
Dynamic Behavior of a Predator–Prey Model with Double Delays and Beddington–DeAngelis Functional Response

Abstract
:1. Introduction
2. Positivity and Boundedness
2.1. Positivity
2.2. Boundedness
3. Stability Analysis
3.1. Equilibrium Points and Existence Criterion
- (1)
- the trivial equilibrium point ;
- (2)
- the free equilibrium point ;
- (3)
- the coexistence equilibrium point satisfying the following equation:
3.2. Local Stability Analysis and Hopf Bifurcation of Equilibria
- (1)
- At :
- (2)
- At :and the characteristic equation is
- (3)
- At :where .
4. Stochastic Delay Model Analysis
4.1. Existence and Uniqueness of Positive Solution
4.2. Stochastic Ultimate Boundedness
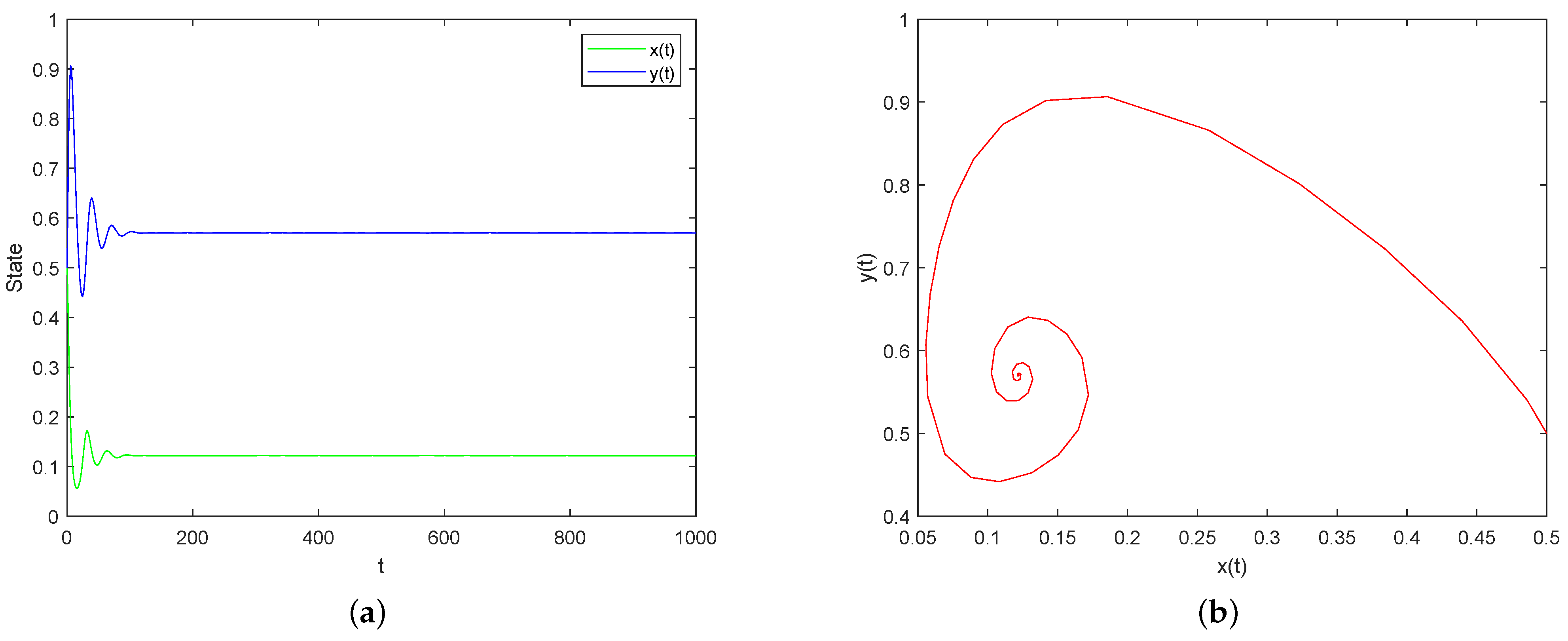
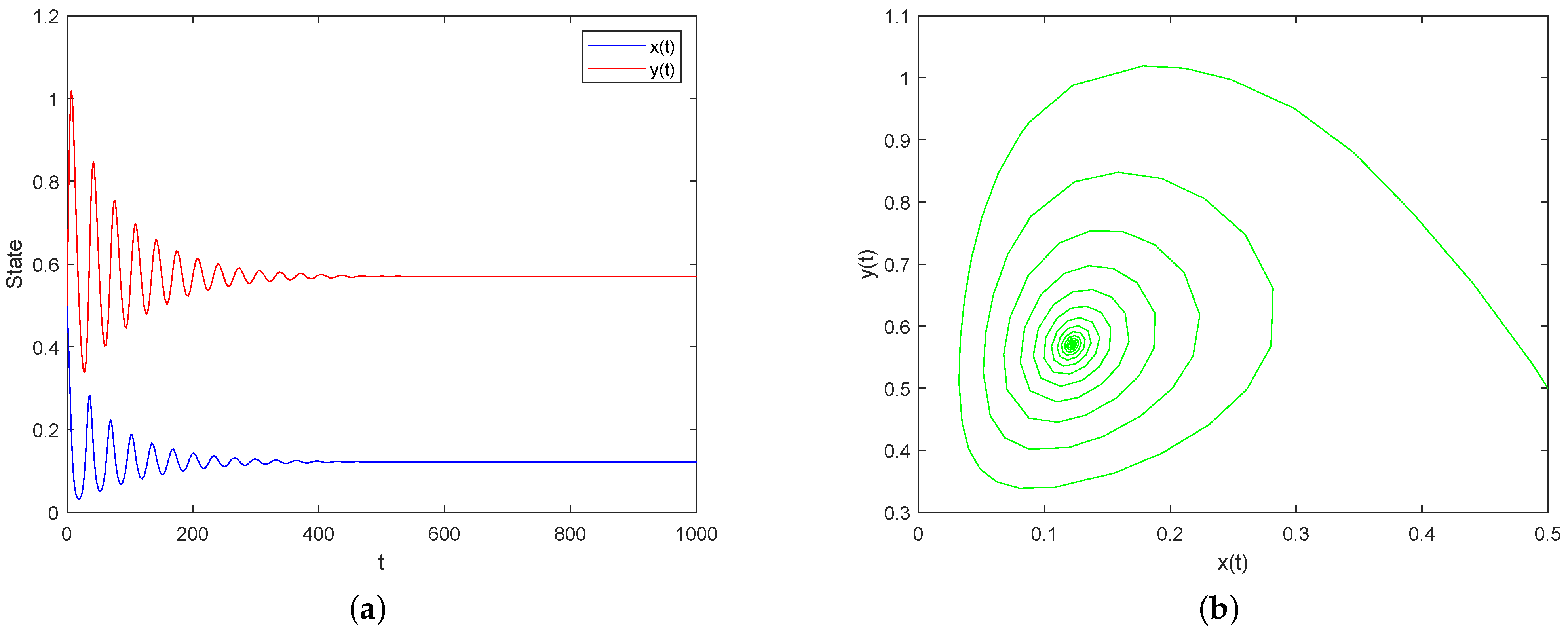
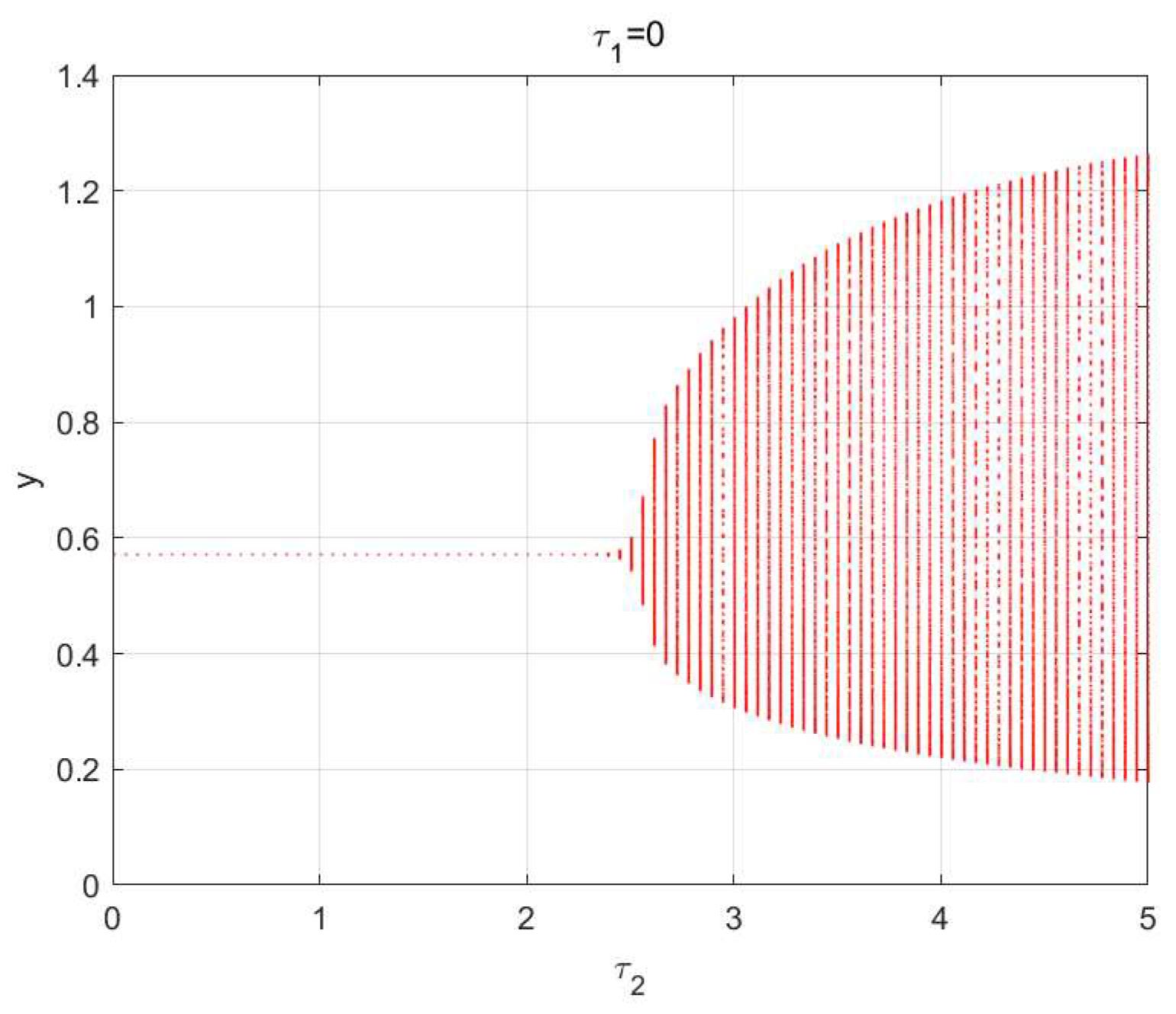
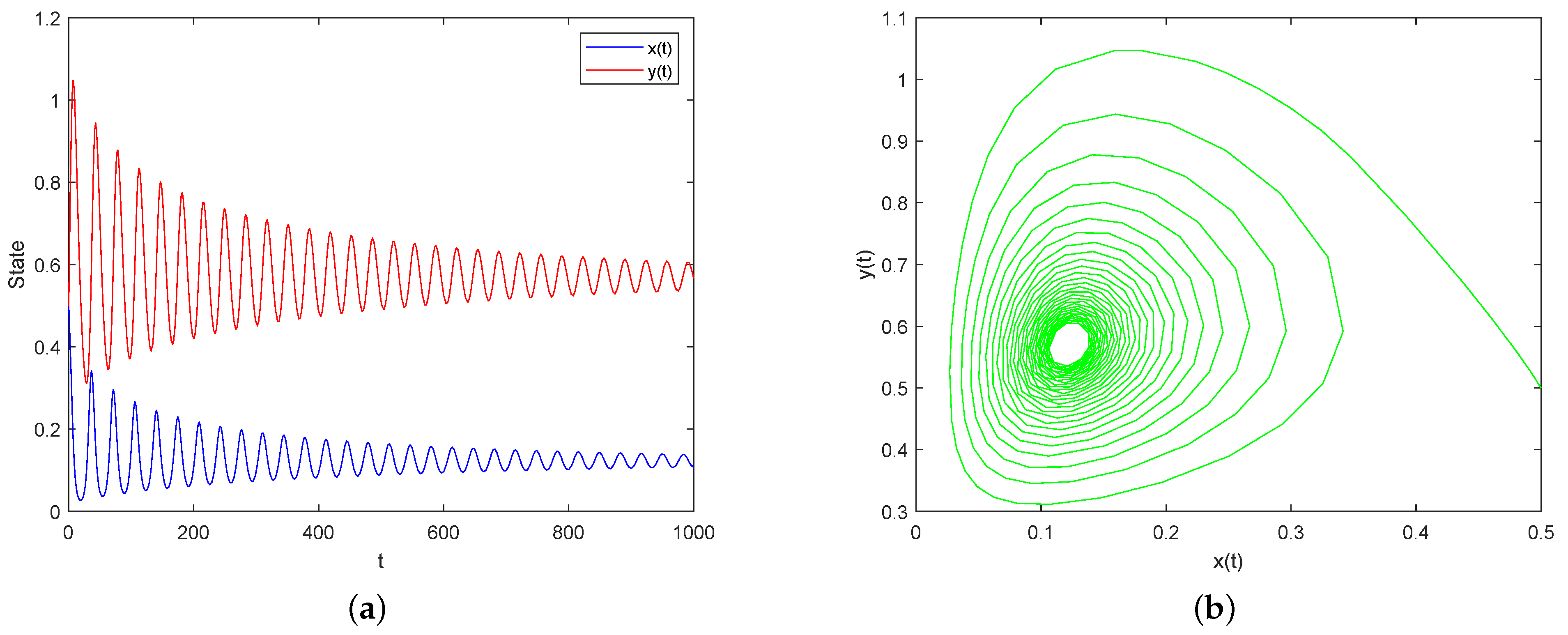
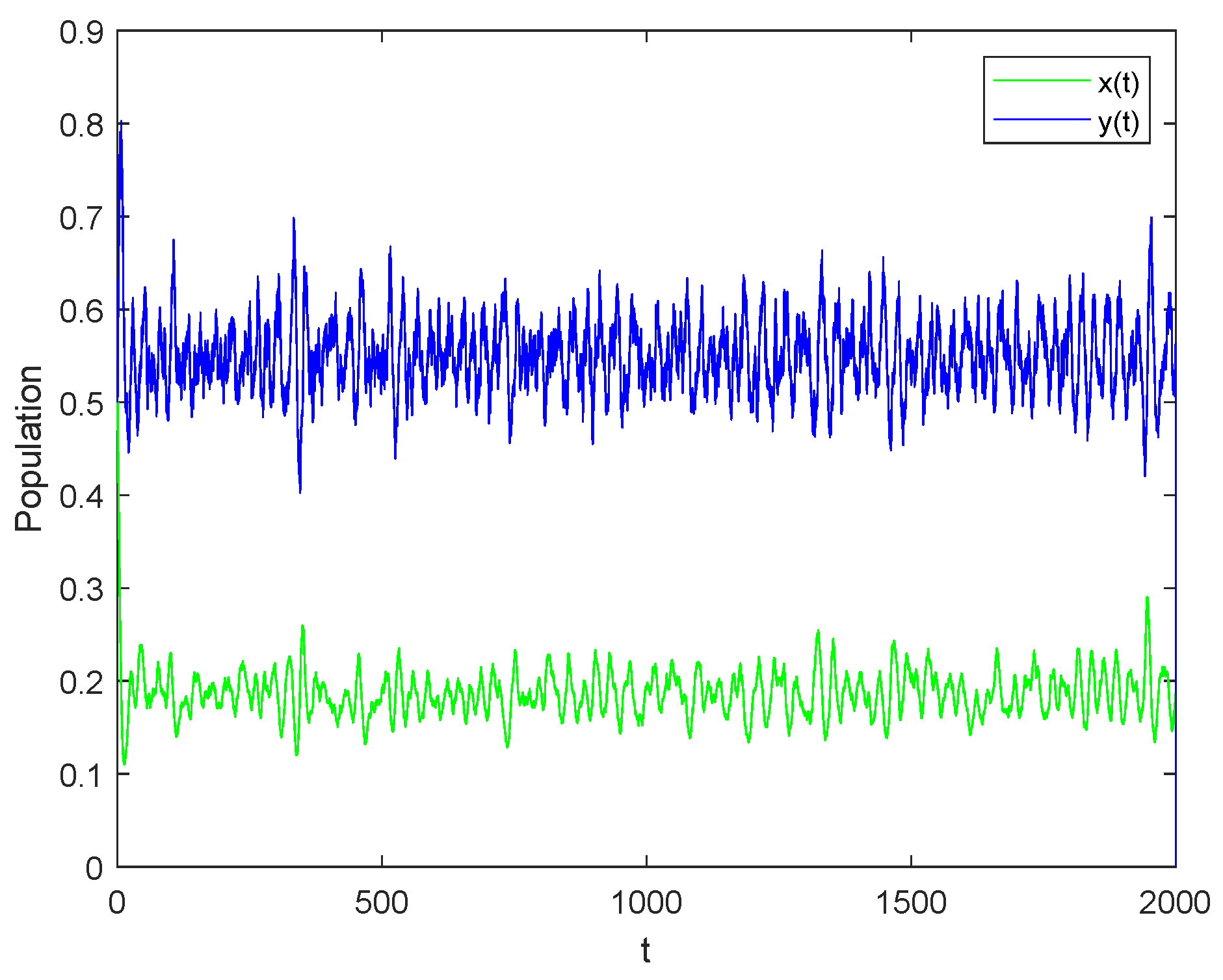
5. Numerical Simulations
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sahoo, D.; Samanta, G.P. Impact of fear effect in a two prey-one predator system with switching behaviour in predation. Differ. Equ. Dyn. Syst. 2021, 1–23. [Google Scholar] [CrossRef]
- Mondal, B.; Sarkar, S.; Ghosh, U. Complex dynamics of a generalist predator–prey model with hunting cooperation in predator. Eur. Phys. J. Plus 2022, 137, 43. [Google Scholar] [CrossRef]
- Pal, S.; Majhi, S.; Mandal, S.; Pal, N. Role of fear in a predator–prey model with Beddington–DeAngelis functional response. Z. FüR Nat. A 2019, 74, 581–595. [Google Scholar] [CrossRef]
- Huang, G.; Ma, W.; Takeuchi, Y. Global analysis for delay virus dynamics model with Beddington–DeAngelis functional response. Appl. Math. Lett. 2011, 24, 1199–1203. [Google Scholar] [CrossRef] [Green Version]
- Alam, S. Risk of disease-selective predation in an infected prey-predator system. J. Biol. Syst. 2009, 17, 111–124. [Google Scholar] [CrossRef]
- Roy, J.; Alam, S. Dynamics of an autonomous food chain model and existence of global attractor of the associated non-autonomous system. Int. J. Biomath. 2019, 12, 1950082. [Google Scholar] [CrossRef]
- Zanette, L.Y.; White, A.F.; Allen, M.C.; Clinchy, M. Perceived predation risk reduces the number of offspring songbirds produce per year. Science 2011, 334, 1398–1401. [Google Scholar] [CrossRef]
- Creel, S.; Christianson, D.; Liley, S.; Winnie, J.A., Jr. Predation risk affects reproductive physiology and demography of elk. Science 2007, 315, 960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eggers, S.; Griesser, M.; Nystr, M.; Ekman, J. Predation risk induces changes in nest-site selection and clutch size in the Siberian jay. PRoceedings R. Soc. B Biol. Sci. 2006, 273, 701–706. [Google Scholar] [CrossRef] [Green Version]
- Orrock, J.L.; Fletcher, R.J., Jr. An island-wide predator manipulation reveals immediate and long-lasting matching of risk by prey. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140391. [Google Scholar] [CrossRef]
- Sheriff, M.J.; Krebs, C.J.; Boonstra, R. The sensitive hare: Sublethal effects of predator stress on reproduction in snowshoe hares. J. Anim. Ecol. 2009, 78, 1249–1258. [Google Scholar] [CrossRef] [PubMed]
- Creel, S.; Christianson, D. Relationships between direct predation and risk effects. Trends Ecol. Evol. 2008, 23, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Lima, S.L. Predators and the breeding bird: Behavioral and reproductive flexibility under the risk of predation. Biol. Rev. 2009, 84, 485–513. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Maiti, A.; Samanta, G.P. Effects of fear and additional food in a delayed predator–prey model. Biophys. Rev. Lett. 2018, 13, 157–177. [Google Scholar] [CrossRef]
- Shao, Y. Global stability of a delayed predator–prey system with fear and Holling-type II functional response in deterministic and stochastic environments. Math. Comput. Simul. 2022, 200, 65–77. [Google Scholar] [CrossRef]
- Xia, Y.; Yuan, S. Survival analysis of a stochastic predator–prey model with prey refuge and fear effect. J. Biol. Dyn. 2020, 14, 871–892. [Google Scholar] [CrossRef]
- Liu, S.; Beretta, E.; Breda, D. Predator–prey model of Beddington–DeAngelis type with maturation and gestation delays. Nonlinear Anal. Real World Appl. 2010, 11, 4072–4091. [Google Scholar] [CrossRef]
- Bhattacharyyai, A.; Pal, A.K. Complex dynamics of delay induced two-prey one-predator model with Beddington-DeAngelis response function. Nonlinear Stud. 2021, 28. [Google Scholar]
- Li, J.; Liu, X.; Wei, C. The impact of fear factor and self-defence on the dynamics of predator-prey model with digestion delay. Math. Biosci. Eng. 2021, 18, 5478–5504. [Google Scholar] [CrossRef]
- Djilali, S.; Cattani, C.; Guin, L.N. Delayed predator–prey model with prey social behavior. Eur. Phys. J. Plus 2021, 136, 940. [Google Scholar] [CrossRef]
- Shu, H.; Hu, X.; Wang, L.; Watmough, J. Delay induced stability switch, multitype bistability and chaos in an intraguild predation model. J. Math. Biol. 2015, 71, 1269–1298. [Google Scholar] [CrossRef]
- Yuan, J.; Zhao, L.; Huang, C.; Xiao, M. Stability and bifurcation analysis of a fractional predator–prey model involving two nonidentical delays. Math. Comput. Simul. 2021, 181, 562–580. [Google Scholar] [CrossRef]
- Kong, W.; Shao, Y. The long time behavior of equilibrium status of a predator-prey system with delayed fear in deterministic and stochastic scenarios. J. Math. 2022, 2022, 3214358. [Google Scholar] [CrossRef]
- Pay, P.; Samanta, S.; Pal, N.; Chattopadhyay, J. Delay induced multiple stability switch and chaos in a predator–prey model with fear effect. Math. Comput. Simul. 2020, 172, 134–158. [Google Scholar]
- Kumar, A.; Dubey, B. Modeling the effect of fear in a prey–predator system with prey refuge and gestation delay. Int. J. Bifurc. Chaos 2019, 29, 1950195. [Google Scholar] [CrossRef]
- Roy, J.; Alam, S. Fear factor in a prey–predator system in deterministic and stochastic environment. Phys. A Stat. Mech. Its Appl. 2020, 541, 123359. [Google Scholar] [CrossRef]
- Yang, X.; Chen, L.; Chen, J. Permanence and positive periodic solution for the single-species nonautonomous delay diffusive models. Comput. Math. Appl. 1996, 32, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Freedman, H.L.; Sree Hari Rao, V. The trade-off between mutual interference and time lags in predator-prey systems. Bull. Math. Biol. 1983, 45, 991–1004. [Google Scholar] [CrossRef]
- Peng, S.; Zhu, X. Necessary and sufficient condition for comparison theorem of 1-dimensional stochastic differential equations. Stoch. Process. Their Appl. 2006, 116, 370–380. [Google Scholar] [CrossRef] [Green Version]
- Higham, D.J. An algorithmic introduction to numerical simulation of stochastic differential equations. SIAM Rev. 2001, 43, 525–546. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, M.; Shao, Y.; Xue, R.; Zhao, J. Dynamic Behavior of a Predator–Prey Model with Double Delays and Beddington–DeAngelis Functional Response. Axioms 2023, 12, 73. https://doi.org/10.3390/axioms12010073
Cui M, Shao Y, Xue R, Zhao J. Dynamic Behavior of a Predator–Prey Model with Double Delays and Beddington–DeAngelis Functional Response. Axioms. 2023; 12(1):73. https://doi.org/10.3390/axioms12010073
Chicago/Turabian StyleCui, Minjuan, Yuanfu Shao, Renxiu Xue, and Jinxing Zhao. 2023. "Dynamic Behavior of a Predator–Prey Model with Double Delays and Beddington–DeAngelis Functional Response" Axioms 12, no. 1: 73. https://doi.org/10.3390/axioms12010073