Mycoextraction: Rapid Cadmium Removal by Macrofungi-Based Technology from Alkaline Soil


Abstract
:1. Introduction
2. Methods and Materials
2.1. Strains and Soil
2.2. Cd Aging in the Soil
2.3. Removable Mushroom Bagging Design and Conditions for Fruiting Body Development
2.4. Pot Experiment of Cd Mycoextraction
2.5. Chemical Analysis
2.6. Transcriptomic Sequencing
2.7. Reference Mapping and RNA-Seq Analysis
2.8. Differentially Gene Expression Analysis
2.9. Functional Annotation of Candidate Genes
2.10. Statistical Analysis
3. Results
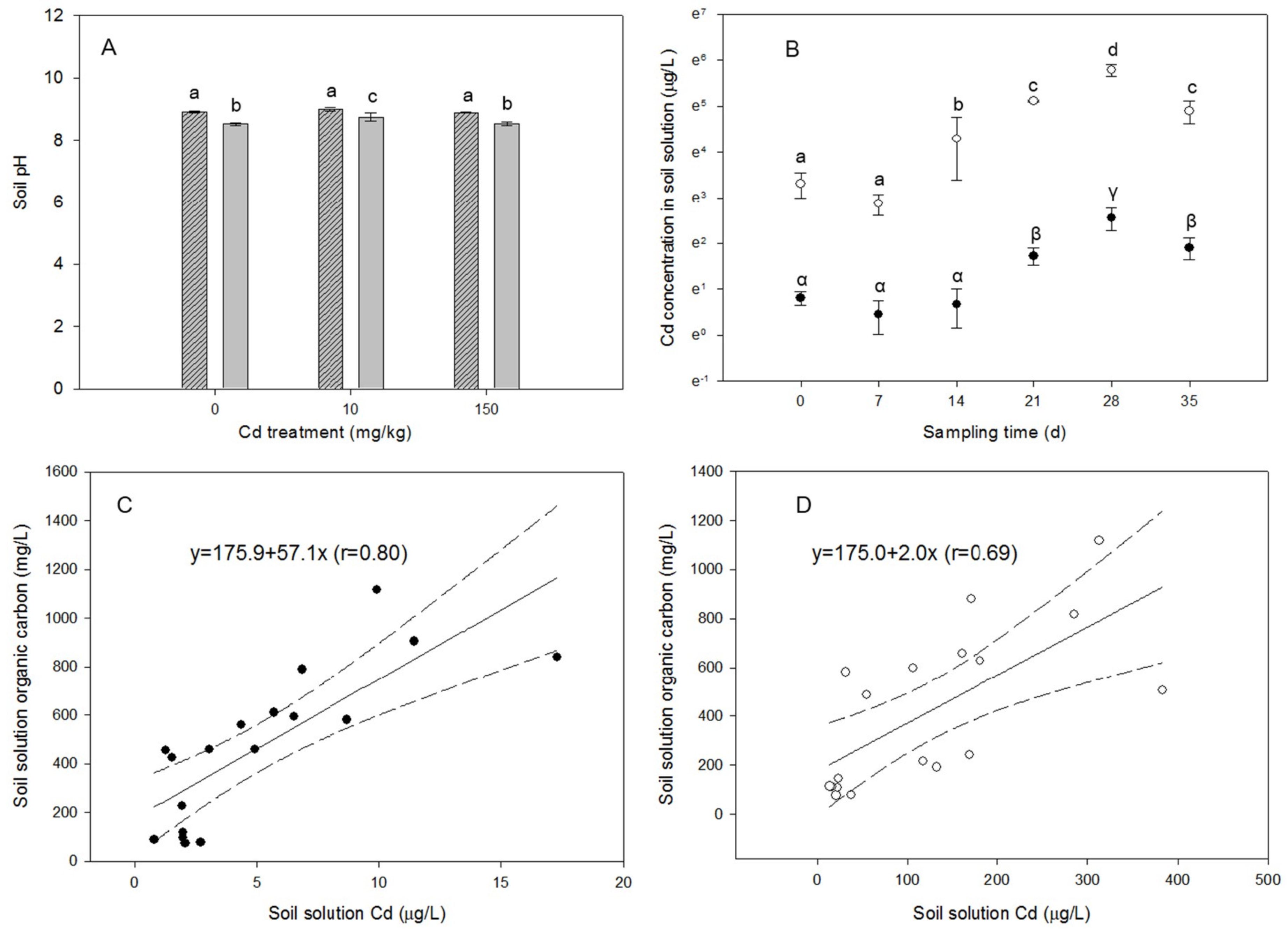
3.1. The Soil
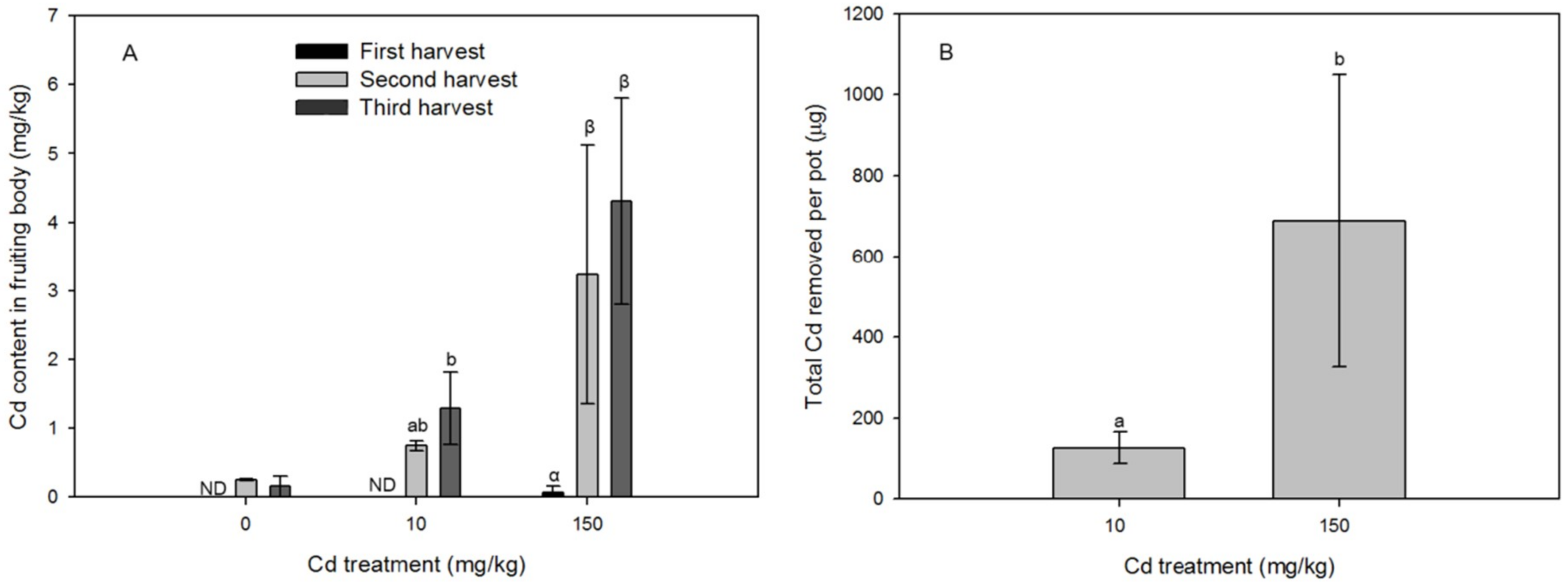
3.2. Dynamics in Mushroom Biomass and Biomass Cd
3.3. Dynamics in Soil Chemistry
3.4. Data Quality of the Transcriptomic Sequencing
3.5. Mapping Statistics and DEG Identification
3.6. Function Enrichment Analysis
4. Discussion
4.1. Ability of the Macrofungi System in Cd Sorption in the Alkaline Environment
4.2. Cd Removal Efficiency of the Macrofungi System
4.3. Transcriptomic Response of the Oyster Mushroom to Cd
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cimino, G.; Caristi, C. Acute toxicity of heavy metals to aerobic digestion of waste cheese whey. Biol. Waste 1990, 33, 201–210. [Google Scholar] [CrossRef]
- Pérez-Marín, A.B.; Zapata, V.M.; Ortuño, J.F.; Aguilar, M.; Sáez, J.; Lloréns, M. Removal of cadmium from aqueous solutions by adsorption onto orange waste. J. Hazard. Mater. 2007, 139, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Anahid, S.; Yaghmaei, S.; Ghobadinejad, Z. Heavy metal tolerance of fungi. Sci. Iran. 2011, 18, 502–508. [Google Scholar] [CrossRef]
- Mohammadian, E.; Babai Ahari, A.; Arzanlou, M.; Oustan, S.; Khazaei, S.H. Tolerance to heavy metals in filamentous fungi isolated from contaminated mining soils in the Zanjan Province, Iran. Chemosphere 2017, 185, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Chuang, H.W.; Wang, I.W.; Lin, S.Y.; Chang, Y.L. Transcriptome analysis of cadmium response in Ganoderma lucidum. FEMS Microbiol. Lett. 2009, 293, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Damodaran, D.; Shetty, K.V.; Mohan, B.R. Uptake of certain heavy metals from contaminated soil by mushroom—Galerina vittiformis. Ecotoxicol. Environ. Saf. 2014, 104, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, S.; Mahmoodulhassan, M.; Ahmad, R.; Suthor, V.; Yasin, M. Metal tolerance potential of filamentous fungi isolated from soils irrigated with untreated municipal effluent. Soil Environ. 2013, 32, 55–62. [Google Scholar]
- Akar, T.; Cabuk, A.; Tunali, S.; Yamac, M. Biosorption potential of the macrofungus Ganoderma carnosum for removal of lead(II) ions from aqueous solutions. Environ. Lett. 2006, 41, 2587–2606. [Google Scholar] [CrossRef] [PubMed]
- Xiangliang, P.; Jianlong, W.; Daoyong, Z. Biosorption of Pb(II) by Pleurotus ostreatus immobilized in calcium alginate gel. Process Biochem. 2005, 40, 2799–2803. [Google Scholar] [CrossRef]
- Tabrizi, L.; Mohammadi, S.; Delshad, M.; Moteshare, Z.B. Effect of Arbuscular Mycorrhizal Fungi on Yield and Phytoremediation Performance of Pot Marigold (Calendula officinalis L.) Under Heavy Metals Stress. Int. J. Phytoremed. 2015, 17, 1244–1252. [Google Scholar] [CrossRef] [PubMed]
- Chang, Q.; Diao, F.-W.; Wang, Q.-F.; Pan, L.; Dang, Z.-H.; Guo, W. Effects of arbuscular mycorrhizal symbiosis on growth, nutrient and metal uptake by maize seedlings (Zea mays L.) grown in soils spiked with Lanthanum and Cadmium. Environ. Pollut. 2018, 241, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Shahabivand, S.; Maivan, H.Z.; Goltapeh, E.M.; Sharifi, M.; Aliloo, A.A. The effects of root endophyte and arbuscular mycorrhizal fungi on growth and cadmium accumulation in wheat under cadmium toxicity. Plant Physiol. Biochem. 2012, 60, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Guo, S.; Jiao, K.; Hou, J.; Xie, H.; Xu, H. Bioremediation of soils co-contaminated with heavy metals and 2,4,5-trichlorophenol by fruiting body of Clitocybe maxima. J. Hazard. Mater. 2015, 294, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Liu, H.; Li, Q.; Gao, N.; Yao, Y.; Xu, H. Combined remediation of Cd–phenanthrene co-contaminated soil by Pleurotus cornucopiae and Bacillus thuringiensis FQ1 and the antioxidant responses in Pleurotus cornucopiae. Ecotoxicol. Environ. Saf. 2015, 120, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zheng, X.; Chen, L.; Li, X. Cadmium Resistance Screening of Oyster Mushrooms from North China for Mycoremediation. Pedosphere 2018, 28. in press. [Google Scholar] [CrossRef]
- Li, X.F.; Huang, Y.Z.; Ma, Y.B.; Sun, J.W.; Cui, H.J. Leaching impacts Ni toxicity differently among soils but increases its predictability according to nitrification assay. J. Soils Sediments 2010, 10, 579–589. [Google Scholar] [CrossRef]
- Matejovic, I. Determination of carbon and nitrogen in samples of various soils by the dry combustion. Commun. Soil Sci. Plant Anal. 1997, 28, 1499–1511. [Google Scholar] [CrossRef]
- Roje, V. A fast method for multi-metal determination in soil samples by high-resolution inductively-coupled plasma-mass spectrometry (HR-ICP-MS). Chem. Speciat. Bioavailab. 2010, 22, 135–139. [Google Scholar] [CrossRef]
- Riley, R.; Salamov, A.A.; Brown, D.W.; Nagy, L.G.; Floudas, D.; Held, B.W.; Levasseur, A.; Lombard, V.; Morin, E.; Otillar, R.; et al. Extensive sampling of basidiomycete genomes demonstrates inadequacy of the white-rot/brown-rot paradigm for wood decay fungi. Proc. Natl. Acad. Sci. USA 2014, 111, 9923–9928. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N.; Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323–323. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol 2014, 15. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: tool for the unification of biology. Nat Genet 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koonin, E.V.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Krylov, D.M.; Makarova, K.S.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; Rao, B.S.; et al. A comprehensive evolutionary classification of proteins encoded in complete eukaryotic genomes. Genome Biol 2004, 5. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Ge, M.; Hu, H.X.; Li, G. Effects of EDTA and EDDS on heavy metal activation and accumulation of metals by soybean in alkaline soil. Soil Sediment Contam. 2015, 24, 353–367. [Google Scholar]
- Ma, Y.B.; Lombi, E.; Oliver, I.W.; Nolan, A.L.; McLaughlin, M.J. Long-term aging of copper added to soils. Environ. Sci. Technol. 2006, 40, 6310–6317. [Google Scholar] [CrossRef] [PubMed]
- van Gestel, C.A.M.; Koolhaas, J.E. Water-extractability, free ion activity, and pH explain cadmium sorption and toxicity to Folsomia candida (Collembola) in seven soil-pH combinations. Environ. Toxicol. Chem. 2004, 23, 1822–1833. [Google Scholar] [CrossRef]
- Yang, Z.; Chen, Y.; Qian, R.; Guo, C.; Zhu, H. A study of the effect of soil pH on chemical species of cadmium by simulated experiments. Earth Sci. Front. 2005, 12, 252–260. [Google Scholar]
- Li, X.F.; Sun, J.W.; Huang, Y.Z.; Ma, Y.B.; Zhu, Y.G. Copper toxicitythresholds in Chinese soils based on substrate-induced nitrification assay. Environ. Toxicol. Chem. 2010, 29, 294–300. [Google Scholar] [CrossRef]
- Yuan, S.H.; Xi, Z.M.; Jiang, Y.; Wan, J.Z.; Wu, C.; Zheng, Z.H.; Lu, X.H. Desorption of copper and cadmium from soils enhanced by organic acids. Chemosphere 2007, 68, 1289–1297. [Google Scholar] [CrossRef]
- Christensen, J.B.; Jensen, D.L.; Christensen, T.H. Effect of dissolved organic carbon on the mobility of cadmium, nickel and zinc in leachate polluted groundwater. Water Res. 1996, 30, 3037–3049. [Google Scholar] [CrossRef]
- Hernandez-Soriano, M.C.; Jimenez-Lopez, J.C. Effects of soil water content and organic matter addition on the speciation and bioavailability of heavy metals. Sci. Total Environ. 2012, 423, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.T.; Wang, Y.; Yeh, K.C. Role of root exudates in metal acquisition and tolerance. Curr. Opin. Plant Biol. 2017, 39, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Li, D.; Ye, X.; Xu, H.; Yao, G.; Wang, J.; Zhang, Q.; Hu, J.; Gao, N. Enhancement of Cd phytoextraction by hyperaccumulator Sedum alfredii using electrical field and organic amendments. Environ. Sci. Pollut. Res. Int. 2017, 24, 5060–5067. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Zhou, Z.; Liu, Y.; Jiang, J.; Li, Q.; Song, H.; Pei, D.; Xu, H. Mycoremediation potential and tolerance responses of Oudemansiella radicata in cadmium-pyrene co-contaminated soil. J. Soils Sediments 2015, 15, 1083–1093. [Google Scholar] [CrossRef]
- Lin, L.; Chen, F.; Wang, J.; Liao, M.; Lv, X.; Wang, Z.; Li, H.; Deng, Q.; Xia, H.; Liang, D. Effects of living hyperaccumulator plants and their straws on the growth and cadmium accumulation of Cyphomandra betacea seedlings. Ecotoxicol. Environ. Saf. 2018, 155, 109–116. [Google Scholar] [CrossRef]
- Lu, M.; Zhang, Z.Z.; Wang, J.X.; Zhang, M.; Xu, Y.X.; Wu, X.J. Interaction of heavy metals and pyrene on their fates in soil and tall fescue (Festuca arundinacea). Environ. Sci. Technol. 2014, 48, 1158–1165. [Google Scholar] [CrossRef] [PubMed]
- Li, N.Y.; Fu, Q.L.; Zhuang, P.; Guo, B.; Zou, B.; Li, Z.A. Effect of fertilizers on Cd uptake of Amaranthus hypochondriacus, a high biomass, fast growing and easily cultivated potential Cd hyperaccumulator. Int. J. Phytoremed. 2012, 14, 162–173. [Google Scholar] [CrossRef]
- Cojocaru, P.; Gusiatin, Z.M.; Cretescu, I. Phytoextraction of Cd and Zn as single or mixed pollutants from soil by rape (Brassica napus). Environ. Sci. Pollut. Res. Int. 2016, 23, 10693–10701. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, W.; Jiang, R.F.; Li, H.F. Comparison of phytoextraction efficiency of Cd with the hyperaccumulator Thlaspi caerulescens and three high biomass species. Acta Ecol. Sin. 2009, 29, 2732–2737. (In Chinese) [Google Scholar]
- Ji, L.Y.; Zhang, W.W.; Yu, D.; Cao, Y.R.; Xu, H. Effect of heavy metal-solubilizing microorganisms on zinc and cadmium extractions from heavy metal contaminated soil with Tricholoma lobynsis. World J. Microbiol. Biotechnol. 2012, 28, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.B.; He, N.; Zhang, Y.; Cao, Y.R.; Xu, H. Isolation and characterization of heavy-metal-mobilizing bacteria from contaminated soils and their potential in promoting Pb, Cu, and Cd accumulation by Coprinus comatus. Can. J. Microbiol. 2012, 58, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Demirbas, A. Metal ion uptake by mushrooms from natural and artificially enriched soils. Food Chem. 2002, 78, 89–93. [Google Scholar] [CrossRef]
- Lasat, M.M. Phytoextraction of toxic metals: A review of biological mechanisms. J. Environ. Qual. 2002, 31, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Dushenkov, V.; Motto, H.; Raskin, I. Phytoextraction—The use of plants to remove heavy-metals from soils. Environ. Sci. Technol. 1995, 29, 1232–1238. [Google Scholar] [CrossRef] [PubMed]
- Cen, F.; Chen, L.; Hu, Y.J.; Xu, H. Chelator-induced bioextraction of heavy metals from artificially contaminated soil by mushroom (Coprinus comatus). Chem. Ecol. 2012, 28, 267–280. [Google Scholar] [CrossRef]
- Krishnani, K.K.; Meng, X.G.; Boddu, V.M. Fixation of Heavy Metals onto Lignocellulosic Sorbent Prepared from Paddy Straw. Water Environ. Res. 2008, 80, 2165–2174. [Google Scholar] [CrossRef] [PubMed]
- Dhir, B.; Kumar, R. Adsorption of Heavy Metals by Salvinia Biomass and Agricultural Residues. Int. J. Environ. Res. 2010, 4, 427–432. [Google Scholar]
- Rutkowska, B.; Szulc, W.; Bomze, K.; Gozdowski, D.; Spychaj-Fabisiak, E. Soil factors affecting solubility and mobility of zinc in contaminated soils. Int. J. Environ. Sci. Technol. 2015, 12, 1687–1694. [Google Scholar] [CrossRef]
- Yan, N. Structural Biology of the Major Facilitator Superfamily Transporters. Ann. Rev. Biophys. 2015, 44, 257. [Google Scholar] [CrossRef] [PubMed]
- Balaska, S.; Myrianthopoulos, V.; Tselika, M.; Hatzinikolaou, D.G.; Mikros, E.; Diallinas, G. NmeA, a novel efflux transporter specific for nucleobases and nucleosides, contributes to metal resistance in Aspergillus nidulans. Mol. Microbiol. 2017, 105, 426–439. [Google Scholar] [CrossRef] [PubMed]
- Haydon, M.J.; Cobbett, C.S. A novel major facilitator superfamily protein at the tonoplast influences zinc tolerance and accumulation in Arabidopsis. Plant Physiol. 2007, 143, 1705–1719. [Google Scholar] [CrossRef]
- Pittman, J.K.; Hirschi, K.D. CAX-ing a wide net: Cation/H+ transporters in metal remediation and abiotic stress signalling. Plant Biol. 2016, 18, 741–749. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, I.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Time course analysis of gene regulation under cadmium stress in rice. Plant Soil 2009, 325, 97. [Google Scholar] [CrossRef]
- Li, L.; He, Z.; Pandey, G.K.; Tsuchiya, T.; Luan, S. Functional cloning and characterization of a plant efflux carrier for multidrug and heavy metal detoxification. J. Biol. Chem. 2002, 277, 5360. [Google Scholar] [CrossRef] [PubMed]
- Aiking, H.; Stijnman, A.; Van, G.C.; Van, H.H.; Van’t Riet, J. Inorganic phosphate accumulation and cadmium detoxification in Klebsiella aerogenes NCTC 418 growing in continuous culture. Appl. Environ. Microbiol. 1984, 47, 374–377. [Google Scholar]
- Aragunde, H.; Biarnés, X.; Planas, A. Substrate recognition and specificity of chitin deacetylases and related family 4 carbohydrate esterases. Int. J. Mol. Sci. 2018, 19, 412. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, J.; Farkaš, V.; Sanz, A.B.; Cabib, E. Strengthening the fungal cell wall through chitin-glucan cross-links: Effects on morphogenesis and cell integrity. Cell. Microbiol. 2016, 18, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Balestrini, R.; Bonfante, P. Cell wall remodeling in mycorrhizal symbiosis: A way towards biotrophism. Front. Plant Sci. 2014, 5, 237. [Google Scholar] [CrossRef]
- Di, M.F.; Rapanà, P.; Tomati, U.; Galli, E. Chitin and chitosan from Basidiomycetes. Int. J. Biol. Macromol. 2008, 43, 8–12. [Google Scholar]
- Bhanoori, M.; Venkateswerlu, G. In vivo chitin-cadmium complexation in cell wall of Neurospora crassa. Biochim. Biophys. Acta (BBA) Gen. Subj. 2000, 1523, 21–28. [Google Scholar] [CrossRef]
- Das, S.K.; Guha, A.K. Biosorption of chromium by Termitomyces clypeatus. Colloids Surf. B Biointerfaces 2007, 60, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Strandberg, G.W.; Shumate, S.E.; Parrott, J.R. Microbial cells as biosorbents for heavy metals: Accumulation of uranium by Saccharomyces cerevisiae and Pseudomonas aeruginosa. Appl. Environ. Microbiol. 1981, 41, 237–245. [Google Scholar] [PubMed]
- Helbig, K.; Grosse CNies, D.H. Cadmium toxicity in glutathione mutants of Escherichia coli. J. Bacteriol. 2008, 190, 5439–5454. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Wang, X.; Dou, Y.; Liu, D.; Si, W.; Fang, H.; Zhao, C.; Chen, S.; Xi, J.; Li, J. Hydrogen sulfide—Cysteine cycle system enhances cadmium tolerance through alleviating cadmium-induced oxidative stress and ion toxicity in Arabidopsis roots. Sci. Rep. 2016, 6, 39702–39715. [Google Scholar] [CrossRef] [PubMed]
- Cobbett, C.; Goldsbrough, P. Phytochelatins and metallothioneins: Roles in heavy metal detoxification and homeostasis. Annu. Rev. Plant Biol. 2002, 53, 159–182. [Google Scholar] [CrossRef]
- Zimeri, A.M.; Dhankher, O.P.; McCaig, B.; Meagher, R.B. The plant MT1 metallothioneins are stabilized by binding cadmiums and are required for cadmium tolerance and accumulation. Plant Mol. Biol. 2005, 58, 839–855. [Google Scholar] [CrossRef] [PubMed]
- García, S.; Prado, M.; Dégano, R.; Domínguez, A. A copper-responsive transcription factor, CRF1, mediates copper and cadmium resistance in Yarrowia lipolytica. J. Biol. Chem. 2002, 277, 37359–37368. [Google Scholar] [CrossRef]
- Courbot, M.; Diez, L.; Ruotolo, R.; Chalot, M.; Leroy, P. Cadmium-responsive thiols in the ectomycorrhizal fungus Paxillus involutus. Appl. Environ. Microbiol. 2004, 70, 7413–7417. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. of Reads | Percentage (%) | |||
|---|---|---|---|---|
| Total mapped | 31,654,173–42,895,795 | 70.64–74.32 | ||
| Multiple mapped | 674,534–1,210,904 | 1.82–3.40 | ||
| Uniquely mapped | 30,909,273–40,887,377 | 96.6–98.18 | ||
| CDS | 91.98–92.59 | |||
| Introns | 1.78–1.98 | |||
| Intergenic | 0.21–0.24 | |||
| 5’UTR | 2.45–2.92 | |||
| 3’UTR | 2.72–2.97 | |||
| Unmapped | 11,506,812–15,988,114 | 25.68–29.36 | ||
| Total reads | 43,930,516–58,542,646 |
| Species | Cd Concentration (mg/kg) | pH | Growth Time (day) | Accumulation Concentration (mg/kg Dry Biomass) | Cd Accumulation (μg) | Removal Efficiency per Year (%) | Reference | |
|---|---|---|---|---|---|---|---|---|
| Plant | Cyphomandra betacea | 10 | 7.0 | 40 | Stem 23.46; Leave 24.95; Shoot 24.57 | Stem 12; Leave 39; Shoot 52 | 3.2 | [36] |
| Festuca arundinacea | 500 | Unknown | 90 | Shoot 50 | Shoot 210 | 0.2 | [37] | |
| Amaranthus hypochondriacus | 5 | 7.2 | 60 | Shoot 146.5 | Unknown | 9.3 | [38] | |
| Sedum alfredii | 31.7 | 7.7–7.8 | 118 | Shoot 178.7–212.6 | Shoot 1676.5–2635.2 | 3.3–5.2 | [30] | |
| Brassica napus | 29.4–43.7 | 5.7 | 60 | Shoot 680.6–1221.0 | Unknown | 4.9–7.3 | [39] | |
| Thlaspi caerulescens | 2.9 | 7.3 | 60 | 47.4 | 35 | 3.7 | [40] | |
| Macrofungi | Oudemansiella radicata | 5–30 | 7.5 | 63 | 2.6–18.50 | 69.8–437.9 | 1.6–1.9 | [31] |
| Tricholoma lobynsis | 12 | 7.5 | 45 | 13.67 | 31.7 | 0.3 | [41] | |
| Pleurotus cornucopiae | 5–20 | 7.42 | Unknown | 0.7–3.8 | 11.6–59.9 | Unknown | [14] | |
| Coprinus comatus | 50 | 7.5 | Unknown | 2.8 | 28.2 | Unknown | [42] | |
| Clitocybe maxima | 30 | 6.33 | >35 | 2.5 | 60 | >0.5 | [13] | |
| Armillaria mellea | 0.24, wild | Unknown | Unknown | 1.14 | Unknown | Unknown | [43] | |
| Polvporus squamosus | 0.56 | Unknown | Unknown | 32.4 | Unknown | Unknown | ||
| Oyster mushroom | 150 | 8.92 | 35 | Fruiting body 4.3; substrate 4.88 | Fruiting body 56.3; substrate 660 | 4.6 | This study | |
| 10 | 8.92 | 35 | Fruiting body 1.3; substrate 0.87 | Fruiting body 8.96; substrate 118 | 12.3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, M.; Wang, L.; Hou, J.; Yang, S.; Zheng, X.; Chen, L.; Li, X. Mycoextraction: Rapid Cadmium Removal by Macrofungi-Based Technology from Alkaline Soil. Minerals 2018, 8, 589. https://doi.org/10.3390/min8120589
Chen M, Wang L, Hou J, Yang S, Zheng X, Chen L, Li X. Mycoextraction: Rapid Cadmium Removal by Macrofungi-Based Technology from Alkaline Soil. Minerals. 2018; 8(12):589. https://doi.org/10.3390/min8120589
Chicago/Turabian StyleChen, Miaomiao, Likun Wang, Junliang Hou, Shushen Yang, Xin Zheng, Liang Chen, and Xiaofang Li. 2018. "Mycoextraction: Rapid Cadmium Removal by Macrofungi-Based Technology from Alkaline Soil" Minerals 8, no. 12: 589. https://doi.org/10.3390/min8120589