 2007, 8, 1142–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, B.H.; Willerford, D.M. Biology of the interleukin-2 receptor. Adv. Immunol. 1998, 70, 1–81. [Google Scholar] [PubMed]
- Doersch, K.M.; DelloStritto, D.J.; Newell-Rogers, M.K. The contribution of interleukin-2 to effective wound healing. Exp. Biol. Med. (Maywood) 2017, 242, 384–396. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Rickert, M.; Garcia, K.C. Structure of the quaternary complex of interleukin-2 with its alpha, beta, and gammac receptors. Science 2005, 310, 1159–1163. [Google Scholar] [CrossRef]
- Ahmadzadeh, M.; Rosenberg, S.A. IL-2 administration increases CD4+ CD25(hi) Foxp3+ regulatory T cells in cancer patients. Blood 2006, 107, 2409–2414. [Google Scholar] [CrossRef] [Green Version]
- Antony, P.A.; Piccirillo, C.A.; Akpinarli, A.; Finkelstein, S.E.; Speiss, P.J.; Surman, D.R.; Palmer, D.C.; Chan, C.C.; Klebanoff, C.A.; Overwijk, W.W.; et al. CD8+ T cell immunity against a tumor/self-antigen is augmented by CD4+ T helper cells and hindered by naturally occurring T regulatory cells. J. Immunol. 2005, 174, 2591–2601. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, D.; Adams, J.A.; McCarthy, D.; Barrett, A.J. Interleukin-2 inhibits growth of fibroblasts derived from human bone marrow. Acta Haematol. 1990, 83, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Gerritsma, J.S.; Gerritsen, A.F.; van Kooten, C.; van Es, L.A.; Daha, M.R. Expression of the IL-2 receptor on human renal proximal tubular epithelial cells. J. Am. Soc. Nephrol. 1997, 8, 1510–1516. [Google Scholar] [PubMed]
- Eguizabal, C.; Boyano, M.D.; Diez-Torre, A.; Andrade, R.; Andollo, N.; De Felici, M.; Arechaga, J. Interleukin-2 induces the proliferation of mouse primordial germ cells in vitro. Int. J. Dev. Biol. 2007, 51, 731–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.J.; Luo, Q.L.; Zhang, M.X. Effects of interleukin on orbital fibroblasts of patients with Graves’ ophthalmopathy. Sichuan Da Xue Xue Bao Yi Xue Ban 2004, 35, 241–243. [Google Scholar]
- Corrigall, V.M.; Arastu, M.; Khan, S.; Shah, C.; Fife, M.; Smeets, T.; Tak, P.P.; Panayi, G.S. Functional IL-2 receptor beta (CD122) and gamma (CD132) chains are expressed by fibroblast-like synoviocytes: Activation by IL-2 stimulates monocyte chemoattractant protein-1 production. J. Immunol. 2001, 166, 4141–4147. [Google Scholar] [CrossRef] [Green Version]
- Ozawa, A.; Tada, H.; Sugawara, Y.; Uehara, A.; Sasano, T.; Shimauchi, H.; Takada, H.; Sugawara, S. Endogenous IL-15 sustains recruitment of IL-2Rbeta and common gamma and IL-2-mediated chemokine production in normal and inflamed human gingival fibroblasts. J. Immunol. 2004, 173, 5180–5188. [Google Scholar] [CrossRef]
- Ozawa, A.; Tada, H.; Tamai, R.; Uehara, A.; Watanabe, K.; Yamaguchi, T.; Shimauchi, H.; Takada, H.; Sugawara, S. Expression of IL-2 receptor beta and gamma chains by human gingival fibroblasts and up-regulation of adhesion to neutrophils in response to IL-2. J. Leukoc. Biol. 2003, 74, 352–359. [Google Scholar] [CrossRef]
- Gruss, H.J.; Scott, C.; Rollins, B.J.; Brach, M.A.; Herrmann, F. Human fibroblasts express functional IL-2 receptors formed by the IL-2R alpha- and beta-chain subunits: Association of IL-2 binding with secretion of the monocyte chemoattractant protein-1. J. Immunol. 1996, 157, 851–857. [Google Scholar]
- Mishra, J.; Waters, C.M.; Kumar, N. Molecular mechanism of interleukin-2-induced mucosal homeostasis. Am. J. Physiol. Cell Physiol. 2012, 302, C735–C747. [Google Scholar] [CrossRef]
- Den Otter, W.; Jacobs, J.J.; Battermann, J.J.; Hordijk, G.J.; Krastev, Z.; Moiseeva, E.V.; Stewart, R.J.; Ziekman, P.G.; Koten, J.W. Local therapy of cancer with free IL-2. Cancer Immunol. Immunother. 2008, 57, 931–950. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, S.A.; Lotze, M.T.; Muul, L.M.; Leitman, S.; Chang, A.E.; Ettinghausen, S.E.; Matory, Y.L.; Skibber, J.M.; Shiloni, E.; Vetto, J.T.; et al. Observations on the systemic administration of autologous lymphokine-activated killer cells and recombinant interleukin-2 to patients with metastatic cancer. N. Engl. J. Med. 1985, 313, 1485–1492. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, S.A. IL-2: The first effective immunotherapy for human cancer. J. Immunol. 2014, 192, 5451–5458. [Google Scholar] [CrossRef] [PubMed]
- Siegel, J.P.; Puri, R.K. Interleukin-2 toxicity. J. Clin. Oncol. 1991, 9, 694–704. [Google Scholar] [CrossRef] [PubMed]
- Boyman, O.; Sprent, J. The role of interleukin-2 during homeostasis and activation of the immune system. Nat. Rev. Immunol. 2012, 12, 180–190. [Google Scholar] [CrossRef]
- Donohue, J.H.; Rosenberg, S.A. The fate of interleukin-2 after in vivo administration. J. Immunol. 1983, 130, 2203–2208. [Google Scholar]
- Rosenstein, M.; Ettinghausen, S.E.; Rosenberg, S.A. Extravasation of intravascular fluid mediated by the systemic administration of recombinant interleukin 2. J. Immunol. 1986, 137, 1735–1742. [Google Scholar]
- Huland, E.; Heinzer, H. Renal cell carcinoma-innovative medical treatments. Curr. Opin. Urol. 2004, 14, 239–244. [Google Scholar] [CrossRef]
- Takezawa, Y.; Izumi, K.; Shimura, Y.; Aerken, M.; Natsagdorji, A.; Iijima, M.; Shigehara, K.; Nohara, T.; Narimoto, K.; Kadono, Y.; et al. Treatment Outcome of Low-dose Interleukin-2 Therapy in Patients with Metastatic Renal Cell Carcinoma. Anticancer Res. 2016, 36, 4961–4964. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.H.; Kim, M.S.; Chung, H.S.; Hwang, E.C. Novel immunotherapy in metastatic renal cell carcinoma. Investig. Clin. Urol. 2017, 58, 220–227. [Google Scholar] [CrossRef]
- Fishman, M.; Dutcher, J.P.; Clark, J.I.; Alva, A.; Miletello, G.P.; Curti, B.; Agarwal, N.; Hauke, R.; Mahoney, K.M.; Moon, H.; et al. Overall survival by clinical risk category for high dose interleukin-2 (HD IL-2) treated patients with metastatic renal cell cancer (mRCC): Data from the PROCLAIM(SM) registry. J. Immunother. Cancer 2019, 7, 84. [Google Scholar] [CrossRef] [Green Version]
- Sanlorenzo, M.; Vujic, I.; Posch, C.; Dajee, A.; Yen, A.; Kim, S.; Ashworth, M.; Rosenblum, M.D.; Algazi, A.; Osella-Abate, S.; et al. Melanoma immunotherapy. Cancer Biol. Ther. 2014, 15, 665–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastholt, L.; Svane, I.M.; Bjerregaard, J.K.; Herrstedt, J.; Hrobjartsson, A.; Schmidt, H. High-dose interleukin-2 and interferon as first-line immunotherapy for metastatic melanoma: Long-term follow-up in a large unselected Danish patient cohort. Eur. J. Cancer 2019, 115, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Dutcher, J.P.; Schwartzentruber, D.J.; Kaufman, H.L.; Agarwala, S.S.; Tarhini, A.A.; Lowder, J.N.; Atkins, M.B. High dose interleukin-2 (Aldesleukin)-expert consensus on best management practices-2014. J. Immunother. Cancer 2014, 2, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noble, S.; Goa, K.L. Aldesleukin (recombinant interleukin-2). BioDrugs 1997, 7, 394–422. [Google Scholar] [CrossRef]
- Melero, I.A.E.; Mau Sorensen, M.; Lassen, U.; Lolkema, M.; Robbrecht, D.G.; Gomez-Roca, C.A.; Martin-Liberal, J.; Tabernero, J.; Ros, W.; Ahmed, S.; et al. 412PDClinical activity, safety, and PK/PD from a phase I study of RO6874281, a fibroblast activation protein (FAP) targeted interleukin-2 variant (IL-2v). Ann. Oncol. 2018, 29. [Google Scholar] [CrossRef]
- Burger, M.; Catto, J.W.; Dalbagni, G.; Grossman, H.B.; Herr, H.; Karakiewicz, P.; Kassouf, W.; Kiemeney, L.A.; La Vecchia, C.; Shariat, S.; et al. Epidemiology and risk factors of urothelial bladder cancer. Eur. Urol. 2013, 63, 234–241. [Google Scholar] [CrossRef]
- Ploeg, M.; Aben, K.K.; Kiemeney, L.A. The present and future burden of urinary bladder cancer in the world. World J. Urol. 2009, 27, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Sylvester, R.J.; van der Meijden, A.P.; Oosterlinck, W.; Witjes, J.A.; Bouffioux, C.; Denis, L.; Newling, D.W.; Kurth, K. Predicting recurrence and progression in individual patients with stage Ta T1 bladder cancer using EORTC risk tables: A combined analysis of 2596 patients from seven EORTC trials. Eur. Urol. 2006, 49, 466–477. [Google Scholar] [CrossRef]
- Altay, B.; Girgin, C.; Kefi, A.; Cikili, N. The best management of superficial bladder tumours: Comparing TUR alone versus TUR combined with intravesical chemotherapy modalities? Int. Urol. Nephrol. 2000, 32, 53–58. [Google Scholar] [CrossRef]
- Kandoth, C.; McLellan, M.D.; Vandin, F.; Ye, K.; Niu, B.; Lu, C.; Xie, M.; Zhang, Q.; McMichael, J.F.; Wyczalkowski, M.A.; et al. Mutational landscape and significance across 12 major cancer types. Nature 2013, 502, 333–339. [Google Scholar] [CrossRef] [Green Version]
- Horiguchi, Y.; Larchian, W.A.; Kaplinsky, R.; Fair, W.R.; Heston, W.D. Intravesical liposome-mediated interleukin-2 gene therapy in orthotopic murine bladder cancer model. Gene Ther. 2000, 7, 844–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tubaro, A.; Stoppacciaro, A.; Velotti, F.; Bossola, P.C.; Cusumano, G.; Vicentini, C.; De Carli, P.; Ruco, L.; Santoni, A.; Cancrini, A.; et al. Local immunotherapy of superficial bladder cancer by intravesical instillation of recombinant interleukin-2. Eur. Urol. 1995, 28, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Ferlazzo, G.; Magno, C.; Lupo, G.; Rizzo, M.; Iemmo, R.; Semino, C.; Melioli, G. A phase I study of intravesical continuous perfusion of recombinant interleukin-2 in patients with superficial bladder cancer. Am. J. Clin. Oncol. 1995, 18, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, B.; Moy, R.L. Effect of perilesional injections of PEG-interleukin-2 on basal cell carcinoma. Dermatol. Surg. 2000, 26, 1037–1040. [Google Scholar] [CrossRef]
- Pizza, G.; Severini, G.; Menniti, D.; De Vinci, C.; Corrado, F. Tumour regression after intralesional injection of interleukin 2 (IL-2) in bladder cancer. Preliminary report. Int. J. Cancer 1984, 34, 359–367. [Google Scholar] [CrossRef]
- Huland, E.; Huland, H. Local continuous high dose interleukin 2: A new therapeutic model for the treatment of advanced bladder carcinoma. Cancer Res. 1989, 49, 5469–5474. [Google Scholar]
- Morales, A.; Eidinger, D.; Bruce, A.W. Intracavitary Bacillus Calmette-Guerin in the treatment of superficial bladder tumors. J. Urol. 1976, 116, 180–183. [Google Scholar] [CrossRef]
- Babjuk, M.; Burger, M.; Comperat, E.M.; Gontero, P.; Mostafid, A.H.; Palou, J.; van Rhijn, B.W.G.; Roupret, M.; Shariat, S.F.; Sylvester, R.; et al. European Association of Urology Guidelines on Non-muscle-invasive Bladder Cancer (TaT1 and Carcinoma in Situ)—2019 Update. Eur. Urol. 2019, 76, 639–657. [Google Scholar] [CrossRef]
- Chang, S.S.; Boorjian, S.A.; Chou, R.; Clark, P.E.; Daneshmand, S.; Konety, B.R.; Pruthi, R.; Quale, D.Z.; Ritch, C.R.; Seigne, J.D.; et al. Diagnosis and Treatment of Non-Muscle Invasive Bladder Cancer: AUA/SUO Guideline. J. Urol. 2016, 196, 1021–1029. [Google Scholar] [CrossRef]
- Woldu, S.L.; Bagrodia, A.; Lotan, Y. Guideline of guidelines: Non-muscle-invasive bladder cancer. BJU Int. 2017, 119, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Peyton, C.C.; Chipollini, J.; Azizi, M.; Kamat, A.M.; Gilbert, S.M.; Spiess, P.E. Updates on the use of intravesical therapies for non-muscle invasive bladder cancer: How, when and what. World J. Urol. 2019, 37, 2017–2029. [Google Scholar] [CrossRef] [PubMed]
- Proleukin 18 × 106 IU Powder for Solution for Injection or Infusion. Available online: https://www.medicines.org.uk/emc/product/291/smpc/print (accessed on 14 September 2020).
- Villoldo, G.M.; Loresi, M.; Giudice, C.; Damia, O.; Moldes, J.M.; DeBadiola, F.; Barbich, M.; Argibay, P. Histologic changes after urethroplasty using small intestinal submucosa unseeded with cells in rabbits with injured urethra. Urology 2013, 81, 1380.e1–1380.e5. [Google Scholar] [CrossRef] [PubMed]
- Besser, M.J.; Schallmach, E.; Oved, K.; Treves, A.J.; Markel, G.; Reiter, Y.; Schachter, J. Modifying interleukin-2 concentrations during culture improves function of T cells for adoptive immunotherapy. Cytotherapy 2009, 11, 206–217. [Google Scholar] [CrossRef] [PubMed]
- El-Assmy, A.; El-Hamid, M.A.; Hafez, A.T. Urethral replacement: A comparison between small intestinal submucosa grafts and spontaneous regeneration. BJU Int. 2004, 94, 1132–1135. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, P.; Carroll, K.L.; Berceli, S.A.; Barnhill, S.; Wrenshall, L.E. Expression of a Functional IL-2 Receptor in Vascular Smooth Muscle Cells. J. Immunol. 2019, 202, 694–703. [Google Scholar] [CrossRef] [Green Version]
- Conradt, H.S.; Ausmeier, M.; Dittmar, K.E.; Hauser, H.; Lindenmaier, W. Secretion of glycosylated human interleukin-2 by recombinant mammalian cell lines. Carbohydr. Res. 1986, 149, 443–450. [Google Scholar] [CrossRef]
- Courtney, L.P.; Phelps, J.L.; Karavodin, L.M. An anti-IL-2 antibody increases serum half-life and improves anti-tumor efficacy of human recombinant interleukin-2. Immunopharmacology 1994, 28, 223–232. [Google Scholar] [CrossRef]
- Zwietering, M.H.; Jongenburger, I.; Rombouts, F.M.; van’t Riet, K. Modeling of the bacterial growth curve. Appl. Environ. Microbiol. 1990, 56, 1875–1881. [Google Scholar] [CrossRef] [Green Version]
- Martz, E.; Steinberg, M.S. Contact inhibition of what? An analytical review. J. Cell. Physiol. 1973, 81, 25–37. [Google Scholar] [CrossRef]
- Puliafito, A.; Hufnagel, L.; Neveu, P.; Streichan, S.; Sigal, A.; Fygenson, D.K.; Shraiman, B.I. Collective and single cell behavior in epithelial contact inhibition. Proc. Natl. Acad. Sci. USA 2012, 109, 739–744. [Google Scholar] [CrossRef] [Green Version]
- Santosh, A.B.; Jones, T.J. The epithelial-mesenchymal interactions: Insights into physiological and pathological aspects of oral tissues. Oncol. Rev. 2014, 8, 239. [Google Scholar] [CrossRef] [PubMed]
- Coulomb, B.; Lebreton, C.; Dubertret, L. Influence of human dermal fibroblasts on epidermalization. J. Investig. Dermatol. 1989, 92, 122–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maas-Szabowski, N.; Shimotoyodome, A.; Fusenig, N.E. Keratinocyte growth regulation in fibroblast cocultures via a double paracrine mechanism. J. Cell Sci. 1999, 112 Pt 12, 1843–1853. [Google Scholar]
- van Engeland, M.; Nieland, L.J.; Ramaekers, F.C.; Schutte, B.; Reutelingsperger, C.P. Annexin V-affinity assay: A review on an apoptosis detection system based on phosphatidylserine exposure. Cytometry 1998, 31, 1–9. [Google Scholar] [CrossRef]
- Sun, P. Contact inhibition against senescence. Oncotarget 2014, 5, 7212–7213. [Google Scholar] [CrossRef] [PubMed]
- Childs, B.G.; Durik, M.; Baker, D.J.; van Deursen, J.M. Cellular senescence in aging and age-related disease: From mechanisms to therapy. Nat. Med. 2015, 21, 1424–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holley, R.W.; Kiernan, J.A. “Contact inhibition” of cell division in 3T3 cells. Proc. Natl. Acad. Sci. USA 1968, 60, 300–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkins, M.B.; Lotze, M.T.; Dutcher, J.P.; Fisher, R.I.; Weiss, G.; Margolin, K.; Abrams, J.; Sznol, M.; Parkinson, D.; Hawkins, M.; et al. High-dose recombinant interleukin 2 therapy for patients with metastatic melanoma: Analysis of 270 patients treated between 1985 and 1993. J. Clin. Oncol. 1999, 17, 2105–2116. [Google Scholar] [CrossRef]
- Gomella, L.G.; McGinnis, D.E.; Lattime, E.C.; Butler, K.; Baltish, M.; Thompson, I.; Marshall, M.E. Treatment of transitional cell carcinoma of the bladder with intravesical interleukin-2: A pilot study. Cancer Biother. 1993, 8, 223–227. [Google Scholar] [CrossRef]
- Den Otter, W.; Dobrowolski, Z.; Bugajski, A.; Papla, B.; Van Der Meijden, A.P.; Koten, J.W.; Boon, T.A.; Siedlar, M.; Zembala, M. Intravesical interleukin-2 in T1 papillary bladder carcinoma: Regression of marker lesion in 8 of 10 patients. J. Urol. 1998, 159, 1183–1186. [Google Scholar] [CrossRef]
- Huland, E.; Heinzer, H.; Jorres, R.A.; Loppow, D.; Huland, H. Therapeutic approaches in metastatic renal cell carcinoma: Local immunotherapy. Urologe A 2004, 43 (Suppl. 3), S140–S144. [Google Scholar] [CrossRef] [PubMed]
- Huland, E.; Heinzer, H.; Huland, H.; Yung, R. Overview of interleukin-2 inhalation therapy. Cancer J. Sci. Am. 2000, 6 (Suppl. 1), S104–S112. [Google Scholar]
- Feinerman, O.; Jentsch, G.; Tkach, K.E.; Coward, J.W.; Hathorn, M.M.; Sneddon, M.W.; Emonet, T.; Smith, K.A.; Altan-Bonnet, G. Single-cell quantification of IL-2 response by effector and regulatory T cells reveals critical plasticity in immune response. Mol. Syst. Biol. 2010, 6, 437. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Lin, J.X.; Leonard, W.J. Interleukin-2 at the crossroads of effector responses, tolerance, and immunotherapy. Immunity 2013, 38, 13–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrd, V.M.; Ballard, D.W.; Miller, G.G.; Thomas, J.W. Fibroblast growth factor-1 (FGF-1) enhances IL-2 production and nuclear translocation of NF-kappaB in FGF receptor-bearing Jurkat T cells. J. Immunol. 1999, 162, 5853–5859. [Google Scholar] [PubMed]
- Cozzolino, F.; Torcia, M.; Lucibello, M.; Morbidelli, L.; Ziche, M.; Platt, J.; Fabiani, S.; Brett, J.; Stern, D. Interferon-alpha and interleukin 2 synergistically enhance basic fibroblast growth factor synthesis and induce release, promoting endothelial cell growth. J. Clin. Investig. 1993, 91, 2504–2512. [Google Scholar] [CrossRef] [Green Version]
- Krieg, C.; Letourneau, S.; Pantaleo, G.; Boyman, O. Improved IL-2 immunotherapy by selective stimulation of IL-2 receptors on lymphocytes and endothelial cells. Proc. Natl. Acad. Sci. USA 2010, 107, 11906–11911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmenate, T.; Pacios, A.; Enamorado, M.; Moreno, E.; Garcia-Martinez, K.; Fuente, D.; Leon, K. Human IL-2 mutein with higher antitumor efficacy than wild type IL-2. J. Immunol. 2013, 190, 6230–6238. [Google Scholar] [CrossRef] [Green Version]
- Levin, A.M.; Bates, D.L.; Ring, A.M.; Krieg, C.; Lin, J.T.; Su, L.; Moraga, I.; Raeber, M.E.; Bowman, G.R.; Novick, P.; et al. Exploiting a natural conformational switch to engineer an interleukin-2 ‘superkine’. Nature 2012, 484, 529–533. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.R.; Joseph, R.R.; Rosen, K.; Reginato, M.J.; Jackson, A.; Allaire, N.; Brugge, J.S.; Jobin, C.; Stadnyk, A.W. Activation of NF-kappaB following detachment delays apoptosis in intestinal epithelial cells. Oncogene 2005, 24, 6482–6491. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Garcia, A.; Merida, I.; Martinez, A.C.; Carrera, A.C. Intermediate affinity interleukin-2 receptor mediates survival via a phosphatidylinositol 3-kinase-dependent pathway. J. Biol. Chem. 1997, 272, 10220–10226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, P.J. The JAK-STAT signaling pathway: Input and output integration. J. Immunol. 2007, 178, 2623–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamat, A.M.; Colombel, M.; Sundi, D.; Lamm, D.; Boehle, A.; Brausi, M.; Buckley, R.; Persad, R.; Palou, J.; Soloway, M.; et al. BCG-unresponsive non-muscle-invasive bladder cancer: Recommendations from the IBCG. Nat. Rev. Urol. 2017, 14, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Lamm, D.L. Complications of bacillus Calmette-Guerin immunotherapy. Urol. Clin. N. Am. 1992, 19, 565–572. [Google Scholar]
- Brausi, M.; Oddens, J.; Sylvester, R.; Bono, A.; van de Beek, C.; van Andel, G.; Gontero, P.; Turkeri, L.; Marreaud, S.; Collette, S.; et al. Side effects of Bacillus Calmette-Guerin (BCG) in the treatment of intermediate- and high-risk Ta, T1 papillary carcinoma of the bladder: Results of the EORTC genito-urinary cancers group randomised phase 3 study comparing one-third dose with full dose and 1 year with 3 years of maintenance BCG. Eur. Urol. 2014, 65, 69–76. [Google Scholar] [CrossRef]
- Sylvester, R.J.; Brausi, M.A.; Kirkels, W.J.; Hoeltl, W.; Calais Da Silva, F.; Powell, P.H.; Prescott, S.; Kirkali, Z.; van de Beek, C.; Gorlia, T.; et al. Long-term efficacy results of EORTC genito-urinary group randomized phase 3 study 30911 comparing intravesical instillations of epirubicin, bacillus Calmette-Guerin, and bacillus Calmette-Guerin plus isoniazid in patients with intermediate- and high-risk stage Ta T1 urothelial carcinoma of the bladder. Eur. Urol. 2010, 57, 766–773. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Lu, J.; Huang, Y.; Ma, L. Clinical Spectrum of Complications Induced by Intravesical Immunotherapy of Bacillus Calmette-Guerin for Bladder Cancer. J. Oncol. 2019, 2019, 6230409. [Google Scholar] [CrossRef]
- Guallar-Garrido, S.; Julian, E. Bacillus Calmette-Guerin (BCG) Therapy for Bladder Cancer: An Update. Immunotargets Ther. 2020, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Larsen, E.S.; Joensen, U.N.; Poulsen, A.M.; Goletti, D.; Johansen, I.S. Bacillus Calmette-Guerin immunotherapy for bladder cancer: A review of immunological aspects, clinical effects and BCG infections. APMIS 2020, 128, 92–103. [Google Scholar] [CrossRef] [Green Version]
- Pichler, R.; Fritz, J.; Zavadil, C.; Schafer, G.; Culig, Z.; Brunner, A. Tumor-infiltrating immune cell subpopulations influence the oncologic outcome after intravesical Bacillus Calmette-Guerin therapy in bladder cancer. Oncotarget 2016, 7, 39916–39930. [Google Scholar] [CrossRef] [Green Version]
- Tosello, A.C.; Mary, F.; Amiot, M.; Bernard, A.; Mary, D. Activation of T cells via CD55: Recruitment of early components of the CD3-TCR pathway is required for IL-2 secretion. J. Inflamm. 1998, 48, 13–27. [Google Scholar] [PubMed]
- Fishman, M.N.; Thompson, J.A.; Pennock, G.K.; Gonzalez, R.; Diez, L.M.; Daud, A.I.; Weber, J.S.; Huang, B.Y.; Tang, S.; Rhode, P.R.; et al. Phase I trial of ALT-801, an interleukin-2/T-cell receptor fusion protein targeting p53 (aa264-272)/HLA-A*0201 complex, in patients with advanced malignancies. Clin. Cancer Res. 2011, 17, 7765–7775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Z.; Hanel, E.; Mak, A.; Moore, R.B. Antitumor Efficacy of Intravesical BCG, Gemcitabine, Interferon-alpha and Interleukin-2 as Mono- or Combination-Therapy for Bladder Cancer in an Orthotopic Tumor Model. Clin. Med. Insights Oncol. 2011, 5, 315–323. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmitz, L.; Berdien, B.; Huland, E.; Dase, P.; Beutel, K.; Fisch, M.; Engel, O. The Impact of a New Interleukin-2-Based Immunotherapy Candidate on Urothelial Cells to Support Use for Intravesical Drug Delivery. Life 2020, 10, 231. https://doi.org/10.3390/life10100231
Schmitz L, Berdien B, Huland E, Dase P, Beutel K, Fisch M, Engel O. The Impact of a New Interleukin-2-Based Immunotherapy Candidate on Urothelial Cells to Support Use for Intravesical Drug Delivery. Life. 2020; 10(10):231. https://doi.org/10.3390/life10100231
Chicago/Turabian StyleSchmitz, Lisa, Belinda Berdien, Edith Huland, Petra Dase, Karin Beutel, Margit Fisch, and Oliver Engel. 2020. "The Impact of a New Interleukin-2-Based Immunotherapy Candidate on Urothelial Cells to Support Use for Intravesical Drug Delivery" Life 10, no. 10: 231. https://doi.org/10.3390/life10100231