
Characterization of Extracellular Polymeric Substances Produced by an Acidianus Species and Their Relevance to Bioleaching

Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Cultivation
2.2. Cell Surface Characterization
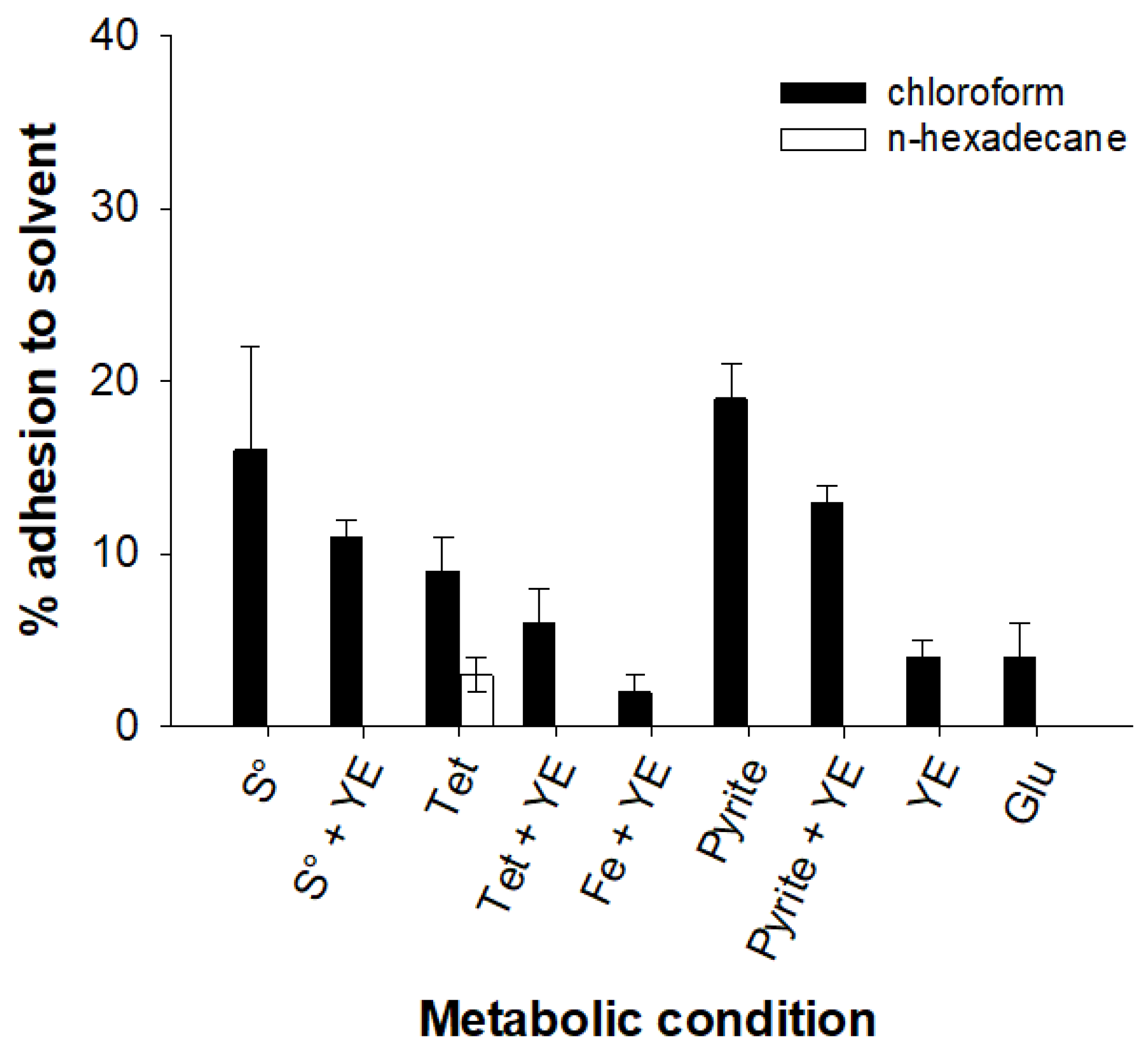
2.2.1. Hydrophobicity
2.2.2. Zeta Potential Determinations
2.2.3. Scanning and Transmission Electron Microscopy (SEM/TEM)
2.3. EPS Extraction
2.4. Characterization of EPS
2.4.1. Biochemical Analyses
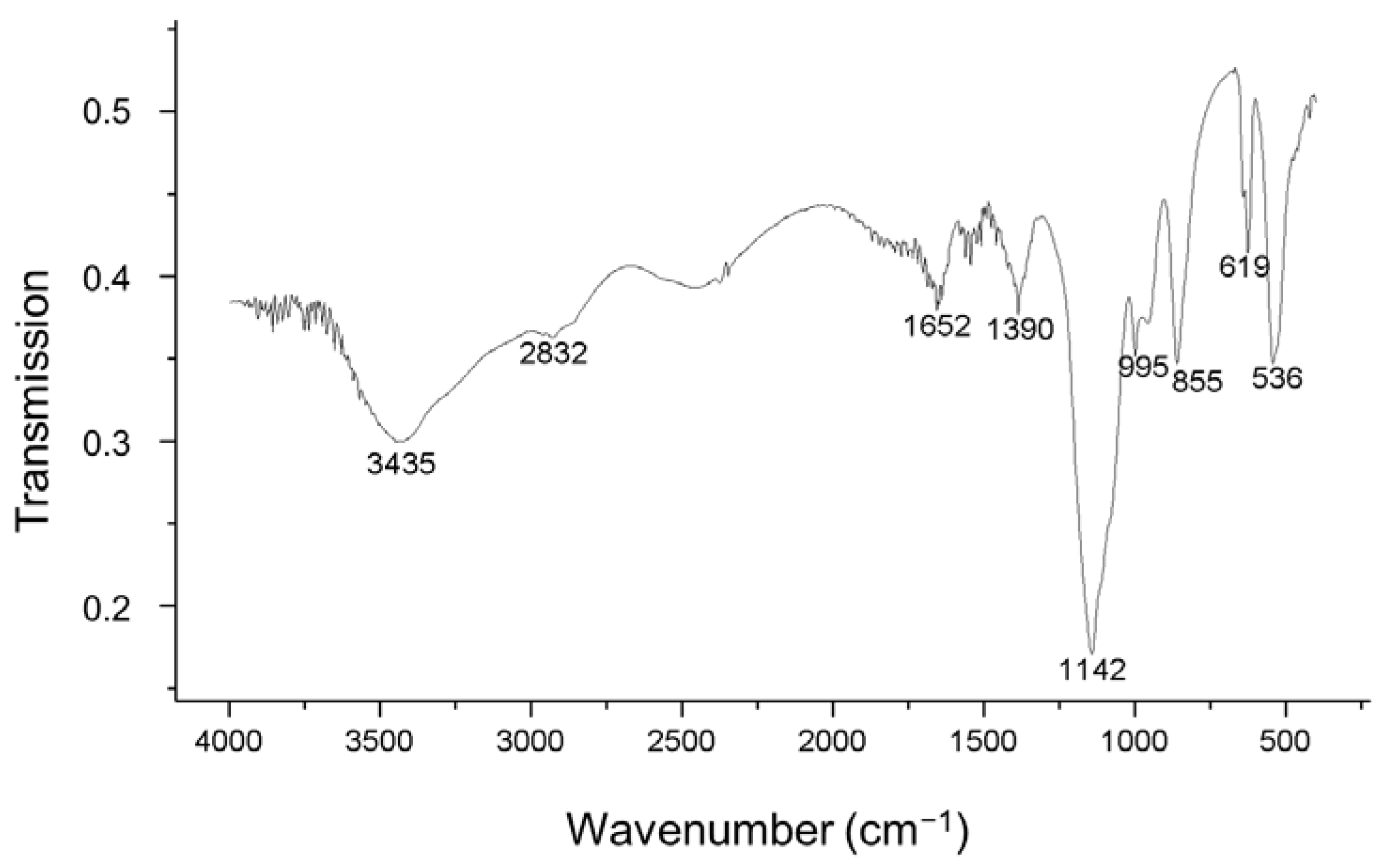
2.4.2. Fourier Transform Infrared (FTIR) Spectroscopy
2.4.3. Fluorescence Lectin-Binding Assays (FLBA)
2.5. Bioleaching Tests
3. Results and Discussion
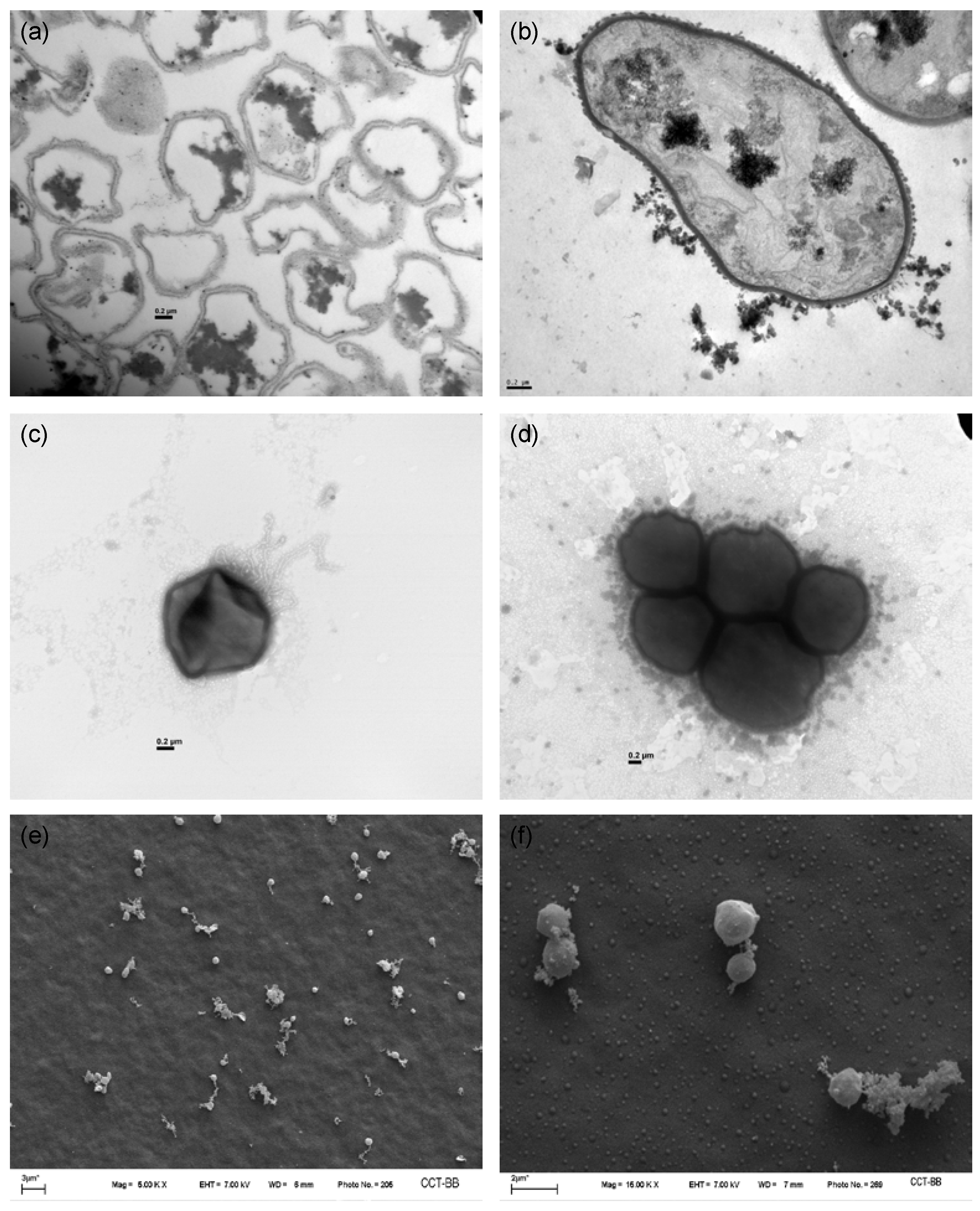
3.1. Cell Morphology
3.2. Cell Surface Characterization
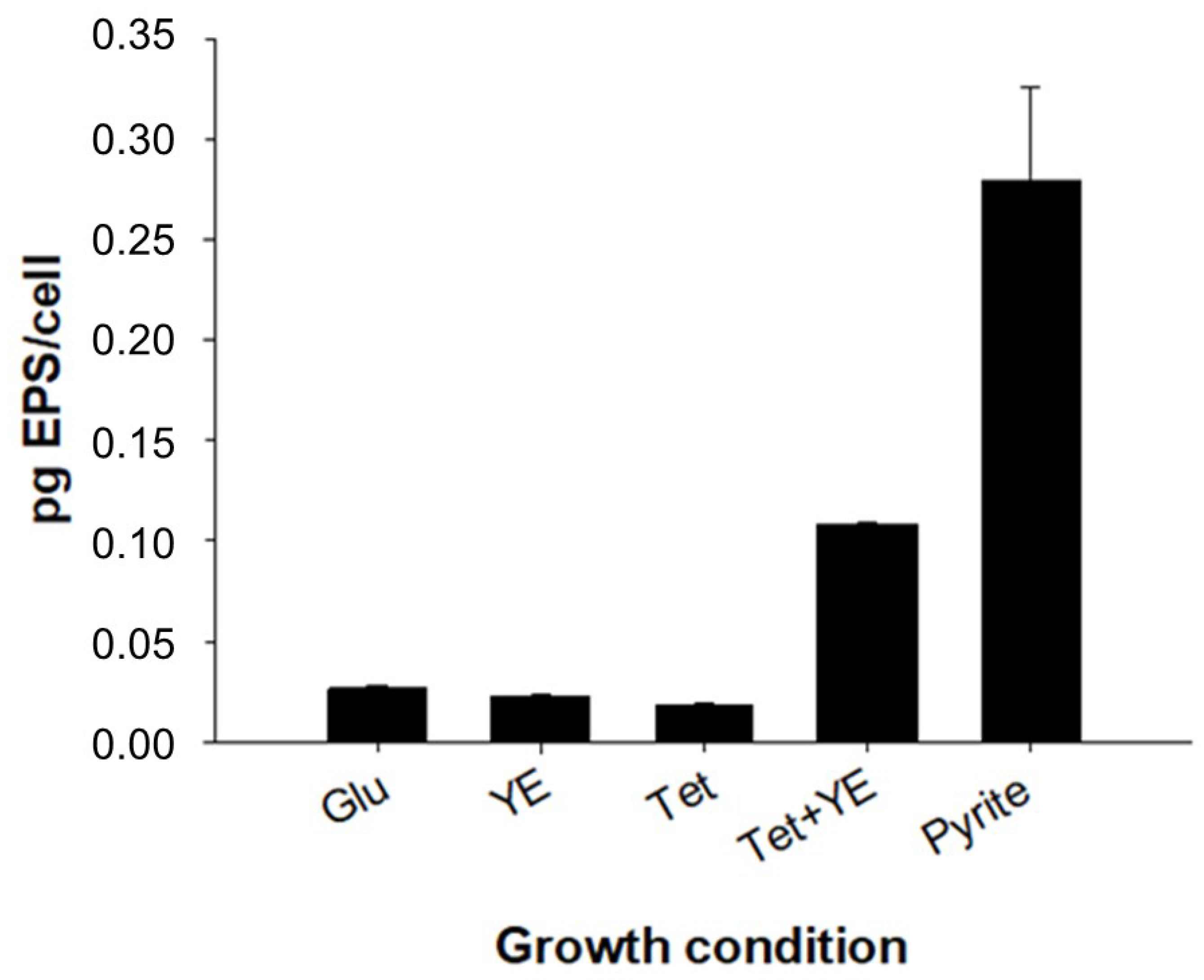
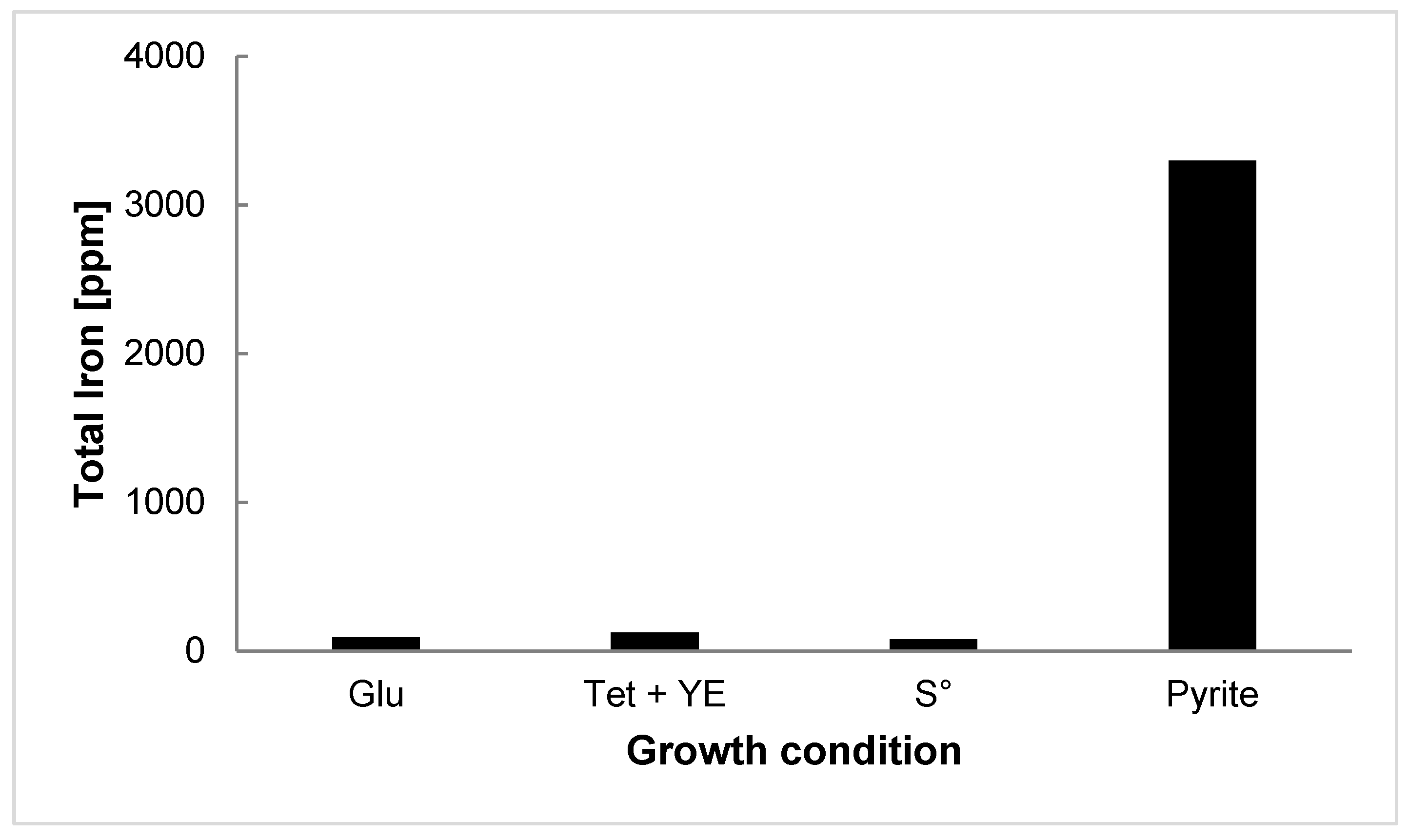
3.3. EPS Production and Characterization
3.4. Pyrite Leaching
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kaksonen, A.H.; Boxall, N.J.; Gumulya, Y.; Khaleque, H.N.; Morris, C.; Bohu, T.; Cheng, K.Y.; Usher, K.M.; Lakaniemi, A.M. Recent Progress in Biohydrometallurgy and Microbial Characterisation. Hydrometallurgy 2018, 180, 7–25. [Google Scholar] [CrossRef]
- Sand, W.; Schippers, A.; Hedrich, S.; Vera, M. Progress in Bioleaching: Fundamentals and Mechanisms of Microbial Metal Sulfide Oxidation–Part A. Appl. Microbiol. Biotechnol. 2022, 106, 6933–6952. [Google Scholar] [CrossRef]
- More, T.T.; Yadav, J.S.S.; Yan, S.; Tyagi, R.D.; Surampalli, R.Y. Extracellular Polymeric Substances of Bacteria and Their Potential Environmental Applications. J. Environ. Manag. 2014, 144, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Castro, C.; Urbieta, M.S.; Plaza Cazón, J.; Donati, E.R. Metal Biorecovery and Bioremediation: Whether or Not Thermophilic Are Better than Mesophilic Microorganisms. Bioresour. Technol. 2019, 279, 317–326. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Yang, Y.; Zhou, S.; Zhong, H.; Sun, W. The Effect of Culture Condition and Ionic Strength on Proton Adsorption at the Surface of the Extreme Thermophile Acidianus Manzaensis. Colloids Surf. B Biointerfaces 2013, 102, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Castro, C.; Zhang, R.; Liu, J.; Bellenberg, S.; Neu, T.R.; Donati, E.; Sand, W.; Vera, M. Biofilm Formation and Interspecies Interactions in Mixed Cultures of Thermo-Acidophilic Archaea Acidianus Spp. and Sulfolobus metallicus. Res. Microbiol. 2016, 167, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Giaveno, M.A.; Urbieta, M.S.; Ulloa, J.R.; González Toril, E.; Donati, E.R. Physiologic Versatility and Growth Flexibility as the Main Characteristics of a Novel Thermoacidophilic Acidianus Strain Isolated from Copahue Geothermal Area in Argentina. Microb. Ecol. 2012, 65, 336–346. [Google Scholar] [CrossRef]
- Sofía Urbieta, M.; Rascovan, N.; Vázquez, M.P.; Donati, E. Genome Analysis of the Thermoacidophilic Archaeon Acidianus Copahuensis Focusing on the Metabolisms Associated to Biomining Activities. BMC Genom. 2017, 18, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Castro, C.; Donati, E.R. Improving Zinc Recovery by Thermoacidophilic Archaeon Acidianus Copahuensis Using Tetrathionate. Trans. Nonferrous Met. Soc. China 2016, 26, 3004–3014. [Google Scholar] [CrossRef]
- Castro, C.; Donati, E. Effects of Different Energy Sources on Cell Adhesion and Bioleaching of a Chalcopyrite Concentrate by Extremophilic Archaeon Acidianus Copahuensis. Hydrometallurgy 2016, 162, 49–56. [Google Scholar] [CrossRef]
- Safar, C.; Castro, C.; Donati, E. Importance of Initial Interfacial Steps during Chalcopyrite Bioleaching by a Thermoacidophilic Archaeon. Microorganisms 2020, 8, 1009. [Google Scholar] [CrossRef] [PubMed]
- Mackintosh, M.E. Nitrogen Fixation by Thiobacillus Ferrooxidans. J. Gen. Microbiol. 1978, 105, 215–218. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Sand, W. Mechanical and Chemical Studies on EPS from Sulfobacillus Thermosulfidooxidans: From Planktonic to Biofilm Cells. Colloids Surf. B Biointerfaces 2017, 153, 34–40. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Blumenkrantz, N.; Asboe-Hansen, G. New Method for Quantitative Determination of Uronic Acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Frølund, B.; Suci, P.A.; Langille, S.; Weiner, R.M.; Geesey, G.G. Influence of Protein Conditioning Films on Binding of a Bacterial Polysaccharide Adhesin from Hyphomonas MHS-3. Biofouling 1996, 10, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, D.H.; Adams, R.B. Colorimetric Determination of Iron in Water With O-Phenanthroline. J. Am. Water Work. Assoc. 1946, 38, 727–730. [Google Scholar] [CrossRef]
- Remonsellez, F.; Orell, A.; Jerez, C.A. Copper Tolerance of the Thermoacidophilic Archaeon Sulfolobus Metallicus: Possible Role of Polyphosphate Metabolism. Microbiology 2006, 152, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Paula, F.S.; Chin, J.P.; Schnürer, A.; Müller, B.; Manesiotis, P.; Waters, N.; Macintosh, K.A.; Quinn, J.P.; Connolly, J.; Abram, F.; et al. The Potential for Polyphosphate Metabolism in Archaea and Anaerobic Polyphosphate Formation in Methanosarcina Mazei. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Toso, D.B.; Javed, M.M.; Czornyj, E.; Gunsalus, R.P.; Hong Zhou, Z.; Zhou, H. Discovery and Characterization of Iron Sulfide and Polyphosphate Bodies Coexisting in Archaeoglobus Fulgidus Cells. Archaea 2016, 2016, 4706532. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Liu, Q.; Wu, X.; Huang, Y.; Wise, M.J.; Liu, Z.; Wang, W.; Hu, J.; Wang, C. Bioinformatics Analysis of Metabolism Pathways of Archaeal Energy Reserves OPEN. Sci. Rep. 2019, 9, 1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorlas, A.; Marguet, E.; Gill, S.; Geslin, C.; Guigner, J.-M.; Guyot, F.; Forterre, P. Sulfur Vesicles from Thermococcales: A Possible Role in Sulfur Detoxifying Mechanisms. Biochimie 2015, 118, 356–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahl, C.; Prange, A. Bacterial Sulfur Globules: Occurrence, Structure and Metabolism. In Inclusions in Prokaryotes; Springer-Verlag: Berlin/Heidelberg, Germany, 2006; pp. 21–51. [Google Scholar]
- Franz, B.; Lichtenberg, H.; Hormes, J.; Modrow, H.; Dahl, C.; Prange, A. Utilization of Solid “elemental” Sulfur by the Phototrophic Purple Sulfur Bacterium Allochromatium Vinosum: A Sulfur K-Edge X-Ray Absorption Spectroscopy Study. Microbiology 2007, 153, 1268–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichinger, I.; Schmitz-Esser, S.; Schmid, M.; Fisher, C.R.; Bright, M. Symbiont-Driven Sulfur Crystal Formation in a Thiotrophic Symbiosis from Deep-Sea Hydrocarbon Seeps. Environ. Microbiol. Rep. 2014, 6, 364–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarrell, K.F.; Albers, S.-V.; Nuno De Sousa Machado, J. A Comprehensive History of Motility and Archaellation in Archaea. FEMS Microbes 2021, 2, 2. [Google Scholar] [CrossRef]
- Albers, S.-V.; Jarrell, K.F. The Archaellum: An Update on the Unique Archaeal Motility Structure. Trends Microbiol. 2018, 26, 351–362. [Google Scholar] [CrossRef]
- Ng, S.Y.M.; Zolghadr, B.; Driessen, A.J.M.; Albers, S.-V.; Jarrell, K.F. Cell Surface Structures of Archaea. J. Bacteriol. 2008, 190, 6039–6047. [Google Scholar] [CrossRef] [Green Version]
- Albers, S.-V.; Meyer, B.H. The Archaeal Cell Envelope. Nat. Rev. Microbiol. 2011, 9, 414–426. [Google Scholar] [CrossRef]
- Wheaton, G.; Counts, J.; Mukherjee, A.; Kruh, J.; Kelly, R. The Confluence of Heavy Metal Biooxidation and Heavy Metal Resistance: Implications for Bioleaching by Extreme Thermoacidophiles. Minerals 2015, 5, 397–451. [Google Scholar] [CrossRef] [Green Version]
- Zolghadr, B.; Klingl, A.; Koerdt, A.; Driessen, A.J.M.; Rachel, R.; Albers, S.-V. Appendage-Mediated Surface Adherence of Sulfolobus Solfataricus. J. Bacteriol. 2010, 192, 104–110. [Google Scholar] [CrossRef] [Green Version]
- Bromfield, L.; Africa, C.-J.; Harrison, S.T.L.; van Hille, R.P. The Effect of Temperature and Culture History on the Attachment of Metallosphaera Hakonensis to Mineral Sulfides with Application to Heap Bioleaching. Miner. Eng. 2011, 24, 1157–1165. [Google Scholar] [CrossRef]
- He, H.; Yang, Y.; Xia, J.; Ding, J.; Zhao, X.; Nie, Z. Growth and Surface Properties of New Thermoacidophilic Archaea Strain Acidianus Manzaensis YN-25 Grown on Different Substrates. Trans. Nonferrous Met. Soc. China 2008, 18, 1374–1378. [Google Scholar] [CrossRef]
- Zhang, R.; Neu, T.R.; Blanchard, V.; Vera, M.; Sand, W. Biofilm Dynamics and EPS Production of a Thermoacidophilic Bioleaching Archaeon. New Biotechnol. 2019, 51, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, H.L.; Newman, D.K.; Kappler, A. Geomicrobiology, 4th ed.; M. Dekker: New York, NY, USA, 2002; ISBN 0824707648. [Google Scholar]
- Ohmura, N.; Kitamura, K.; Saiki, H. Selective Adhesion of Thiobacillus Ferrooxidans to Pyrite. Appl. Environ. Microbiol. 1993, 59, 4050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellenberg, S.; Huynh, D.; Poetsch, A.; Sand, W.; Vera, M. Proteomics Reveal Enhanced Oxidative Stress Responses and Metabolic Adaptation in Acidithiobacillus Ferrooxidans Biofilm Cells on Pyrite. Front. Microbiol. 2019, 10, 592. [Google Scholar] [CrossRef]
- Camacho, D.; Frazao, R.; Fouillen, A.; Nanci, A.; Lang, B.F.; Apte, S.C.; Baron, C.; Warren, L.A. New Insights Into Acidithiobacillus Thiooxidans Sulfur Metabolism Through Coupled Gene Expression, Solution Chemistry, Microscopy, and Spectroscopy Analyses. Front. Microbiol. 2020, 11, 411. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Yang, H.Y.; Lu, L.S.; Sand, W.; Tong, L.L.; Chen, G.B.; Zhao, M.M. miao Interfacial Alteration of Pyrite Caused by Bioleaching. Hydrometallurgy 2020, 195, 105356. [Google Scholar] [CrossRef]
- Devasia, P.; Natarajan, K.A.; Sathyanarayana, D.N.; Rao, G.R. Surface Chemistry of Thiobacillus Ferrooxidans Relevant to Adhesion on Mineral Surfaces. Appl. Environ. Microbiol. 1993, 59, 4051–4055. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, K.A.; Das, A. Surface Chemical Studies on ‘Acidithiobacillus’ Group of Bacteria with Reference to Mineral Flocculation. Int. J. Miner. Process. 2003, 72, 189–198. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, L.; Gu, G.; Hu, Y.; Su, L. Effects of Microorganisms on Surface Properties of Chalcopyrite and Bioleaching. Trans. Nonferrous Met. Soc. China 2008, 18, 1421–1426. [Google Scholar] [CrossRef]
- Liu, H.; Gu, G.; Xu, Y. Surface Properties of Pyrite in the Course of Bioleaching by Pure Culture of Acidithiobacillus Ferrooxidans and a Mixed Culture of Acidithiobacillus Ferrooxidans and Acidithiobacillus Thiooxidans. Hydrometallurgy 2011, 108, 143–148. [Google Scholar] [CrossRef]
- Sharma, P.; Das, A.; Hanumantha Rao, K.; Forssberg, K.S. Surface Characterization of Acidithiobacillus Ferrooxidans Cells Grown under Different Conditions. Hydrometallurgy 2003, 71, 285–292. [Google Scholar] [CrossRef]
- Weerasooriya, R.; Tobschall, H.J. Pyrite–Water Interactions: Effects of PH and PFe on Surface Charge. Colloids Surf. A Physicochem. Eng. Asp. 2005, 264, 68–74. [Google Scholar] [CrossRef]
- Paredes, A.; Acuña, S.M.; Gutiérrez, L.; Toledo, P.G. Zeta Potential of Pyrite Particles in Concentrated Solutions of Monovalent Seawater Electrolytes and Amyl Xanthate. Minerals 2019, 9, 584. [Google Scholar] [CrossRef] [Green Version]
- Rijnaarts, H.H.M.; Norde, W.; Lyklema, J.; Zehnder, A.J.B. The Isoelectric Point of Bacteria as an Indicator for the Presence of Cell Surface Polymers That Inhibit Adhesion. Colloids Surf. B Biointerfaces 1995, 4, 191–197. [Google Scholar] [CrossRef]
- Zhang, R.; Neu, T.R.; Li, Q.; Blanchard, V.; Zhang, Y.; Schippers, A.; Sand, W. Insight into Interactions of Thermoacidophilic Archaea with Elemental Sulfur: Biofilm Dynamics and EPS Analysis. Front. Microbiol. 2019, 10, 896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellenberg, S.; Leon-Morales, C.-F.; Sand, W.; Vera, M. Visualization of Capsular Polysaccharide Induction in Acidithiobacillus Ferrooxidans. Hydrometallurgy 2012, 129–130, 82–89. [Google Scholar] [CrossRef]
- He, Z.; Yang, Y.; Zhou, S.; Hu, Y.; Zhong, H. Effect of Pyrite, Elemental Sulfur and Ferrous Ions on EPS Production by Metal Sulfide Bioleaching Microbes. Trans. Nonferrous Met. Soc. China 2014, 24, 1171–1178. [Google Scholar] [CrossRef]
- Tourney, J.; Ngwenya, B.T. The Role of Bacterial Extracellular Polymeric Substances in Geomicrobiology. Chem. Geol. 2014, 386, 115–132. [Google Scholar] [CrossRef]
- Yu, R.L.; Ou, Y.; Tan, J.X.; Wu, F.D.; Sun, J.; Miao, L.; Zhong, D.L. Effect of EPS on Adhesion of Acidithiobacillus Ferrooxidans on Chalcopyrite and Pyrite Mineral Surfaces. Trans. Nonferrous Met. Soc. China 2011, 21, 407–412. [Google Scholar] [CrossRef]
- Govender, Y.; Gericke, M. Extracellular Polymeric Substances (EPS) from Bioleaching Systems and Its Application in Bioflotation. Miner. Eng. 2011, 24, 1122–1127. [Google Scholar] [CrossRef]
- Rondel, C.; Marcato-Romain, C.-E.; Girbal-Neuhauser, E. Development and Validation of a Colorimetric Assay for Simultaneous Quantification of Neutral and Uronic Sugars. Water Res. 2013, 47, 2901–2908. [Google Scholar] [CrossRef] [PubMed]
- Seviour, T.; Lambert, L.K.; Pijuan, M.; Yuan, Z. Structural Determination of a Key Exopolysaccharide in Mixed Culture Aerobic Sludge Granules Using NMR Spectroscopy. Environ. Sci. Technol. 2010, 44, 8964–8970. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J. The Biofilm Matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Shi, Y.; Huang, J.; Zeng, G.; Gu, Y.; Chen, Y.; Hu, Y.; Tang, B.; Zhou, J.; Yang, Y.; Shi, L. Exploiting Extracellular Polymeric Substances (EPS) Controlling Strategies for Performance Enhancement of Biological Wastewater Treatments: An Overview. Chemosphere 2017, 180, 396–411. [Google Scholar] [CrossRef]
- Barrie Johnson, D.; Hallberg, K.B. Carbon, Iron and Sulfur Metabolism in Acidophilic Micro-Organisms. Adv. Microb. Physiol. 2008, 54, 201–255. [Google Scholar] [CrossRef]
- Liu, L.J.; Jiang, Z.; Wang, P.; Qin, Y.L.; Xu, W.; Wang, Y.; Liu, S.J.; Jiang, C.Y. Physiology, Taxonomy, and Sulfur Metabolism of the Sulfolobales, an Order of Thermoacidophilic Archaea. Front. Microbiol. 2021, 12, 3096. [Google Scholar] [CrossRef]
- Barreto, M.; Jedlicki, E.; Holmes, D.S. Identification of a Gene Cluster for the Formation of Extracellular Polysaccharide Precursors in the Chemolithoautotroph Acidithiobacillus Ferrooxidans. Appl. Environ. Microbiol. 2005, 71, 2902–2909. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Bellenberg, S.; Castro, L.; Neu, T.R.; Sand, W.; Vera, M. Colonization and Biofilm Formation of the Extremely Acidophilic Archaeon Ferroplasma Acidiphilum. Hydrometallurgy 2014, 150, 245–252. [Google Scholar] [CrossRef]
- Orell, A.; Fröls, S.; Albers, S.-V. Archaeal Biofilms: The Great Unexplored. Annu. Rev. Microbiol. 2013, 67, 337–354. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, A.; Aguirre, P.; Gentina, J.C. Biooxidation of Iron by Acidithiobacillus Ferrooxidans in the Presence of D-Galactose: Understanding Its Influence on the Production of EPS and Cell Tolerance to High Concentrations of Iron. Front. Microbiol. 2020, 11, 759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Neu, T.R.; Zhang, Y.; Bellenberg, S.; Kuhlicke, U.; Li, Q.; Sand, W.; Vera, M. Visualization and Analysis of EPS Glycoconjugates of the Thermoacidophilic Archaeon Sulfolobus metallicus. Appl. Microbiol. Biotechnol. 2015, 99, 7343–7356. [Google Scholar] [CrossRef] [PubMed]
- Fröls, S. Archaeal Biofilms: Widespread and Complex. Biochem. Soc. Trans. 2013, 41, 393–398. [Google Scholar] [CrossRef]
- Meyer, B.H.; Albers, S.-V. Hot and Sweet: Protein Glycosylation in Crenarchaeota. Biochem. Soc. Trans. 2013, 41, 384–392. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Guo, G.; Ma, Q.; Zhang, F.; Ma, F.; Liu, J.; Xiao, D.; Yang, X.; Sun, M. Diversity in S-Layers. Prog. Biophys. Mol. Biol. 2017, 123, 1–15. [Google Scholar] [CrossRef]
- Castro, C.; Vera, M.; Donati, E.; Sand, W. Visualization of Attachment and Colonization of Pyrite Surfaces by a Novel Species of Acidianus. In Advanced Materials Research; Trans Tech Publications Ltd.: Wollerau, Switzerland, 2013; Volume 825, ISBN 9783037858912. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Condition | Zeta Potential at pH 2 (mV) | IEP |
|---|---|---|
| S° | 19 ± 4 | 5.2 |
| S° + YE | 7 ± 2 | 4.1 |
| Tet | 11 ± 2 | 4.5 |
| Tet + YE | 14 ± 1 | 4.4 |
| Fe + YE | 6 ± 1 | 4.2 |
| Pyrite | 15 ± 1 | 5.5 |
| Pyrite + YE | 11 ± 1 | 6.0 |
| YE | 2 ± 1 | 4.0 |
| Glu | 7 ± 2 | 3.1 |
| Growth Condition | Proteins | Carbohydrates | Uronic Acids | Humic-Like Substances | Nucleic Acids |
|---|---|---|---|---|---|
| Glu | 66 ± 3 | 580 ± 80 | 70 ± 10 | 38 ± 1 | BDL |
| YE | 105 ± 5 | 52 ± 9 | 40 ± 2 | 130 ± 5 | BDL |
| Tet + YE | 61.2 ± 0.5 | 26 ± 7 | 27 ± 1 | 100 ± 4 | 0.6 ± 0.3 |
| Tet | 10.0 ± 0.5 | BDL | 33 ± 1 | 19 ± 3 | BDL |
| Pyrite (planktonic cells) | 18 ± 1 | 91 ± 20 | 29 ± 2 | 19 ± 4 | BDL |
| Pyrite (sessile cells) | 6 ± 1 | 37 ± 10 | 27 ± 8 | 14 ± 4 | BDL |
| Lectin | Specificities | Growth Condition | |||
|---|---|---|---|---|---|
| S° + YE | Fe + YE | FeS2 | |||
| Planktonic | Planktonic | Planktonic | Sessile | ||
| ConA | D(+)glucose; D(+)mannose | + | + | + | + |
| PNA | D(+)galactose | + | - | - | + |
| LcH | α-mannose; α-glucose | - | + | + | + |
| WGA | N-acetyl-glucosamine | - | - | - | + |
| ECA | N-acetyl-D-galactosamine; D-galactose | - | - | - | - |
| SBA | α- and β-N-acetylgalactosamine | x | + | - | + |
| UEA I | L(-)fucose | - | + | + | + |
| PWM | N-acetyl-glucosamine | + | + | + | + |
| BS I | α-D-galactosyl; N-acetyl-α-D-galactosaminyl | - | + | - | + |
| PHA-E | galactose | - | + | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro, C.; Donati, E.R.; Vera, M. Characterization of Extracellular Polymeric Substances Produced by an Acidianus Species and Their Relevance to Bioleaching. Minerals 2023, 13, 310. https://doi.org/10.3390/min13030310
Castro C, Donati ER, Vera M. Characterization of Extracellular Polymeric Substances Produced by an Acidianus Species and Their Relevance to Bioleaching. Minerals. 2023; 13(3):310. https://doi.org/10.3390/min13030310
Chicago/Turabian StyleCastro, Camila, Edgardo R. Donati, and Mario Vera. 2023. "Characterization of Extracellular Polymeric Substances Produced by an Acidianus Species and Their Relevance to Bioleaching" Minerals 13, no. 3: 310. https://doi.org/10.3390/min13030310