
Vascular Underpinnings of Cerebral Lateralisation in the Neonate
 , , , and
, , , and
Abstract
:1. Introduction
2. Methods
2.1. Participants
2.2. Transcranial Doppler Ultrasound
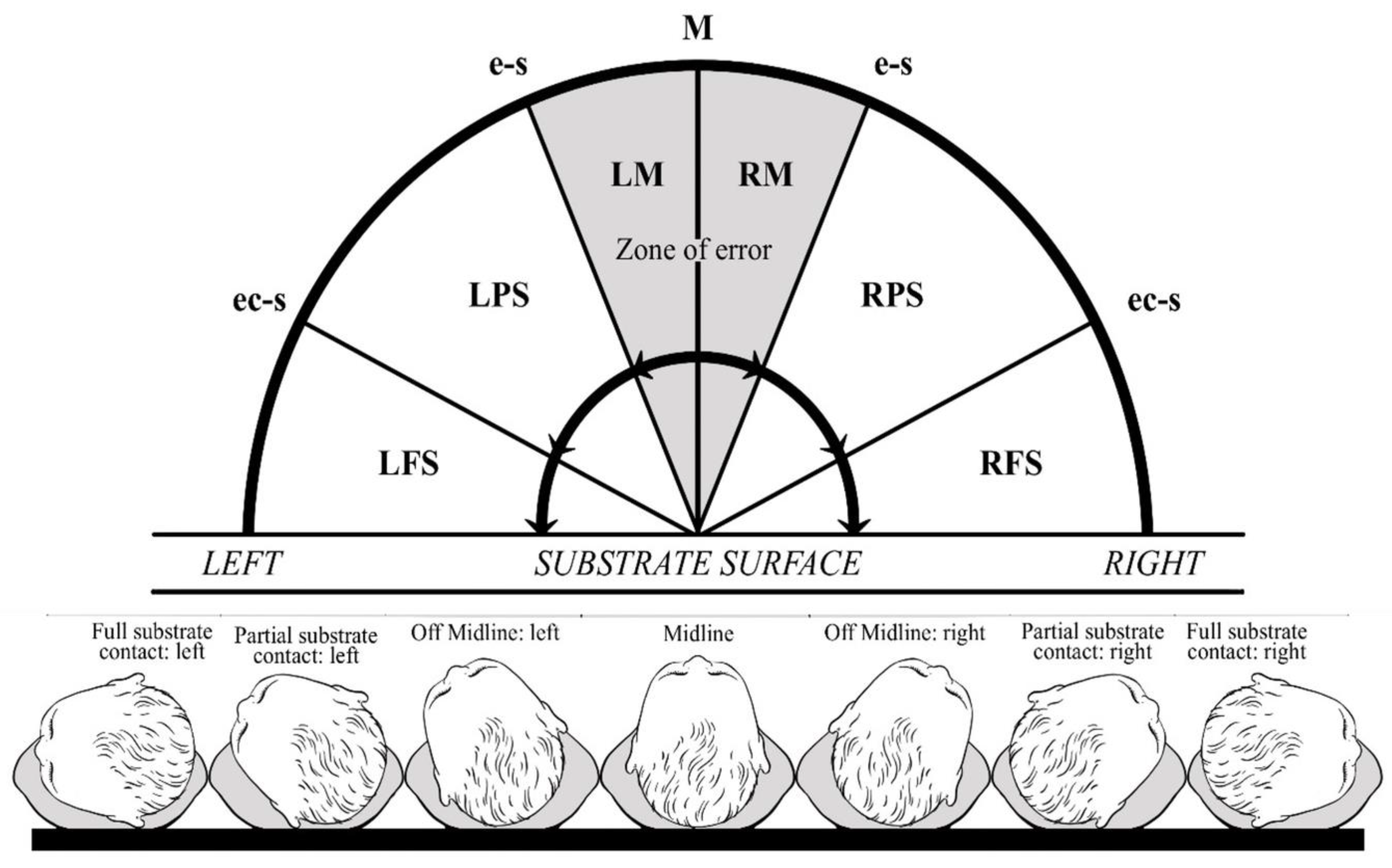
2.3. Neonatal Head Posture
- Midline (M): no left/right rotation.
- Off-midline: left (LM)/right (RM) head rotation without ipsilateral ear contact with the substrate surface.
- Partial substrate contact: left (LPS)/right (RPS) head rotation with ipsilateral ear contact with the substrate surface.
- Full substrate contact: left (LFS)/right (RFS) head rotation with ipsilateral ear and cheek contact with the substrate surface.
2.4. Revised Edinburgh Handedness Inventory
3. Statistical Analysis
4. Results
4.1. Asymmetries in the Origin and Terminus of the Middle Cerebral Artery Trunk
4.2. Predicting the First Head Turn from Arterial Characteristics
4.3. Natural Clusters of Arterial Asymmetry
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jansen van Vuuren, A.; Saling, M.M.; Ameen, O.; Naidoo, N.; Solms, M. Hand preference is selectively related to common and internal carotid arterial asymmetry. Laterality Asymmetry Body Brain Cogn. 2016, 4, 377–398. [Google Scholar] [CrossRef]
- Broca, P. Sur le siège de la faculté du langage articulé. Bull. Société Anat. Paris 1865, 6, 377–393. [Google Scholar] [CrossRef]
- Dax, G. Notes sur le même sujet. Gaz. Hebd. Med. Chir. 1865, 2, 259–262. [Google Scholar]
- Hyrtl, J. Handbuch der Topographischen Anatomie, 4th ed.; Braumüller: Vienna, IT, USA, 1860. [Google Scholar]
- Magendi, F. Recherches Physiologique sur la Vie er la Mort, 4th ed.; Gabou, Bechet Jeuve: Paris, France, 1822. [Google Scholar]
- Ogle, W. On dextral pre-eminence. Trans. R. Med. Chir. Soc. Lond. 1871, 54, 279–301. [Google Scholar] [CrossRef] [PubMed]
- de Fleury, A. Du dynamisme comparé des hémisphères cérèbraux dans l’homme. Association Française Pour l’Avancement des Sciences (comptes-rendus de la première session). Congrès Bordx. 1873, 1, 834–845. [Google Scholar]
- Beeley, A.L. Left-handedness. Am. J. Phys. Anthropol. 1919, 2, 389–400. [Google Scholar] [CrossRef]
- Kellogg, G.M. The physiology of right-and left-handedness. J. Am. Med. Assoc. 1989, 30, 356–358. [Google Scholar] [CrossRef]
- Crichton-Browne, J. Dexterity and the bend sinister. Proc. R. Inst. Great Br. 1907, 18, 623–652. [Google Scholar]
- Geschwind, N.; Levitsky, W. Human brain: Left-right asymmetries in temporal speech region. Science 1968, 161, 186–187. [Google Scholar] [CrossRef] [PubMed]
- Gur, R.C.; Obrist, W.B.; Hungerbuhler, J.P.; Younkin, D.; Rosen, A.D.; Skolnick, B.E.; Reivich, M. Sex and head posture differences in cerebral blood flow during rest and cognitive activity. Science 1982, 217, 659–660. [Google Scholar] [CrossRef]
- Mazziotta, J.C.; Phelps, M.E.; Carson, R.E.; Kuhl, D.E. Tomographic mapping of human cerebral metabolism: Sensory deprivation. Ann. Neurol. 1982, 12, 435–444. [Google Scholar] [CrossRef]
- Wada, J.A.; Clarke, R.; Hamm, A. Cerebral hemispheric asymmetry in humans. Arch. Neurol. 1975, 32, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Chiron, C.; Jambaque, I.; Nabbout, R.; Lounes, R.; Syrota, A.; Dulac, O. The right brain hemisphere is dominant in human infants. Brain 1997, 120, 1057–1065. [Google Scholar] [CrossRef] [PubMed]
- Kamath, S. Observations on the length and diameter of vessels forming the circle of Willis. J. Anat. 1981, 133 Pt 3, 419–423. [Google Scholar] [PubMed]
- Willis, M.W.; Ketter, T.A.; Kimbrell, T.A.; George, M.S.; Herscovitch, P.; Danielson, A.L.; Benson, B.E.; Post, R.M. Age, sex and laterality effects on cerebral glucose metabolism in healthy adults. Psychiatry Res. Neuroimaging 2002, 114, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A. The Anatomy and Physiology of Capillaries; Yale University Press: New Haven, CT, USA, 1929. [Google Scholar]
- Gesell, A.; Ames, L.B. The development of handedness. J. Genet. Psychol. 1947, 70, 155–175. [Google Scholar] [CrossRef] [PubMed]
- Liederman, J. Mechanisms underlying instability in the development of hand preference. In Manual Specialization and the Developing Brain; Young, G., Segalowitz, S., Corter, C., Trehub, S., Eds.; Academic Press: New York, NY, USA, 1983; pp. 71–92. [Google Scholar]
- Saling, M.M. Familial handedness, prenatal environmental adversity, and neonatal lateral organisation. In Manual Specialisation and the Developing Brain; Young, G., Segalowitz, S.J., Corter, C.M., Trehab, S.E., Eds.; Academic Press: New York, NY, USA, 1983; pp. 275–284. [Google Scholar]
- Turkewitz, G.; Creighton, S. Changes in lateral differentiation of head posture in the human neonate. Dev. Psychobiol. 1974, 8, 85–89. [Google Scholar] [CrossRef]
- Coryell, J.; Michel, G.F. How supine postural preference of infants can contribute towards the development of handedness. Infant Behav. Dev. 1978, 1, 245–257. [Google Scholar] [CrossRef]
- Michel, G.F. Right-handedness: A consequence of infant supine head-orientation preference? Science 1981, 212, 685–687. [Google Scholar] [CrossRef]
- Goodwin, R.; Michel, G.F. Head orientation position during birth in neonatal period, and hand preference at 19 weeks. Child Dev. 1981, 52, 819–826. [Google Scholar] [CrossRef]
- Oktar, S.O.; Yücel, C.; Karaosmanoglu, D.; Akkan, K.; Ozdemir, H.; Tokgoz, N.; Tali, T. Blood-flow volume quantification in internal carotid and vertebral arteries: Comparison of 3 different ultrasound techniques with phase contrast MR imaging. Am. J. Neuroradiol. 2006, 27, 363–369. [Google Scholar]
- Kloosterman, A.; Hierck, B.; Westerweel, J.; Poelma, C. Quantification of blood flow topology in developing vascular networks. Plos ONE 2014, 9, 1–15. [Google Scholar]
- Gibbons, G.H.; Dzau, V.J. The emerging concept of vascular remodelling. NEJM 1994, 330, 1431–1438. [Google Scholar] [PubMed]
- Lasjaunias, P.; Bernstein, A.; ter Brugge, K.G. Surgical Neuroangiography; Springer: Berlin/Heidelberg, Germany, 2001. [Google Scholar]
- Drayton, M.R.; Skidmore, R. Vasoactivity of the major intracranial arteries in newborn infants. Arch. Dis. Child. 1987, 62, 236–240. [Google Scholar] [CrossRef] [PubMed]
- van Vuuren, A.J.; Saling, M.; Rogerson, S.; Anderson, P.; Cheong, J.; Solms, M. Cerebral arterial asymmetries in the neonate: Insight into the pathogenesis of stroke. Symmetry 2022, 14, 456. [Google Scholar] [CrossRef]
- Hayashi, T.; Ichiyama, T.; Uchida, M.; Tashiro, N.; Tanaka, H. Evaluation by color Doppler and pulsed Doppler sonography of blood-flow velocities in intracranial-arteries during the early neonatal-period. Eur. J. Pediatr. 1992, 151, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Kempley, S.T.; Vyas, S.; Bower, S.; Nicolaides, K.H.; Gamsu, H. Cerebral and renal artery blood flow velocity before and after birth. Early Hum. Dev. 1996, 46, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Pourcelot, L. L’examen Doppler des Vaisseaux Périphériques; ACD Production: Paris, France, 1982. [Google Scholar]
- Gosling, R.C.; King, D.H. Arterial assessment by Doppler shift ultrasound. Proc. R. Soc. Med. 1988, 67, 447–449. [Google Scholar]
- Wu, Y.; Hsieh, W.; Hsu, C.; Chiu, N.; Chou, H.; Chen, C.; Peng, S.; Hung, H.; Chang, J.; Chen, W.J.; et al. Relationship of neonatal cerebral blood flow velocity asymmetry with early motor, cognitive and language development in term infants. Ultrasound Med. Biol. 2013, 39, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Jahromi, A.S.; Cinà, C.S.; Liu, Y.; Clase, C.M. Sensitivity and specificity of color duplex ultrasound measurement in the estimation of internal carotid artery stenosis: A systematic review and meta-analysis. J. Vasc. Surg. 2005, 41, 962–972. [Google Scholar] [CrossRef]
- Kamouchi, M.; Kishikawa, K.; Okada, Y.; Inoue, T.; Ibayashi, S.; Iida, M. Reappraisal of flow velocity ratio in common carotid artery to predict hemodynamic change in carotid stenosis. Am. J. Neuroradiol. 2005, 26, 957–962. [Google Scholar]
- Hansen, N.B.; Stonestreet, B.S.; Rosenkrantz, T.S.; Oh, W. Validity of Doppler measurements of anterior cerebral artery blood flow velocity: Correlation with brain blood flow in piglets. Pediatrics 1983, 72, 526–531. [Google Scholar] [CrossRef]
- Julkunen, M.; Uotila, J.; Eriksson, K.; Janas, M.; Luukkaala, T.; Tammela, O. Obstetric parameters and Doppler findings in cerebral circulation as predictors of 1 year neurodevelopmental outcome in asphyxiated infants. J. Perinatol. 2012, 32, 631–638. [Google Scholar] [CrossRef]
- Prechtl, H.F.R. The behavioural states of the newborn infant (a review). Brain Res. 1974, 76, 185–212. [Google Scholar] [CrossRef]
- Wolf, P.H. The Causes, Controls, and Organization of Behaviour in the Neonate. Psychol. Issues 1966, 5, 1–105. [Google Scholar]
- Saling, M.M. Determinants of Lateral Organisation in Neonates. Unpublished Doctoral Dissertation, University of Witwatersrand, Johannesburg, South Africa, 1982. [Google Scholar]
- Turkewitz, G. The development of lateral differences in the human infant. In Lateralization in the Nervous System; Harnad, S.R., Doty, R.W., Goldstein, L., Jaynes, J., Krauthamer, G., Eds.; Academic Press: San Diego, CA, USA, 1977; pp. 251–259. [Google Scholar]
- Turkewitz, G.; Birch, H.G. Neurobehavioral organisation of the human newborn. In Exceptional Infant: Studies in Abnormalities; Hellmuth, I., Ed.; Bruner/Mazel: New York, NY, USA, 1971; pp. 24–40. [Google Scholar]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- White, K.; Ashton, R. Handedness assessment inventory. Neuropsychologia 1976, 14, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Edlin, J.M.; Leppanen, M.L.; Fain, R.J.; Hackländer, R.P.; Hanaver-Torrez, S.D.; Lyle, K.B. On the use (and misuse?) if the Edinburgh Handedness Inventory. Brain Cogn. 2015, 94, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.M. Handedness inventories: Edinburgh versus Annett. Neuropsychology 1991, 5, 43–48. [Google Scholar] [CrossRef]
- Huberty, C.J. Discriminant Analysis. Rev. Educ. Res. 1975, 45, 543–598. [Google Scholar] [CrossRef]
- Klecka, W.R. Discriminant Analysis: Quantitative Applications in the Social Sciences Series; Sage Publications: Thousand Oaks, CA, USA, 1980. [Google Scholar]
- Kaufman, L.; Rousseeuw, P.J. Finding Groups in Data: An Introduction to Cluster Analysis; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1990. [Google Scholar]
- Hervé, P.Y.; Zago, L.; Petit, L.; Mazoyer, B.; Tzourio-Mazoyer, N. Revisiting human hemispheric specialization with neuroimaging. Trends Cogn. Sci. 2013, 17, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Galaburda, A.M.; Le May, M.; Kemper, T.L.; Geschwind, N. Right-left asymmetries in the brain. Science 1978, 199, 852–856. [Google Scholar] [CrossRef] [PubMed]
- Sowell, E.R.; Thompson, P.M.; Rex, D.; Kornsand, D.; Tessner, K.D.; Jernigan, T.L.; Toga, A.W. Mapping sulcal pattern asymmetry and local cortical surface gray matter distribution in vivo: Maturation in perisylvian cortices. Cereb. Cortex 2002, 12, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Chi, J.G.; Dooling, E.C.; Gilles, F.H. Left-right asymmetries of the temporal speech areas of the human fetus. Arch. Neurol. 1977, 34, 346–348. [Google Scholar] [CrossRef]
- Dehaene-Lambertz, G.; Hertz-Pannier, L.; Dubois, J. Nature and nurture in language acquisition: Anatomical and functional brain-imaging studies in infants. Trends Neurosci. 2006, 29, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Kinsbourne, M. A model for the ontogeny of cerebral organisation in non-right-handers. In Neuropsychology of Left-Handedness; Herron, J., Ed.; Raven Press: New York, NY, USA, 1980. [Google Scholar]
- Peters, M. Differentiation and lateral specialisation in motor development. In Manual Specialisation and the Developing Brain; Young, G., Segalowitz, S.J., Corter, C.M., Trehab, S.E., Eds.; Academic Press: New York, NY, USA, 1983; pp. 141–159. [Google Scholar]
- Glick, S.D.; Jerussi, T.P.; Zimmerberg, B. Behavioural and neuropharmacological correlates of nigrostriatal asymmetry in rats. In Lateralisation in the Nervous System; Harnad, S., Doty, R.W., Goldstein, L., Jaynes, J., Krauthamer, G., Eds.; Academic Press: New York, NY, USA, 1977. [Google Scholar]
- Andrade, C.; Alwarshetty, M.; Sudha, S.; Chandra, J.S. Effect of innate direction bias on T-maze learning in rats: Implications for research. J. Neurosci. Methods 2001, 110, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.; Martin, L.; Santana, C. Ontogenic development of brain asymmetry in dopaminergic neurons. Brain Res. Bull. 1994, 33, 163. [Google Scholar] [CrossRef] [PubMed]
- de la Fuente-Fernández, R.; Kishore, A.; Calne, D.B.; Ruth, T.J.; Stoessl, A.J. Nigrostriatal dopamine system and motor lateralization. Behav. Brain Res. 2000, 112, 63–68. [Google Scholar] [CrossRef]
- Tomer, R.; Slagter, H.A.; Christian, B.T.; Fox, A.S.; King, C.R.; Murali, D.; Davidson, R.J. Dopamine asymmetries predict orienting bias in healthy individuals. Cereb. Cortex 2013, 23, 2899–2904. [Google Scholar] [CrossRef]
- Scholza, V.H.; Flaherty, A.W.; Kraft, E.; Keltner, J.R.; Kwong, K.K.; Chen, Y.I.; Rosen, B.R.; Jenkins, B.G. Laterality, somatotopy and reproducibility of the basal ganglia and motor cortex during motor tasks. Brain Res. 2000, 879, 204–215. [Google Scholar]
- Kang, X.; Herron, T.J.; Ettlinger, M.; Woods, D.L. Hemispheric asymmetries in cortical and subcortical anatomy. Laterality Asymmetries Body Brain Cogn. 2015, 20, 658–684. [Google Scholar] [CrossRef] [PubMed]
- Teicher, M.H.; Andersen, S.L.; Hostetter, J.C., Jr. Evidence for dopamine receptor pruning between adolescence and adulthood in striatum but not nucleus accumbens. Brain Res. Dev. 1995, 89, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, B.; Lems, Y.L.; van Wulfften Palthe, T.; Hoeksma, J.; Kardaun, O.; Butterworth, G. Development of head position preference during early infancy: A longitudinal study in the daily life situation. Dev. Psychobiol. 1990, 23, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Pries, A.R.; Reglin, B.; Secomb, T.W. Modelling of angioadaptation: Insights for vascular development. Int. J. Dev. Biol. 2011, 55, 399–405. [Google Scholar] [CrossRef]
- van Overbeeke, J.J.; Hillen, B.; Tulleken, C.A.F. A comparative study of the circle of Willis in fetal and adult life: The configuration of the posterior communicating artery. J. Anat. 1991, 178, 45–54. [Google Scholar]
- Rossitti, S.; Lofgren, J. Vascular dimensions of cerebral arteries follow the principle of minimum work. Stroke 1993, 24, 371–377. [Google Scholar] [CrossRef]
- Kasprian, G.; Langs, G.; Brugger, P.C.; Bittner, M.; Weber, M.; Arantes, M.; Prayer, D. The Prenatal origin of hemispheric asymmetry: An in utero neuroimaging study. Cereb. Cortex 2011, 21, 1076–1083. [Google Scholar] [CrossRef]
- De Kovel, C.G.F.; Lisgo, S.N.; Fisher, S.E.; Francks, C. Subtle left-right asymmetry of gene expression profiles in embryonic and foetal human brains. Sci. Rep. 2018, 8, 12606. [Google Scholar] [CrossRef]
- Eichmann, A.; Thomas, J.L. Molecular parallels between neural and vascular development. Cold Spring Harb. Perspect. Med. 2013, 3, a006551. [Google Scholar] [CrossRef]
- Cagnie, B.; Petrovic, M.; Voet, D.; Barbaix, E.; Cambie, D. Vertebral artery dominance and hand preference: Is there a correlation? Man. Ther. 2006, 11, 153–156. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Right Posture | Left Posture | Total | |
|---|---|---|---|
| N a | 55 | 31 | 86 |
| Neonatal Gender (%) | |||
| Male | 54.5 | 64.5 | 58.1 |
| Female | 45.5 | 35.5 | 41.9 |
| Neonatal Characteristics (M, SD) Gestational age at birth (weeks) | 38.9 (1.4) | 39.1 (1.6) | 39.0 (1.4) |
| Birth weight (grams) | 3402.4 (552.3) | 3458.9 (553.0) | 3422.7 (550.09) |
| Age at scan (hours) | 48.1 (28.5) | 51.6 (31.4) | 49.35 (29.4) |
| AS1min b | 9 (1.0) | 9 (1.0) | 9 (1.0) |
| AS5min b | 9 (0.0) | 9 (0.0) | 9 (0.0) |
| Heart rate (beats/minute) | 113.1 (12.1) | 114.4 (13.6) | 113.6 (12.6) |
| Maternal Characteristics (M, SD) Maternal age | 35.1 (4.2) | 34.3 (5.0) | 34.8 (4.5) |
| Maternal LQ | 46.38 (8.45) | 44.52 (12.39) | 45.71 (10.02) |
| Paternal LQ | 45.82 (8.64) | 40.42 (14.01) | 43.87 (11.11) |
| Left Hemisphere | Right Hemisphere | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Artery | Posture | Parameter | M | SD | M | SD | t/Z | df | p | d/r |
| MCAO | Right | Diameter (mm) | 2.15 | 0.35 | 1.98 | 0.32 | 3.100 | 54 | 0.001 * | 0.411 |
| PSV (cm/s) | 53.46 | 10.89 | 52.40 | 12.06 | 0.851 | 54 | 0.199 | 0.116 | ||
| EDV (cm/s) | 18.53 | 5.09 | 18.44 | 5.62 | 0.156 | 54 | 0.438 | 0.021 | ||
| VMEAN (cm/s) | 30.17 | 6.56 | 29.76 | 7.32 | 0.546 | 54 | 0.294 | 0.074 | ||
| RI | 0.65 | 0.07 | 0.65 | 0.07 | 0.627 | 54 | 0.266 | 0.000 | ||
| PI | 0.98 | 0.14 | 0.97 | 0.15 | 0.612 | 54 | 0.272 | 0.095 | ||
| Q (mL/min) | 198.36 | 72.48 | 163.91 | 61.47 | 2.866 | 54 | 0.003 * | 0.388 | ||
| Left | Diameter (mm) | 2.08 | 0.34 | 2.32 | 0.44 | −2.163 | 30 | 0.019 * | −0.394 | |
| PSV (cm/s) | 57.38 | 15.05 | 58.40 | 14.06 | −0.530 | 30 | 0.300 | −0.096 | ||
| EDV (cm/s) | 19.59 | 7.11 | 19.71 | 6.71 | −0.147 | 30 | 0.442 | −0.026 | ||
| VMEAN (cm/s) | 32.19 | 9.27 | 32.61 | 8.82 | −0.387 | 30 | 0.351 | −0.070 | ||
| RI | 0.66 | 0.07 | 0.66 | 0.06 | −0.500 | 30 | 0.310 | 0.000 | ||
| PI | 0.99 | 0.15 | 1.00 | 0.14 | −0.439 | 30 | 0.332 | −0.076 | ||
| Q (mL/min) a | 200.89 | 89.10 | 258.45 | 132.55 | −2.332 | 30 | 0.020 * | −0.296 | ||
| MCADT | Right | Diameter (mm) | 2.04 | 0.34 | 1.89 | 0.33 | 3.070 | 54 | 0.002 * | 0.432 |
| PSV (cm/s) | 52.38 | 11.35 | 51.25 | 13.09 | 0.948 | 54 | 0.174 | 0.129 | ||
| EDV (cm/s) | 17.65 | 5.14 | 17.63 | 5.64 | 0.045 | 54 | 0.482 | 0.005 | ||
| VMEAN (cm/s) | 29.23 | 6.76 | 28.83 | 7.69 | 0.562 | 54 | 0.288 | 0.078 | ||
| RI | 0.66 | 0.06 | 0.66 | 0.07 | 1.556 | 54 | 0.063 | 0.000 | ||
| PI | 1.00 | 0.14 | 0.98 | 0.15 | 1.520 | 54 | 0.067 | 0.238 | ||
| Q (mL/min) | 172.69 | 60.00 | 144.51 | 50.19 | 3.080 | 54 | 0.002 * | 0.418 | ||
| Left | Diameter (mm) | 2.00 | 0.33 | 2.07 | 0.32 | −0.864 | 30 | 0.197 | −0.154 | |
| PSV (cm/s) | 55.69 | 15.24 | 55.77 | 13.60 | −0.059 | 30 | 0.476 | −0.011 | ||
| EDV (cm/s) | 17.80 | 6.65 | 19.01 | 13.60 | −1.398 | 30 | 0.086 | −0.163 | ||
| VMEAN (cm/s) | 30.69 | 9.42 | 31.26 | 8.35 | −0.572 | 30 | 0.286 | −0.104 | ||
| RI | 0.68 | 0.06 | 0.66 | 0.07 | 1.998 | 30 | 0.027 * | 0.333 | ||
| PI | 1.04 | 0.15 | 0.99 | 0.16 | 1.922 | 30 | 0.032 * | 0.345 | ||
| Q (mL/min) a | 179.27 | 79.27 | 192.71 | 77.23 | −1.333 | 30 | 0.183 | −0.167 | ||
| Middle Cerebral Origin | Middle Cerebral Distal Trunk | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Source | df | F | p | ηp2 | df | F | p | ηp2 | |
| Diameter | Diameter | 1 | 0.317 | 0.575 | 0.004 | 1 | 0.672 | 0.415 | 0.008 |
| Posture | 1 | 5.608 | 0.020 * | 0.063 | 1 | 1.462 | 0.230 | 0.017 | |
| Diameter * Posture | 1 | 13.707 | <0.001 * | 0.140 | 1 | 5.989 | 0.016 * | 0.067 | |
| PSV | PSV | 1 | 0.000 | 0.985 | 0.000 | 1 | 0.306 | 0.581 | 0.004 |
| Posture | 1 | 3.576 | 0.062 | 0.041 | 1 | 1.987 | 0.162 | 0.023 | |
| PSV * Posture | 1 | 0.894 | 0.347 | 0.011 | 1 | 0.409 | 0.524 | 0.005 | |
| EDV | EDV | 1 | 0.001 | 0.974 | 0.000 | 1 | 1.632 | 0.205 | 0.019 |
| Posture | 1 | 0.882 | 0.350 | 0.010 | 1 | 0.392 | 0.533 | 0.005 | |
| EDV * Posture | 1 | 0.046 | 0.831 | 0.001 | 1 | 1.758 | 0.189 | 0.020 | |
| VMEAN | VMEAN | 1 | 0.000 | 0.995 | 0.000 | 1 | 0.023 | 0.880 | 0.000 |
| Posture | 1 | 2.259 | 0.137 | 0.026 | 1 | 1.369 | 0.245 | 0.016 | |
| VMEAN * Posture | 1 | 0.413 | 0.522 | 0.005 | 1 | 0.651 | 0.422 | 0.008 | |
| RI | RI | 1 | 0.009 | 0.923 | 0.000 | 1 | 7.953 | 0.006 * | 0.086 |
| Posture | 1 | 0.640 | 0.426 | 0.008 | 1 | 0.684 | 0.411 | 0.008 | |
| RI * Posture | 1 | 0.635 | 0.428 | 0.007 | 1 | 1.890 | 0.173 | 0.022 | |
| PI | PI | 1 | 0.005 | 0.943 | 0.000 | 1 | 7.445 | 0.008 * | 0.081 |
| Posture | 1 | 0.638 | 0.427 | 0.008 | 1 | 0.733 | 0.394 | 0.009 | |
| PI * Posture | 1 | 0.543 | 0.463 | 0.006 | 1 | 1.763 | 0.188 | 0.021 | |
| Q | Q | 1 | 0.744 | 0.391 | 0.009 | 1 | 0.659 | 0.419 | 0.008 |
| Posture | 1 | 11.968 | 0.001 * | 0.125 | 1 | 5.901 | 0.017 * | 0.066 | |
| Q * Posture | 1 | 11.800 | 0.001 * | 0.123 | 1 | 5.258 | 0.024 * | 0.059 | |
| Initial Head Turn | Predicted Group Membership | Total | |||
|---|---|---|---|---|---|
| Right | Left | ||||
| Original a | Count | Right | 47 | 8 | 55 |
| Left | 15 | 16 | 31 | ||
| % | Right | 85.5 | 14.5 | 100 | |
| Left | 48.4 | 51.6 | 100 | ||
| Cross-validated b,c | Count | Right | 47 | 8 | 55 |
| Left | 15 | 16 | 31 | ||
| % | Right | 85.5 | 14.5 | 100 | |
| Left | 48.4 | 51.6 | 100 | ||
| Left Hemisphere | Right Hemisphere | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Artery | Cluster | Postural Tendency | Parameter | M | SD | M | SD | t/Z | df | p | d/r |
| MCAO | 1 | Left | Diameter (mm) | 1.89 | 0.29 | 2.59 | 0.50 | −6.115 | 14 | 0.001 * | −1.710 |
| Q (mL/min) a | 151.93 | 53.04 | 317.83 | 156.08 | 3.409 | 14 | 0.001 * | −0.622 | |||
| 2 | Heterogeneous | Diameter (mm) a | 1.99 | 0.21 | 2.17 | 0.30 | 3.094 | 25 | 0.002 * | −0.429 | |
| Q (mL/min) | 170.76 | 43.57 | 219.64 | 74.20 | −5.575 | 25 | 0.001 * | −1.449 | |||
| 3 | Right | Diameter (mm) | 2.70 | 0.18 | 1.94 | 0.21 | 12.882 | 11 | 0.001 * | 3.733 | |
| Q (mL/min) | 326.56 | 69.28 | 148.58 | 42.50 | 10.371 | 11 | 0.001 * | 3.257 | |||
| 4 | Right | Diameter (mm) | 2.13 | 0.27 | 1.87 | 0.21 | 6.647 | 32 | 0.001 * | 1.214 | |
| Q (mL/min) | 196.96 | 64.46 | 144.42 | 42.63 | 6.238 | 32 | 0.001 * | 1.192 | |||
| MCADT | 1 | Left | Diameter (mm) | 1.80 | 0.29 | 2.34 | 0.38 | −12.477 | 14 | 0.001 * | −3.799 |
| Q (mL/min) | 138.80 | 57.40 | 240.06 | 78.02 | −9.739 | 14 | 0.001 * | −2.889 | |||
| 2 | Heterogeneous | Diameter (mm) | 1.93 | 0.20 | 1.98 | 0.26 | −1.893 | 25 | 0.035 * | −0.380 | |
| Q (mL/min) | 157.32 | 46.00 | 174.86 | 53.84 | −2.962 | 25 | 0.003 * | -0.599 | |||
| 3 | Right | Diameter (mm) | 2.54 | 0.26 | 1.90 | 0.22 | 8.358 | 11 | 0.001 * | 2.461 | |
| Q (mL/min) | 274.85 | 70.51 | 126.09 | 36.04 | 9.096 | 11 | 0.001 * | 3.130 | |||
| 4 | Right | Diameter (mm) | 2.02 | 0.27 | 1.79 | 0.25 | 7.312 | 32 | 0.001 * | 3.137 | |
| Q (mL/min) | 169.23 | 50.47 | 129.14 | 36.32 | 6.806 | 32 | 0.001 * | 1.286 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jansen van Vuuren, A.; Saling, M.; Rogerson, S.; Anderson, P.; Cheong, J.; Solms, M. Vascular Underpinnings of Cerebral Lateralisation in the Neonate. Symmetry 2024, 16, 161. https://doi.org/10.3390/sym16020161
Jansen van Vuuren A, Saling M, Rogerson S, Anderson P, Cheong J, Solms M. Vascular Underpinnings of Cerebral Lateralisation in the Neonate. Symmetry. 2024; 16(2):161. https://doi.org/10.3390/sym16020161
Chicago/Turabian StyleJansen van Vuuren, Anica, Michael Saling, Sheryle Rogerson, Peter Anderson, Jeanie Cheong, and Mark Solms. 2024. "Vascular Underpinnings of Cerebral Lateralisation in the Neonate" Symmetry 16, no. 2: 161. https://doi.org/10.3390/sym16020161