
Reversible Transitions in a Fluctuation Assay Modify the Tail of Luria–Delbrück Distribution

Abstract
:1. Introduction
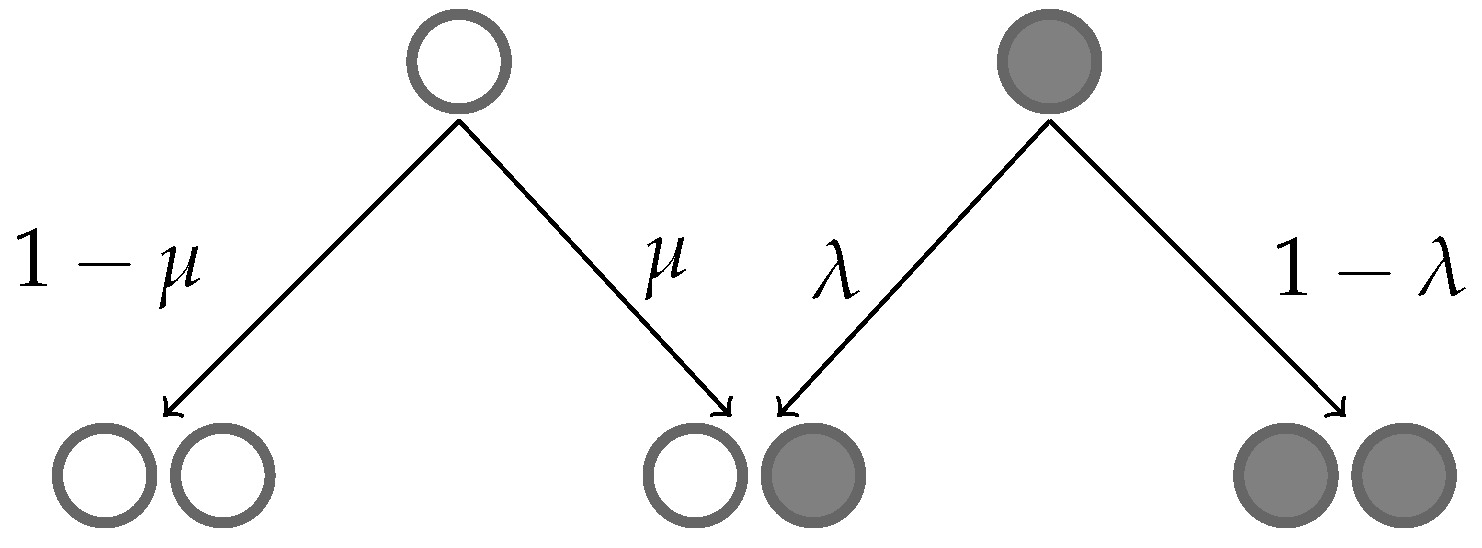
2. Model Formulation
3. Methods
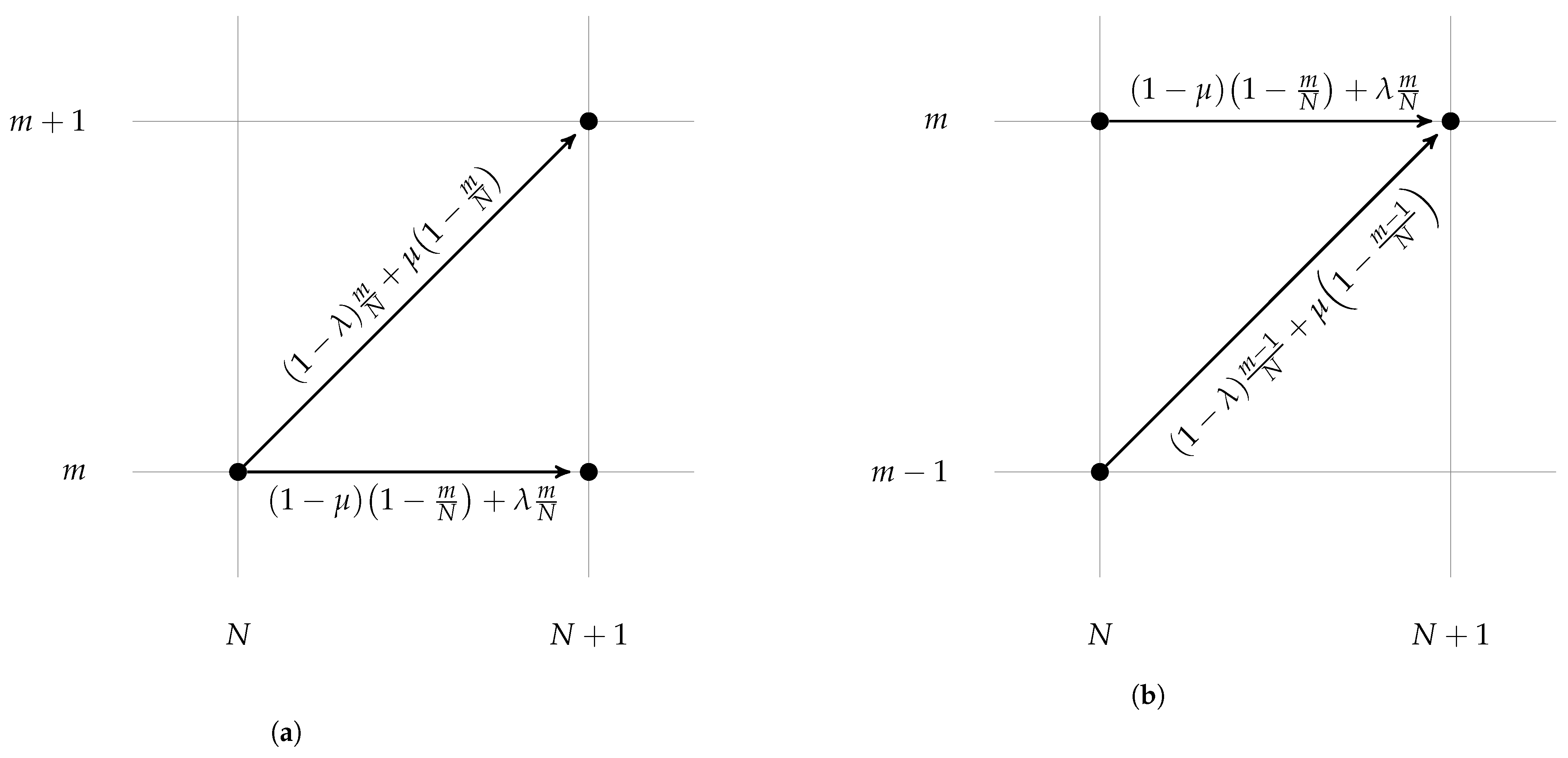
3.1. Overview of Asymptotic Approximations
3.2. Regular Solution
3.2.1. Case and
3.2.2. Case ,
3.2.3. Case
3.3. Regular Left Solution
- Case and
- Case ,
- Case
3.4. Regular Right Solution
3.4.1. Case and
3.4.2. Case ,
3.4.3. Case
3.5. Regular Coarse-Grained Solution
3.5.1. Case and
3.5.2. Case ,
3.5.3. Case
3.6. Left Boundary Layer Solution
3.6.1. Case ,
- x transforms into the shift operator.
- transforms into the summation operator (a discrete convolution with a sequence of ones).
- transforms into the kth power of this operator (a discrete convolution with the sequence ).
- transforms into the Lea–Coulson probability mass function (PMF).
3.6.2. Case ,
3.6.3. Case
3.7. Right Boundary Layer Solution
3.7.1. Case ,
3.7.2. Case ,
3.7.3. Case
3.8. Log-Composite Solution
3.8.1. Case ,
3.8.2. Case ,
3.8.3. Case
3.9. Landau Distribution
4. Results
4.1. One Sensitive Cell at the Beginning
4.2. One Sensitive and One Resistant Cell at the Beginning
4.3. More than One Sensitive or Resistant Cells at the Beginning
4.4. Stochastic Initial Conditions
4.5. Limitations of the Approximations
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PMF | Probability mass function |
| Probability density function | |
| CDF | Cumulative distribution function |
| L–C | Lea–Coulson |
Appendix A. Polya urn Models
Appendix B. Computations

References
- Luria, S.E.; Delbrück, M. Mutations of bacteria from virus sensitivity to virus resistance. Genetics 1943, 28, 491. [Google Scholar] [CrossRef] [PubMed]
- Shaffer, S.M.; Dunagin, M.C.; Torborg, S.R.; Torre, E.A.; Emert, B.; Krepler, C.; Beqiri, M.; Sproesser, K.; Brafford, P.A.; Xiao, M.; et al. Rare cell variability and drug-induced reprogramming as a mode of cancer drug resistance. Nature 2017, 546, 431–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaffer, S.M.; Emert, B.L.; Hueros, R.A.R.; Cote, C.; Harmange, G.; Schaff, D.L.; Sizemore, A.E.; Gupte, R.; Torre, E.; Singh, A.; et al. Memory sequencing reveals heritable single-cell gene expression programs associated with distinct cellular behaviors. Cell 2020, 182, 947–959. [Google Scholar] [CrossRef] [PubMed]
- Hossain, T.; Singh, A.; Butzin, N.C. Escherichia coli cells are primed for survival before lethal antibiotic stress. bioRxiv 2022. [Google Scholar] [CrossRef]
- Chang, C.A.; Jen, J.; Jiang, S.; Sayad, A.; Mer, A.S.; Brown, K.R.; Nixon, A.M.; Dhabaria, A.; Tang, K.H.; Venet, D.; et al. Ontogeny and vulnerabilities of drug-tolerant persisters in her2+ breast cancer. Cancer Discov. 2022, 12, 1022–1045. [Google Scholar] [CrossRef]
- Harmange, G.; Hueros, R.A.R.; Schaff, D.L.; Emert, B.L.; Saint-Antoine, M.M.; Nellore, S.; Fane, M.E.; Alicea, G.M.; Weeraratna, A.T.; Singh, A.; et al. Disrupting cellular memory to overcome drug resistance. bioRxiv 2022. [Google Scholar] [CrossRef]
- Saint-Antoine, M.M.; Grima, R.; Singh, A. A fluctuation-based approach to infer kinetics and topology of cell-state switching. bioRxiv 2022. [Google Scholar] [CrossRef]
- Saint-Antoine, M.M.; Singh, A. Moment-based estimation of state-switching rates in cell populations. bioRxiv 2022. [Google Scholar] [CrossRef]
- Zheng, Q. Progress of a half century in the study of the Luria–Delbrück distribution. Math. Biosci. 1999, 162, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Kessler, D.A.; Levine, H. Large population solution of the stochastic Luria–Delbrück evolution model. Proc. Natl. Acad. Sci. USA 2013, 110, 11682–11687. [Google Scholar] [CrossRef] [Green Version]
- Keller, P.; Antal, T. Mutant number distribution in an exponentially growing population. J. Stat. Mech. Theory Exp. 2015, 2015, P01011. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, M.D.; Antal, T. Universal asymptotic clone size distribution for general population growth. Bull. Math. Biol. 2016, 78, 2243–2276. [Google Scholar] [CrossRef] [Green Version]
- Pakes, A.G. Mutant number laws and infinite divisibility. Axioms 2022, 11, 584. [Google Scholar] [CrossRef]
- Antal, T.; Krapivsky, P. Exact solution of a two-type branching process: Models of tumor progression. J. Stat. Mech. Theory Exp. 2011, 2011, P08018. [Google Scholar] [CrossRef]
- Angerer, W.P. An explicit representation of the Luria–Delbrück distribution. J. Math. Biol. 2001, 42, 145–174. [Google Scholar] [CrossRef] [PubMed]
- Kessler, D.A.; Levine, H. Scaling solution in the large population limit of the general asymmetric stochastic Luria–Delbrück evolution process. J. Stat. Phys. 2015, 158, 783–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armitage, P. The statistical theory of bacterial populations subject to mutation. J. R. Stat. Soc. Ser. B (Methodol.) 1952, 14, 1–33. [Google Scholar] [CrossRef]
- Jolly, C.; Cook, A.; Raferty, J.; Jones, M. Measuring bidirectional mutation. J. Theor. Biol. 2007, 246, 269–277. [Google Scholar] [CrossRef]
- Sorace, R.; Komarova, N.L. Accumulation of neutral mutations in growing cell colonies with competition. J. Theor. Biol. 2012, 314, 84–94. [Google Scholar] [CrossRef] [Green Version]
- Cheek, D.; Antal, T. Genetic composition of an exponentially growing cell population. Stoch. Process. Their Appl. 2020, 130, 6580–6624. [Google Scholar] [CrossRef]
- Lea, D.E.; Coulson, C.A. The distribution of the numbers of mutants in bacterial populations. J. Genet. 1949, 49, 264–285. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q. A fresh approach to a special type of the Luria–Delbrück distribution. Axioms 2022, 11, 730. [Google Scholar] [CrossRef]
- Pakes, A.G. Remarks on the Luria–Delbrück distribution. J. Appl. Probab. 1993, 30, 991–994. [Google Scholar] [CrossRef] [Green Version]
- Angerer, W.P. A note on the evaluation of fluctuation experiments. Mutat. Res. Mol. Mech. Mutagen. 2001, 479, 207–224. [Google Scholar] [CrossRef] [PubMed]
- Bulyak, E.; Shul’ga, N. Landau distribution of ionization losses: History, importance, extensions. arXiv 2022, arXiv:2209.06387. [Google Scholar]
- Nolan, J.P. Univariate Stable Distributions; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Rao, C.R.; Shanbhag, D.N.; Sapatinas, T.; Rao, M.B. Some properties of extreme stable laws and related infinitely divisible random variables. J. Stat. Plan. Inference 2009, 139, 802–813. [Google Scholar]
- Bender, C.M.; Orszag, S.A. Advanced Mathematical Methods for Scientists and Engineers I: Asymptotic Methods and Perturbation Theory, 1st ed.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1999. [Google Scholar]
- Hinch, E.J. Perturbation Methods; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Hinch, R.; Chapman, S.J. Exponentially slow transitions on a Markov chain: The frequency of calcium sparks. Eur. J. Appl. Math. 2005, 16, 427–446. [Google Scholar] [CrossRef] [Green Version]
- Bressloff, P.C. Stochastic Processes in Cell Biology; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Bokes, P. Heavy-tailed distributions in a stochastic gene autoregulation model. J. Stat. Mech. Theory Exp. 2021, 2021, 113403. [Google Scholar] [CrossRef]
- Athreya, K.B.; Ney, P.E.; Ney, P. Branching Processes; Courier Corporation: Chelmsford, CA, USA, 2004. [Google Scholar]
- Kepler, T.B.; Oprea, M. Improved inference of mutation rates: I. An integral representation for the Luria–Delbrück distribution. Theor. Popul. Biol. 2001, 59, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Kevorkian, J.; Cole, J.D. Perturbation Methods in Applied Mathematics; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Murray, J.D. Mathematical Biology: I. An Introduction; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Johnson, N.L.; Kotz, S.; Kemp, A.W. Univariate Discrete Distributions; John Wiley & Sons: Hoboken, NJ, USA, 2005. [Google Scholar]
- Mahmoud, H. Pólya urn Models; Chapman and Hall/CRC: Boca Raton, FL, USA, 2008. [Google Scholar]
- Walczak, A.M.; Mugler, A.; Wiggins, C.H. Analytic methods for modeling stochastic regulatory networks. Comput. Model. Signal. Netw. 2012, 880, 273–322. [Google Scholar]
- Bokes, P.; King, J.R.; Loose, M. A bistable genetic switch which does not require high co-operativity at the promoter: A two-timescale model for the PU. 1–GATA-1 interaction. Math. Med. Biol. A J. IMA 2009, 26, 117–132. [Google Scholar] [CrossRef] [PubMed]
- Landau, L.D. On the energy loss of fast particles by ionization. J. Phys. 1944, 8, 201–205. [Google Scholar]
- Frank, S.A. Numbers of mutations within multicellular bodies: Why it matters. Axioms 2022, 12, 12. [Google Scholar] [CrossRef]
- Su, Y.; Wei, W.; Robert, L.; Xue, M.; Tsoi, J.; Garcia-Diaz, A.; Homet Moreno, B.; Kim, J.; Ng, R.H.; Lee, J.W.; et al. Single-cell analysis resolves the cell state transition and signaling dynamics associated with melanoma drug-induced resistance. Proc. Natl. Acad. Sci. USA 2017, 114, 13679–13684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelini, E.; Wang, Y.; Zhou, J.X.; Qian, H.; Huang, S. A model for the intrinsic limit of cancer therapy: Duality of treatment-induced cell death and treatment-induced stemness. PLoS Comput. Biol. 2022, 18, e1010319. [Google Scholar] [CrossRef] [PubMed]
- Symbolic Math Toolbox. Available online: https://www.mathworks.com/products/symbolic.html (accessed on 16 January 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| regular (4), (10) | |||
| regular left (13), (14) | regular coarse-grained (20), (21) | regular right (16), (17) | |
| left (32), (36) | right (40) (41) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bokes, P.; Hlubinová, A.; Singh, A. Reversible Transitions in a Fluctuation Assay Modify the Tail of Luria–Delbrück Distribution. Axioms 2023, 12, 249. https://doi.org/10.3390/axioms12030249
Bokes P, Hlubinová A, Singh A. Reversible Transitions in a Fluctuation Assay Modify the Tail of Luria–Delbrück Distribution. Axioms. 2023; 12(3):249. https://doi.org/10.3390/axioms12030249
Chicago/Turabian StyleBokes, Pavol, Anna Hlubinová, and Abhyudai Singh. 2023. "Reversible Transitions in a Fluctuation Assay Modify the Tail of Luria–Delbrück Distribution" Axioms 12, no. 3: 249. https://doi.org/10.3390/axioms12030249