
Isolation and Characterization of A Novel Iron–Sulfur Oxidizing Bacterium Acidithiobacillus Ferrooxidans YQ-N3 and its Applicability in Coal Biodesulfurization

Abstract
:1. Introduction
2. Materials and Methods
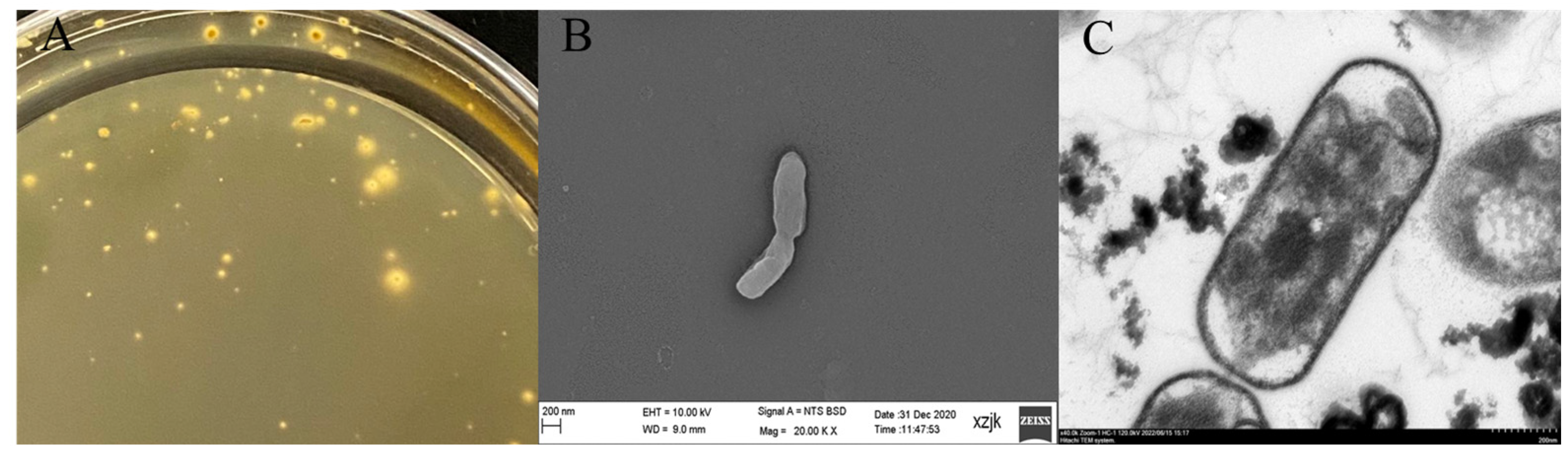
2.1. Isolation, Purification, and Identification
2.2. Whole genome Sequencing
2.3. Comparative Genomic Analysis
2.4. Oxidation Characteristics of Fe2+, S0, and FeS2
2.5. Coal Biodesulfurization Experiment
3. Results and Discussion
3.1. Growth Characteristics of A. ferrooxidans YQ-N3
3.2. Genome Overview of A. ferrooxidans YQ-N3
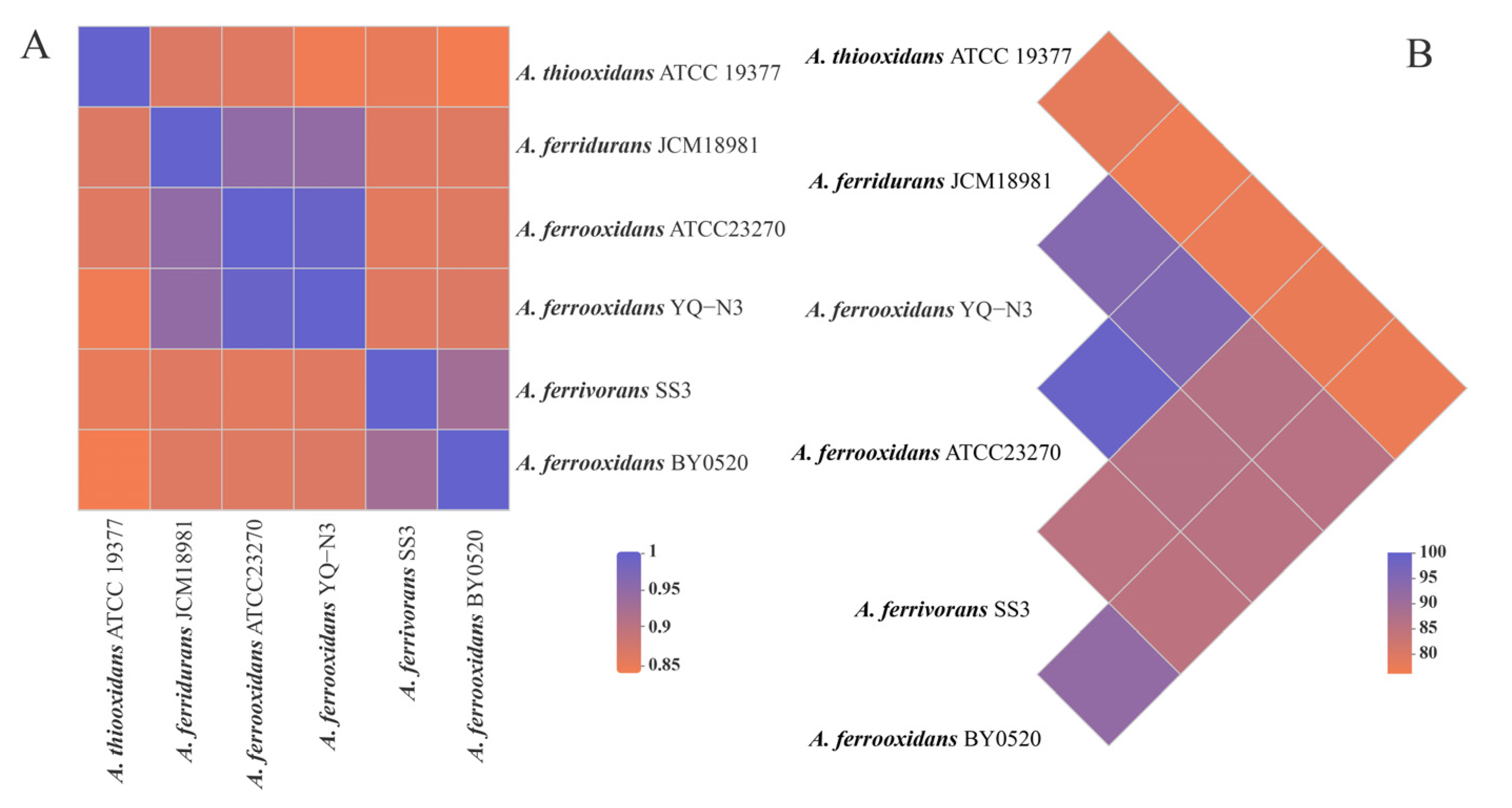
3.3. Comparative Genomic Analysis
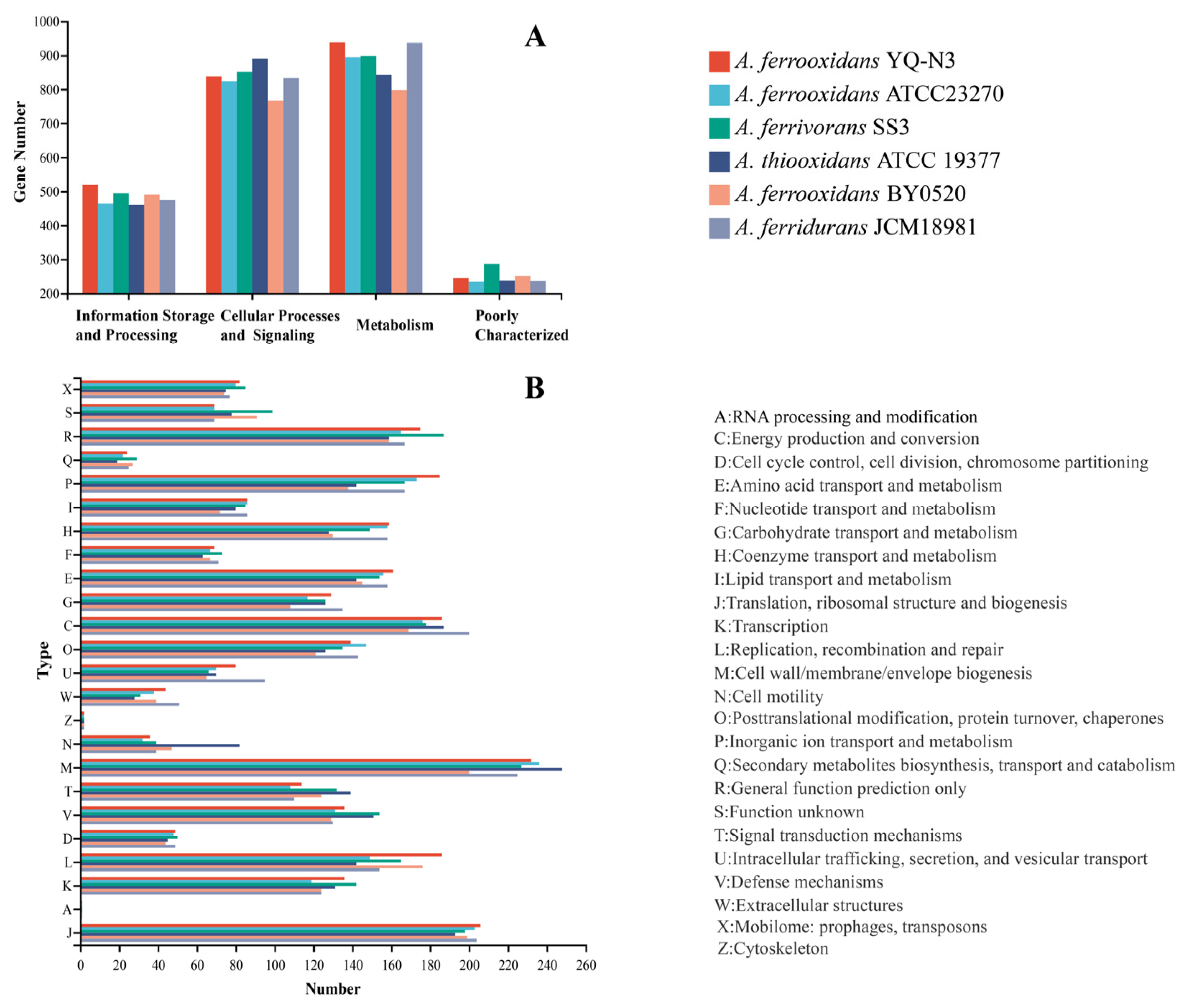
3.4. MEGs
3.5. Genes Associated with Iron and Sulfur Metabolism
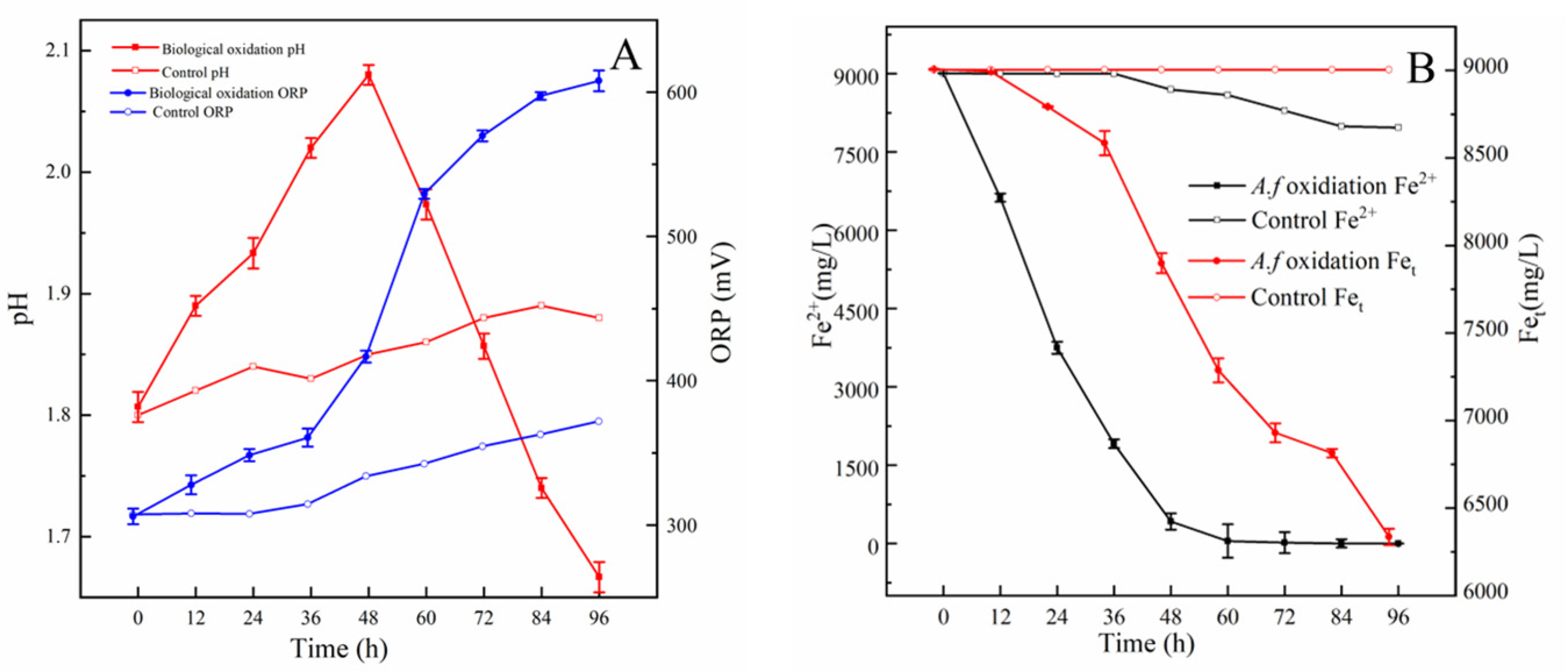
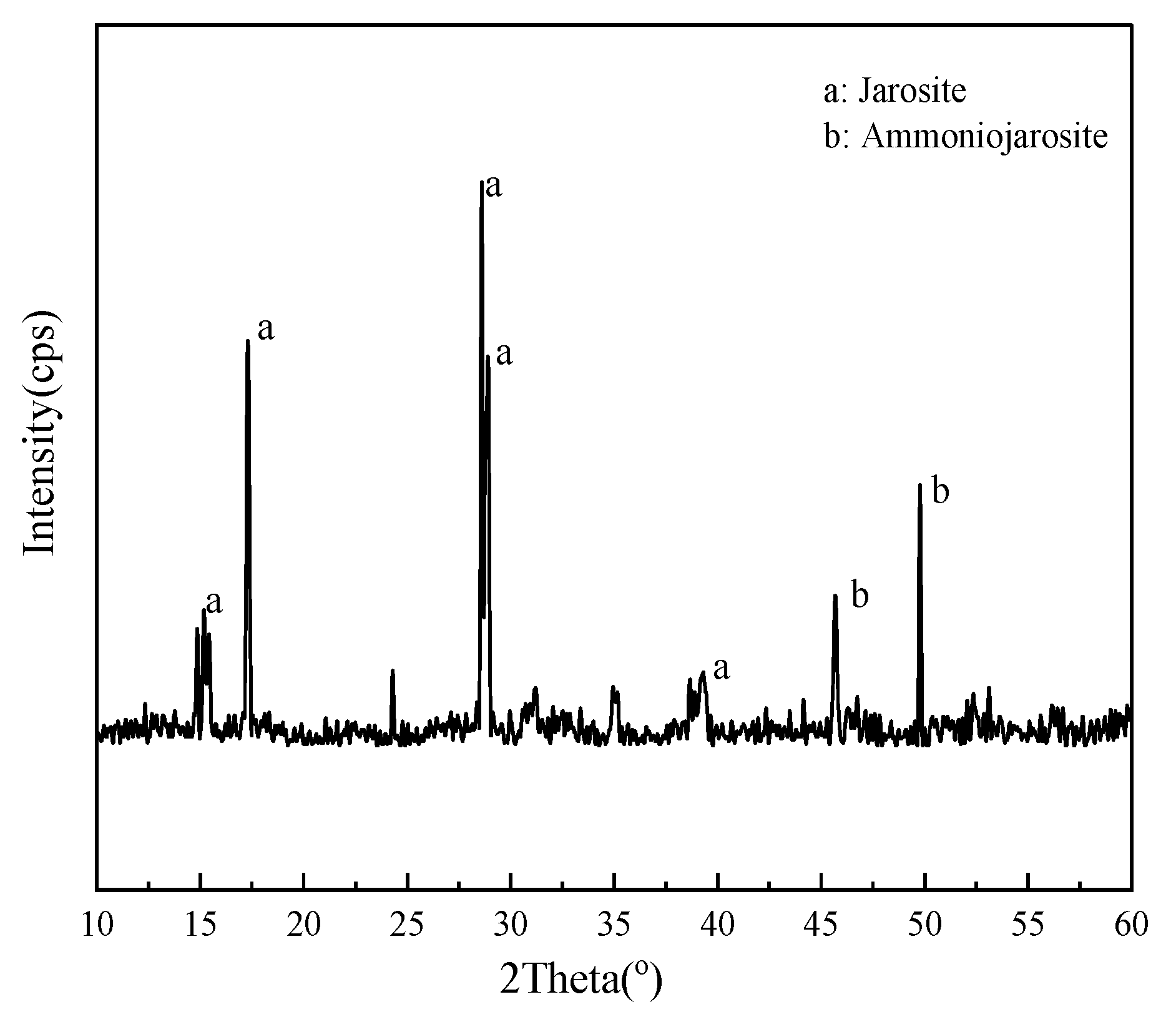
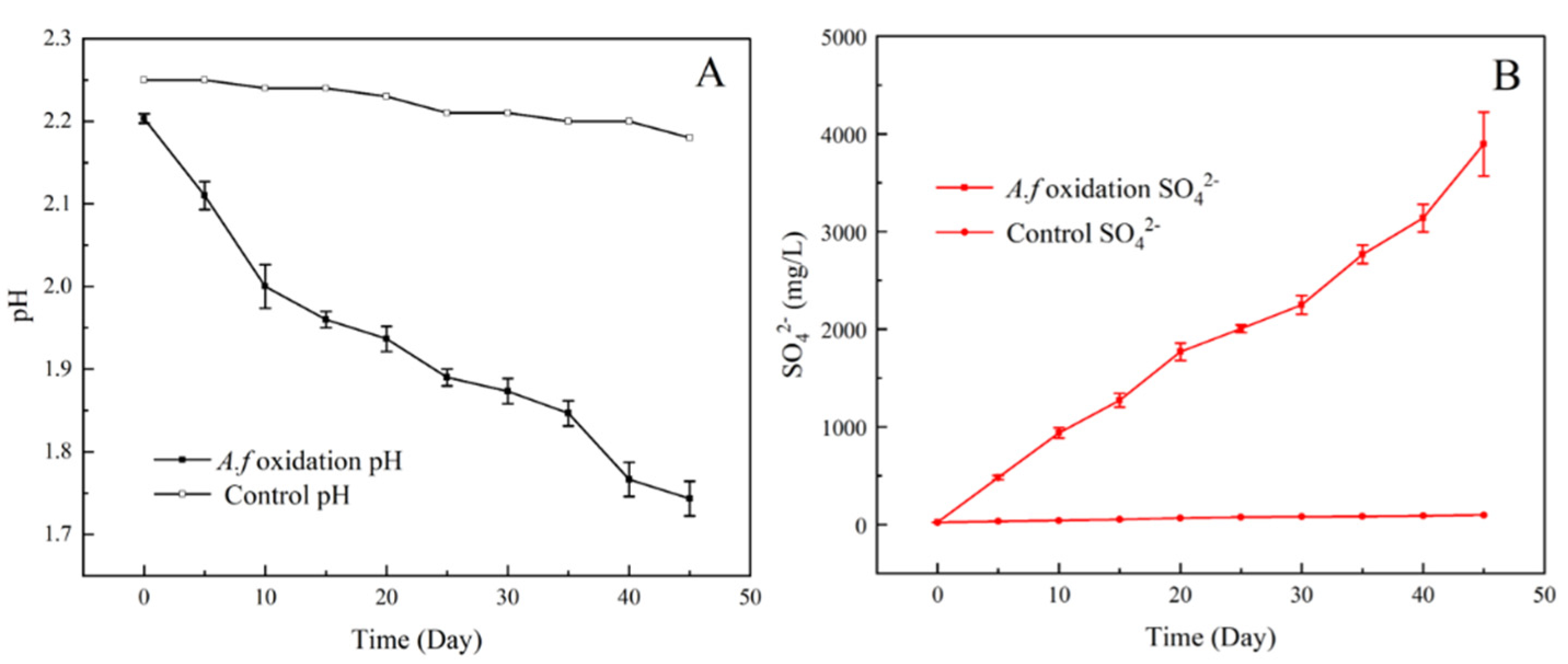
3.6. Oxidation of Fe2+, S0, and Pyrite
3.7. Desulfurization of Coal
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhan, Y.; Yang, M.R.; Zhang, S.; Zhao, D.; Duan, J.G.; Wang, W.D.; Yan, L. Iron and sulfur oxidation pathways of Acidithiobacillus ferrooxidans. World J. Microbiol. Biotechnol. 2019, 35, 12. [Google Scholar] [CrossRef] [PubMed]
- Moinier, D.; Byrne, D.; Amouric, A.; Bonnefoy, V. The Global Redox Responding RegB/RegA Signal Transduction System Regulates the Genes Involved in Ferrous Iron and Inorganic Sulfur Compound Oxidation of the Acidophilic Acidithiobacillus ferrooxidans. Front. Microbiol. 2017, 8, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Yan, L.; Xing, W.J.; Chen, P.; Zhang, Y.; Wang, W.D. Acidithiobacillus ferrooxidans and its potential application. Extremophiles 2018, 22, 563–579. [Google Scholar] [CrossRef]
- Iakovleva, E.; Mäkilä, E.; Salonen, J.; Sitarz, M.; Wang, S.B.; Sillanpää, M. Acid mine drainage (AMD) treatment: Neutralization and toxic elements removal with unmodified and modified limestone. Ecol. Eng. 2014, 81, 30–40. [Google Scholar] [CrossRef]
- Valdes, J.; Pedroso, I.; Quatrini, R.; Dodson, R.J.; Tettelin, H.; Blake, R.; Eisen, J.A.; Holmes, D.S. Acidithiobacillus ferrooxidans metabolism: From genome sequence to industrial applications. BMC Genom. 2008, 9, 24. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo-Tallafigo, J.; Iglesias-González, N.; Mazuelos, A.; Romero, R.; Carranza, F. An alternative approach to recover lead, silver and gold from black gossan (polymetallic ore). Study of biological oxidation and lead recovery stages. J. Clean. Prod. 2019, 207, 510–521. [Google Scholar] [CrossRef]
- Nie, H.Y.; Yang, C.; Zhu, N.W.; Wu, P.X.; Zhang, T.; Zhang, Y.Q.; Xing, Y.J. Isolation of Acidithiobacillus ferrooxidans strain Z1 and its mechanism of bioleaching copper from waste printed circuit boards. J. Chem. Technol. Biotechnol. 2015, 90, 714–721. [Google Scholar] [CrossRef]
- Yang, M.R.; Zhan, Y.; Zhang, S.; Wang, W.D.; Yan, L. Biological materials formed by Acidithiobacillus ferrooxidans and their potential applications. 3 Biotech 2020, 10, 9. [Google Scholar] [CrossRef]
- Rout, P.G.; Mohanty, A.K.; Pradhan, N.; Biswal, S.K.; Behera, S.K. Study on the Reaction Mechanism of Oxidative Microbial Desulfurization of Organic Sulfur-Rich Coal. Geomicrobiol. J. 2022, 39, 210–218. [Google Scholar] [CrossRef]
- Lavalle, L.; Chiacchiarini, P.; Pogliani, C.; Donati, E. Isolation and characterization of acidophilic bacteria from Patagonia, Argentina. Process Biochem. 2005, 40, 1095–1099. [Google Scholar] [CrossRef]
- Kai, M.; Yano, T.; Fukumori, Y.; Yamanaka, T. Cytochrome oxidase of an acidophilic iron-oxidizing bacterium, Thiobacillus ferrooxidans, functions at pH 3.5. Biochem. Biophys. Res. Commun. 1989, 160, 839–843. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, A.; Au, K.F. PacBio Sequencing and Its Applications. Genom. Proteom. Bioinform. 2015, 13, 278–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delcher, A.L.; Bratke, K.A.; Powers, E.C.; Salzberg, S.L. Identifying bacterial genes and endosymbiont DNA with Glimmer. Bioinformatics 2007, 23, 673–679. [Google Scholar] [CrossRef] [Green Version]
- Besemer, J.; Borodovsky, M. GeneMark: Web software for gene finding in prokaryotes, eukaryotes and viruses. Nucleic Acids Res. 2005, 33, W451–W454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertelli, C.; Laird, M.R.; Williams, K.P.; Lau, B.Y.; Hoad, G.; Winsor, G.L.; Brinkman, F.S.L.; Simon Fraser Univ Res Comp, G. IslandViewer 4: Expanded prediction of genomic islands for larger-scale datasets. Nucleic Acids Res. 2017, 45, W30–W35. [Google Scholar] [CrossRef] [PubMed]
- Fouts, D.E. Phage_Finder: Automated identification and classification of prophage regions in complete bacterial genome sequences. Nucleic Acids Res. 2006, 34, 5839–5851. [Google Scholar] [CrossRef] [PubMed]
- Bland, C.; Ramsey, T.L.; Sabree, F.; Lowe, M.; Brown, K.; Kyrpides, N.C.; Hugenholtz, P. CRISPR Recognition Tool (CRT): A tool for automatic detection of clustered regularly interspaced palindromic repeats. BMC Bioinform. 2007, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Cury, J.; Jove, T.; Touchon, M.; Neron, B.; Rocha, E.P.C. Identification and analysis of integrons and cassette arrays in bacterial genomes. Nucleic Acids Res. 2016, 44, 4539–4550. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.J.; Liu, Y.L.; Diep, P.; Mahadevan, R. Genomic Analysis of a Newly Isolated Acidithiobacillus ferridurans JAGS Strain Reveals Its Adaptation to Acid Mine Drainage. Minerals 2021, 11, 14. [Google Scholar] [CrossRef]
- Akcil, A.; Ciftci, H.; Deveci, H. Role and contribution of pure and mixed cultures of mesophiles in bioleaching of a pyritic chalcopyrite concentrate. Miner. Eng. 2007, 20, 310–318. [Google Scholar] [CrossRef]
- Gonzalez-Toril, E.; Llobet-Brossa, E.; Casamayor, E.O.; Amann, R.; Amils, R. Microbial ecology of an extreme acidic environment, the Tinto river. Appl. Environ. Microbiol. 2003, 69, 6959. [Google Scholar] [CrossRef] [Green Version]
- Umanskii, A.B.; Klyushnikov, A.M. Bioleaching of low grade uranium ore containing pyrite using A. ferrooxidans and A. thiooxidans. J. Radioanal. Nucl. Chem. 2013, 295, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, S.; Zhao, D.; Ni, Y.Q.; Wang, W.D.; Yan, L. Complete Genome Sequence of Acidithiobacillus ferrooxidans YNTRS-40, a Strain of the Ferrous Iron- and Sulfur-Oxidizing Acidophile. Microorganisms 2020, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- Hinger, I.; Ansorge, R.; Mussmann, M.; Romano, S. Phylogenomic Analyses of Members of the Widespread Marine Heterotrophic Genus Pseudovibrio Suggest Distinct Evolutionary Trajectories and a Novel Genus, Polycladidibacter gen. nov. Appl. Environ. Microbiol. 2020, 86, 17. [Google Scholar] [CrossRef]
- Hemme, C.L.; Green, S.J.; Rishishwar, L.; Prakash, O.; Pettenato, A.; Chakraborty, R.; Deutschbauer, A.M.; Van Nostrand, J.D.; Wu, L.Y.; He, Z.L.; et al. Lateral Gene Transfer in a Heavy Metal-Contaminated-Groundwater Microbial Community. mBio 2016, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acosta, M.; Beard, S.; Ponce, J.; Vera, M.; Mobarec, J.C.; Jerez, C.A. Identification of putative sulfurtransferase genes in the extremophilic Acidithiobacillus ferrooxidans ATCC 23270 genome: Structural and functional characterization of the proteins. Omics 2005, 9, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Mealman, T.D.; Blackburn, N.J.; McEvoy, M.M. Metal export by CusCFBA, the periplasmic Cu(I)/Ag(I) transport system of Escherichia coli. Curr. Top. Membr. 2012, 69, 163–196. [Google Scholar]
- Chacon, K.N.; Perkins, J.; Mathe, Z.; Alwan, K.; Ho, E.N.; Ucisik, M.N.; Merz, K.M.; Blackburn, N.J. Trapping intermediates in metal transfer reactions of the CusCBAF export pump of Escherichia coli. Commun. Biol. 2018, 1, 11. [Google Scholar] [CrossRef] [Green Version]
- Almarcegui, R.J.; Navarro, C.A.; Paradela, A.; Albar, J.P.; von Bernath, D.; Jerez, C.A. Response to copper of Acidithiobacillus ferrooxidans ATCC 23270 grown in elemental sulfur. Res. Microbiol. 2014, 165, 761–772. [Google Scholar] [CrossRef]
- Wenbin, N.; Dejuan, Z.; Feifan, L.; Lei, Y.; Peng, C.; Xiaoxuan, Y.; Hongyu, L.; Nan, W.B.; Dejuan, L.Z.; Feifan, Y.; et al. Quorum-sensing system in Acidithiobacillus ferrooxidans involved in its resistance to Cu2+. Lett. Appl. Microbiol. 2011, 53, 84–91. [Google Scholar] [CrossRef]
- Popa, O.; Hazkani-Covo, E.; Landan, G.; Martin, W.; Dagan, T. Directed networks reveal genomic barriers and DNA repair bypasses to lateral gene transfer among prokaryotes. Genome Res. 2011, 21, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Polz, M.F.; Alm, E.J.; Hanage, W.P. Horizontal gene transfer and the evolution of bacterial and archaeal population structure. Trends Genet. 2013, 29, 170–175. [Google Scholar] [CrossRef] [Green Version]
- Coutinho, T.J.D.; Franco, G.R.; Lobo, F.P. Homology-Independent Metrics for Comparative Genomics. Comp. Struct. Biotechnol. J. 2015, 13, 352–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominguez-Mirazo, M.; Jin, R.; Weitz, J.S. Functional and comparative genomic analysis of integrated prophage-like sequences in “candidatus liberibacter asiaticus”. mSphere 2019, 4, 409–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cady, K.C.; Bondy-Denomy, J.; Heussler, G.E.; Davidson, A.R.; O’Toole, G.A. The CRISPR/Cas Adaptive Immune System of Pseudomonas aeruginosa Mediates Resistance to Naturally Occurring and Engineered Phages. J. Bacteriol. 2012, 194, 5728–5738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dopson, M.; Holmes, D.S. Metal resistance in acidophilic microorganisms and its significance for biotechnologies. Appl. Microbiol. Biotechnol. 2014, 98, 8133–8144. [Google Scholar] [CrossRef] [PubMed]
- Nies, D.H. Efflux-mediated heavy metal resistance in prokaryotes. Fems Microbiol. Rev. 2003, 27, 313–339. [Google Scholar] [CrossRef]
- Hemme, C.L.; Deng, Y.; Gentry, T.J.; Fields, M.W.; Wu, L.Y.; Barua, S.; Barry, K.; Tringe, S.G.; Watson, D.B.; He, Z.L.; et al. Metagenomic insights into evolution of a heavy metal-contaminated groundwater microbial community. ISME J. 2010, 4, 660–672. [Google Scholar] [CrossRef] [Green Version]
- Hallberg, K.B.; Gonzalez-Toril, E.; Johnson, D.B. Acidithiobacillus ferrivorans, sp nov.; facultatively anaerobic, psychrotolerant iron-, and sulfur-oxidizing acidophiles isolated from metal mine-impacted environments. Extremophiles 2010, 14, 9–19. [Google Scholar] [CrossRef]
- Bonnefoy, V.; Holmes, D.S. Genomic insights into microbial iron oxidation and iron uptake strategies in extremely acidic environments. Environ. Microbiol. 2012, 14, 1597–1611. [Google Scholar] [CrossRef]
- Ponce, J.S.; Moinier, D.; Byrne, D.; Amouric, A.; Bonnefoy, V. Acidithiobacillus ferrooxidans oxidizes ferrous iron before sulfur likely through transcriptional regulation by the global redox responding RegBA signal transducing system. Hydrometallurgy 2012, 127, 187–194. [Google Scholar] [CrossRef]
- Johnson, D.B.; Hallberg, K.B. Carbon, Iron and Sulfur Metabolism in Acidophilic Micro-Organisms. In Advances in Microbial Physiology; Poole, R.K., Ed.; Academic Press Ltd-Elsevier Science Ltd: London, UK, 2009; Volume 54, pp. 201–255. [Google Scholar]
- Bird, L.J.; Bonnefoy, V.; Newman, D.K. Bioenergetic challenges of microbial iron metabolisms. Trends Microbiol. 2011, 19, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Yarzabal, A.; Appia-Ayme, C.; Ratouchniak, J.; Bonnefoy, V. Regulation of the expression of the Acidithiobacillus ferrooxidans rus operon encoding two cytochromes c, a cytochrome oxidase and rusticyanin. Microbiology 2004, 150, 2113–2123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amouric, A.; Appia-Ayme, C.; Yarzabal, A.; Bonnefoy, V. Regulation of the iron and sulfur oxidation pathwayways in the acidophilic A. Ferrooxidans. Adv. Mater. Res. 2009, 71–73, 163–166. [Google Scholar] [CrossRef]
- Kucera, J.; Bouchal, P.; Lochman, J.; Potesil, D.; Janiczek, O.; Zdrahal, Z.; Mandl, M. Ferrous iron oxidation by sulfur-oxidizing Acidithiobacillus ferrooxidans and analysis of the process at the levels of transcription and protein synthesis. Antonie Van Leeuwenhoek 2013, 103, 905–919. [Google Scholar] [CrossRef]
- Wakai, S.; Tsujita, M.; Kikumoto, M.; Manchur, M.A.; Kanao, T.; Kamimura, K. Purification and characterization of sulfide: Quinone oxidoreductase from an acidophilic iron-oxidizing bacterium, acidithiobacillus ferrooxidans. Biosci. Biotechnol. Biochem. 2007, 71, 2735–2742. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.D.; Ji, J.J.; Yu, R.L.; Qiu, G.Z. Expression, Purification and Molecular Modeling of Another HdrC from Acidithiobacillus ferrooxidans Which Binds Only One 4Fe-4S Cluster. Curr. Microbiol. 2012, 65, 416–423. [Google Scholar] [CrossRef]
- Offeddu, F.G.; Cama, J.; Soler, J.M.; Dávila, G.; McDowell, A.; Craciunescu, T. Processes affecting the efficiency of limestone in passive treatments for AMD: Column experiments. J. Environ. Chem. Eng. 2015, 3, 304–316. [Google Scholar] [CrossRef] [Green Version]
- Nazari, B.; Jorjani, E.; Hani, H.; Manfi, Z.; Riahi, A. Formation of jarosite and its effect on important ions for A. ferrooxidans bacteria. Trans. Nonferrous Met. Soc. China 2014, 24, 1152–1160. [Google Scholar] [CrossRef]
- Li, W.B.; Feng, Q.Y.; Liang, H.Q.; Chen, D.; Li, X.D. Passive treatment test of acid mine drainage from an abandoned coal mine in Kaili Guizhou, China. Water Sci. Technol. 2021, 84, 1981–1996. [Google Scholar]
- Konishi, Y.; Asai, S.; Yoshida, N. Growth Kinetics of Thiobacillus thiooxidans on the Surface of Elemental Sulfur. Appl. Environ. Microbiol. 1995, 61, 3617–3622. [Google Scholar] [CrossRef] [Green Version]
- Knickerbocker, C.; Nordstrom, D.K.; Southam, G. The role of “blebbing” in overcoming the hydrophobic barrier during biooxidation of elemental sulfur by Thiobacillus thiooxidans. Chem. Geol. 2000, 169, 425–433. [Google Scholar] [CrossRef]
- Kuang, J.L.; Huang, L.N.; Chen, L.X.; Hua, Z.S.; Li, S.J.; Hu, M.; Li, J.T.; Shu, W.S. Contemporary environmental variation determines microbial diversity patterns in acid mine drainage. ISME J. 2013, 7, 1038–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardona, I.C.; Marquez, M.A. Biodesulfurization of two Colombian coals with native microorganisms. Fuel Process. Technol. 2009, 90, 1099–1106. [Google Scholar] [CrossRef]
- Yang, X.; Wang, S.; Liu, Y.; Zhang, Y. Identification and characterization of Acidithiobacillus ferrooxidans YY2 and its application in the biodesulfurization of coal. Can. J. Microbiol. 2015, 61, 65–71. [Google Scholar] [CrossRef]
- Yang, X.P.; Wang, S.M.; Liu, Y.J.; Liang, Y. A Comparative Study of the Biodesulfurization Efficiency of Acidithiobacillus ferrooxidans LY01 Cells Domesticated with Ferrous Iron and Pyrite. Geomicrobiol. J. 2016, 33, 488–493. [Google Scholar] [CrossRef]
- Newman, D.K. Feasting on Minerals. Science 2010, 327, 793–794. [Google Scholar] [CrossRef]
- Liu, F.W.; Lei, Y.S.; Shi, J.; Zhou, L.X.; Wu, Z.H.; Dong, Y.; Bi, W.L. Effect of microbial nutrients supply on coal bio-desulfurization. J. Hazard. Mater. 2020, 384, 10. [Google Scholar] [CrossRef]
- Arslan, V. The application of combined lignite cleaning processes, bacterial leaching and flotation, for reducing higher ash and sulfur contents. Int. J. Coal Prep. Util. 2022, 42, 2114–2126. [Google Scholar] [CrossRef]
- Silva, R.A.; Park, J.; Ilyas, S.; Borja, D.; Zhao, H.B.; Urik, M.; Rastegar, S.O.; Kim, H. Biodegradation mechanism of arsenopyrite mine tailing with Acidithiobacillus ferrooxidans and influence of ferric supplements. Int. Biodeterior. Biodegrad. 2020, 153, 6. [Google Scholar] [CrossRef]
- Schippers, A. Sand, Bacterial leaching of metal sulfides proceeds by two indirect mechanisms via thiosulfate or via polysulfides and sulfur. Appl. Environ. Microbiol. 1999, 65, 319–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.H.; Huang, X.Y.; He, H.; Tang, J.L.; Tao, X.X.; Huang, H.Z.; Haider, R.; Ali, M.I.; Jamal, A.; Huang, Z.X. Bioleaching Coal Gangue with a Mixed Culture of Acidithiobacillus ferrooxidans and Acidithiobacillus thiooxidans. Minerals 2021, 11, 11. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Energy Source | Bacterial Inoculum |
|---|---|---|
| A.f Oxidation Fe2+ | FeSO4·7H2O (8.95 g) | 10% |
| Control 1 | FeSO4·7H2O (8.95 g) | 0% |
| A.f Oxidation S0 | S0 (0.5 g) | 10% |
| Control 2 | S0 (0.5 g) | 0% |
| A.f Oxidation FeS2 | FeS2 (5 g) | 10% |
| Control 3 | FeS2 (5 g) | 0% |
| Coal Samples | Total S | Sp, Ad | So, Ad | Ss, Ad |
|---|---|---|---|---|
| Ι | 3.46% | 1.02% | 0.81% | 1.63% |
| II | 3.02% | 1.27% | 0.83% | 0.92% |
| III | 2.77% | 0.83% | 1.42% | 0.52% |
| Location | Sequence Length (Bp) | GC Content (%) | Accession Number | Accession Strain Name | Accession Plasmid Name | Identity (%) |
|---|---|---|---|---|---|---|
| Plasmid E | 7910 | 52.4 | - | - | - | - |
| Plasmid D | 23,017 | 60.69 | NC_015188.1 | Acidiphilium multivorum AIU301 | pACMV4 | 88.265 |
| Plasmid C | 29,178 | 62.67 | NC_015178.1 | Acidiphilium multivorum AIU301 | pACMV1 | 88.293 |
| Plasmid B | 34,460 | 60.54 | NC_009470.1 | Acidiphilium cryptum JF-5 | pACRY04 | 96.491 |
| Plasmid A | 79,659 | 61.64 | NC_009469.1 | Acidiphilium cryptum JF-5 | pACRY03 | 99.949 |
| Strain | Geographic Origin | Genome Size (Mb) | GC% | Level | CDS | Genes |
|---|---|---|---|---|---|---|
| A. ferrooxidans YQ-N3 | Shanxi, China | 3.22 | 58.7 | Complete | 3195 | 3252 |
| A. ferrooxidans ATCC23270 | Bituminous coal mine effluent | 2.98 | 58.8 | Complete | 2927 | 3087 |
| A. ferrooxidans BY0502 | Gansu, China | 2.98 | 56.8 | Contig | 3026 | 3186 |
| A. ferridurans JCM18981 | Okayama, Japan | 2.98 | 58.4 | Complete | 2802 | 3043 |
| A. thiooxidans ATCC 19377 | - | 3.42 | 53 | Contig | 3498 | 3584 |
| A. ferrivorans SS3 | Norilsk, Russia | 3.20 | 56.5 | Complete | 3089 | 3200 |
| Strain | Geographic Origin | Highest Desulfurization Rate for Coal | Desulfurization Reaction Time | References |
|---|---|---|---|---|
| A. ferrooxidans YY2 | Guizhou, China | 75% | 30 days | [56] |
| A. ferrooxidans LY01 | Guizhou, China | 67.8% | 13 days | [57] |
| A. ferrooxidans | Johannesburg, South Africa | 79% | 14 days | [58] |
| A. ferrooxidans LX5 | China | 31.6% | 32 days | [59] |
| A. ferrooxidans DSM 583 | - | 30.84% | 12 h | [60] |
| A. ferrooxidans YQ-N3 | Shanxi, China | 62.25% | 30 days | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Feng, Q.; Li, Z. Isolation and Characterization of A Novel Iron–Sulfur Oxidizing Bacterium Acidithiobacillus Ferrooxidans YQ-N3 and its Applicability in Coal Biodesulfurization. Minerals 2023, 13, 95. https://doi.org/10.3390/min13010095
Li W, Feng Q, Li Z. Isolation and Characterization of A Novel Iron–Sulfur Oxidizing Bacterium Acidithiobacillus Ferrooxidans YQ-N3 and its Applicability in Coal Biodesulfurization. Minerals. 2023; 13(1):95. https://doi.org/10.3390/min13010095
Chicago/Turabian StyleLi, Wenbo, Qiyan Feng, and Ze Li. 2023. "Isolation and Characterization of A Novel Iron–Sulfur Oxidizing Bacterium Acidithiobacillus Ferrooxidans YQ-N3 and its Applicability in Coal Biodesulfurization" Minerals 13, no. 1: 95. https://doi.org/10.3390/min13010095