
Striped Expression of Leucine-Rich Repeat Proteins Coordinates Cell Intercalation and Compartment Boundary Formation in the Early Drosophila Embryo

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
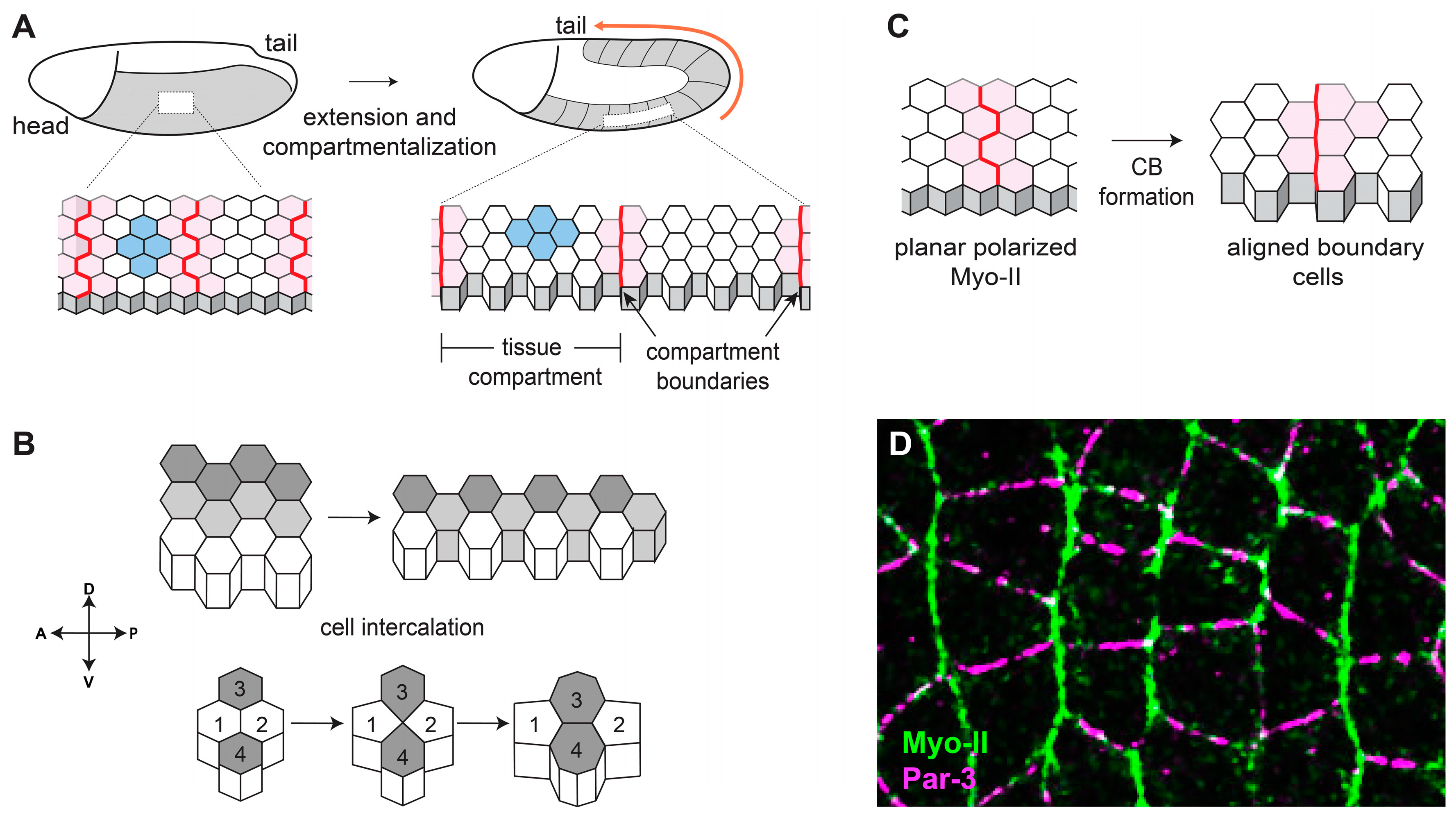
2. Complementary Domains of Cortical Tension and Junctional Adhesion Drive Cell Intercalation
3. Planar Polarity during Drosophila Convergent Extension Requires Patterned Transcriptional Inputs
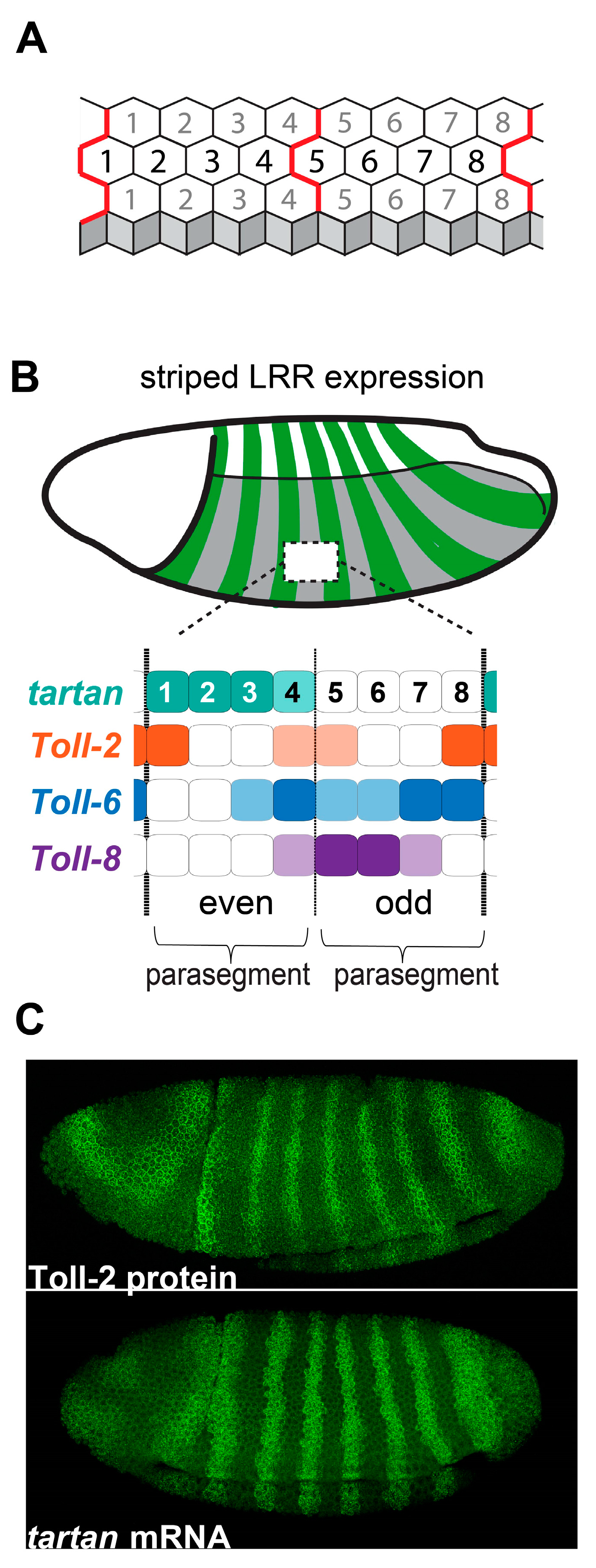
4. Striped Leucine-Rich Repeat Receptors Link Embryonic Patterning to Planar Polarity
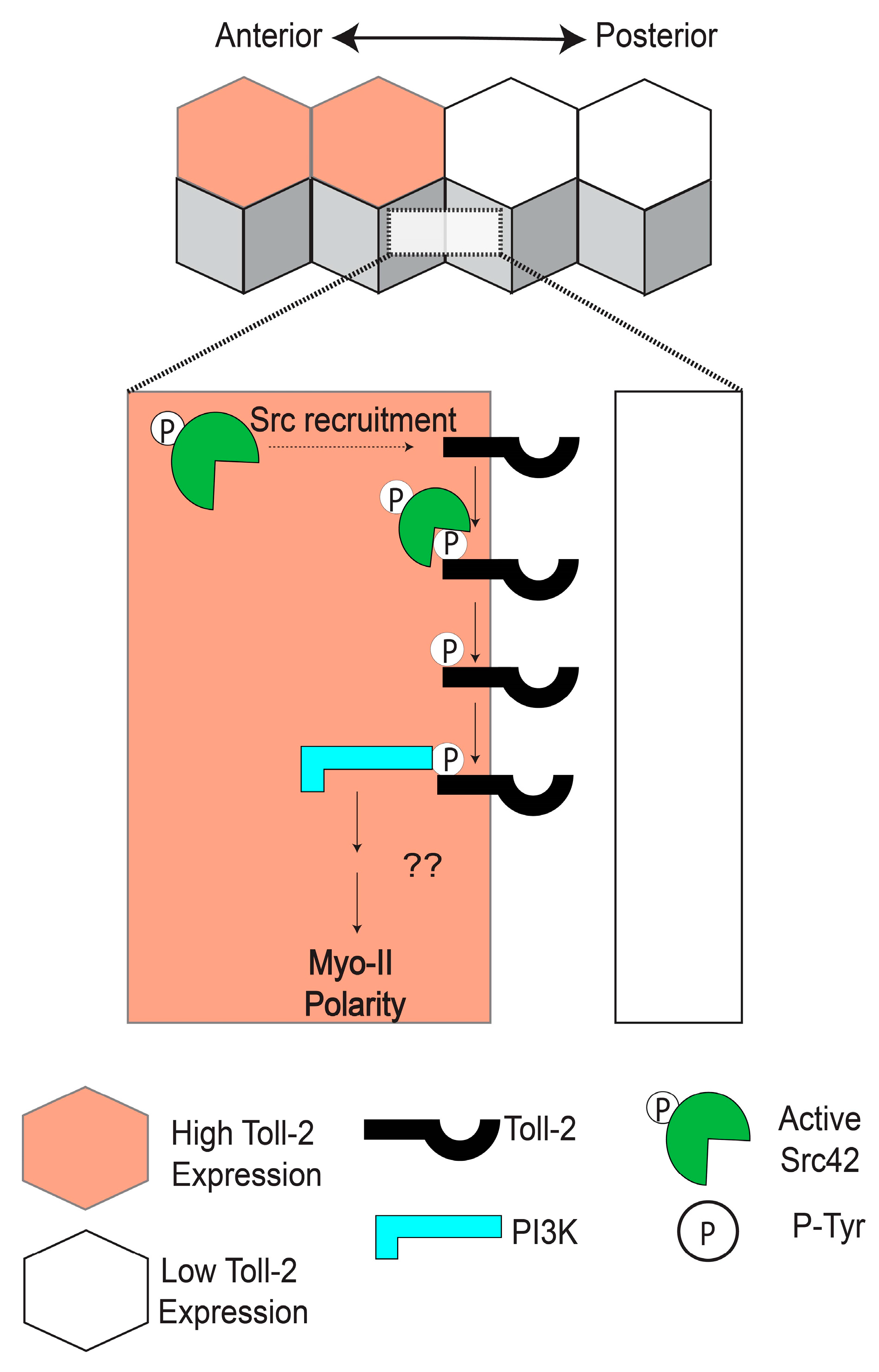
5. Toll-2 Signals through Src and PI3K to Induce Cell Intercalation
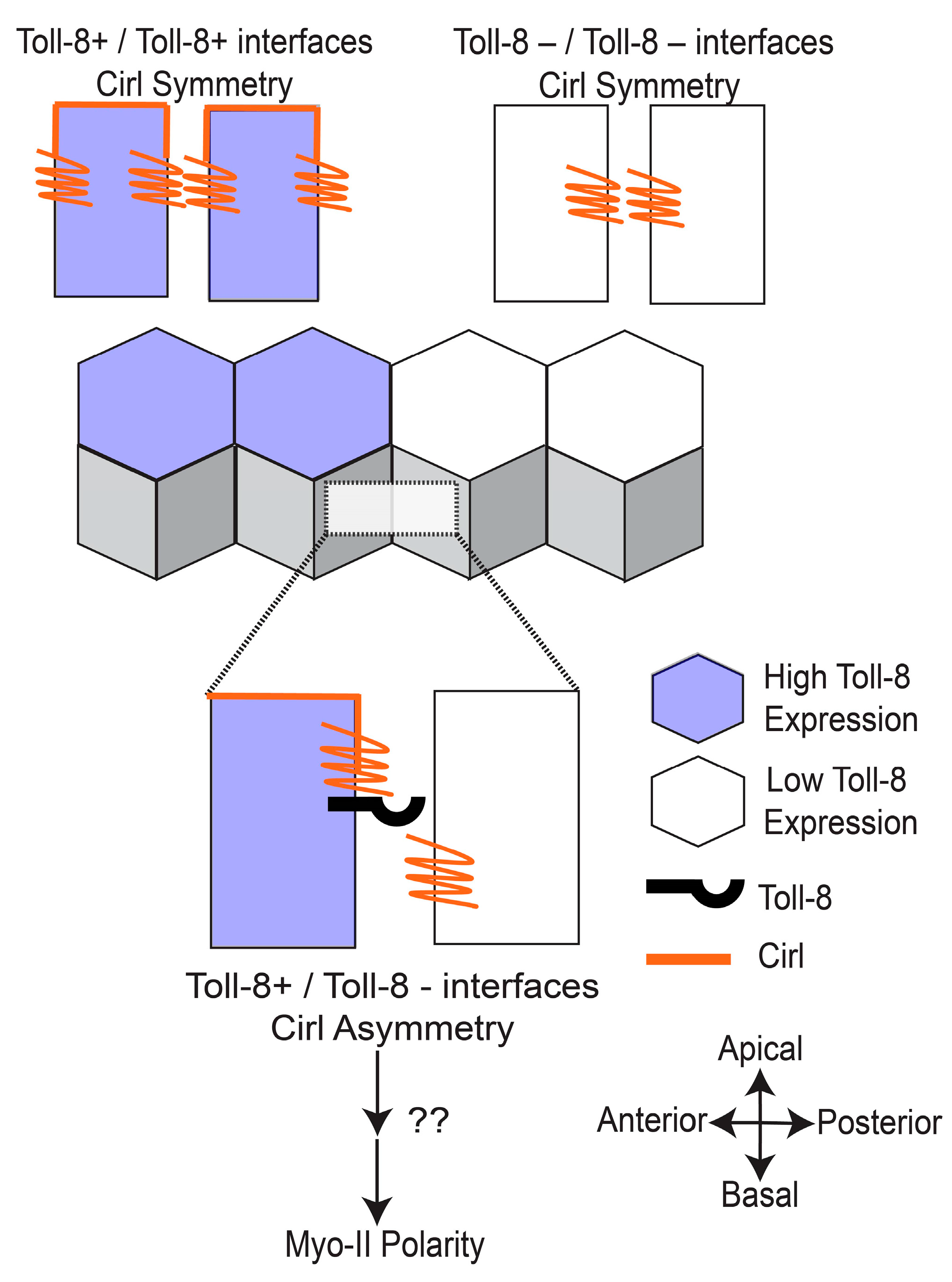
6. Toll-8 Interacts with the GPCR Cirl to Establish Planar Polarity
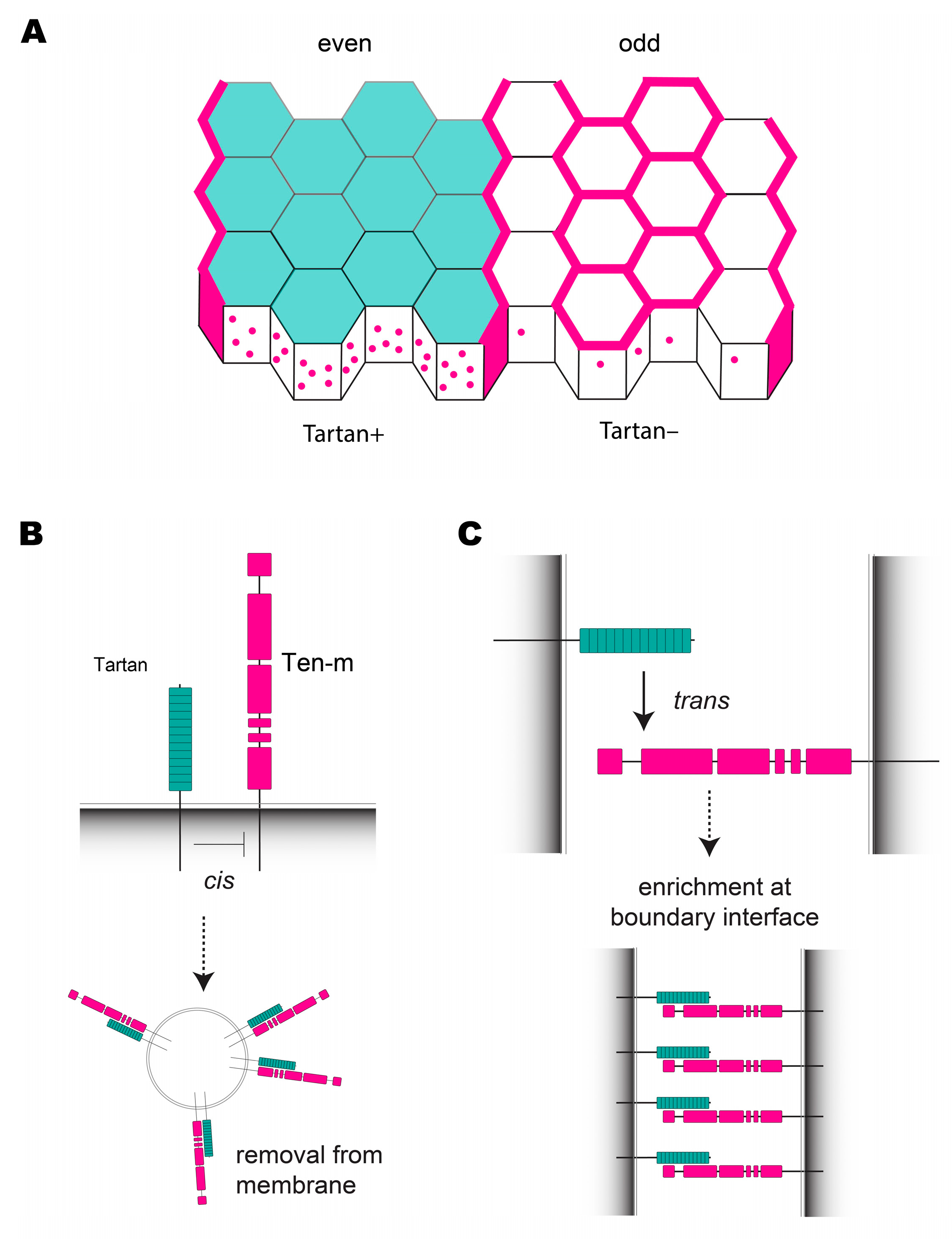
7. Compartment Boundaries in the Early Drosophila Embryo Require Tartan and Ten-m
8. Functional Overlap between LRR Receptor Signaling Pathways during CE
9. Concluding Remarks
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shindo, A. Models of Convergent Extension during Morphogenesis. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, e293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paré, A.C.; Zallen, J.A. Cellular, Molecular, and Biophysical Control of Epithelial Cell Intercalation. In Current Topics in Developmental Biology; Academic Press Inc.: Cambridge, MA, USA, 2020; Volume 136, pp. 167–193. [Google Scholar]
- Keller, R.; Sutherland, A. Convergent Extension in the Amphibian Xenopus Laevis. Curr. Top. Dev. Biol. 2020, 136, 271–317. [Google Scholar] [PubMed]
- Williams, M.L.K.; Solnica-Krezel, L. Cellular and Molecular Mechanisms of Convergence and Extension in Zebrafish. Curr. Top. Dev. Biol. 2020, 136, 377–407. [Google Scholar] [PubMed]
- Sun, Z.; Amourda, C.; Shagirov, M.; Hara, Y.; Saunders, T.E.; Toyama, Y. Basolateral Protrusion and Apical Contraction Cooperatively Drive Drosophila Germ-Band Extension. Nat. Cell Biol. 2017, 19, 375–383. [Google Scholar]
- Fagotto, F. The Cellular Basis of Tissue Separation. Development 2014, 141, 3303–3318. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Dahmann, C. Establishing Compartment Boundaries in Drosophila Wing Imaginal Discs: An Interplay between Selector Genes, Signaling Pathways and Cell Mechanics. Semin. Cell Dev. Biol. 2020, 107, 161–169. [Google Scholar]
- Sharrock, T.E.; Sanson, B. Cell Sorting and Morphogenesis in Early Drosophila Embryos. Semin. Cell Dev. Biol. 2020, 107, 147–160. [Google Scholar]
- Williams, M.; Yen, W.; Lu, X.; Sutherland, A. Distinct Apical and Basolateral Mechanisms Drive Planar Cell Polarity-Dependent Convergent Extension of the Mouse Neural Plate. Dev. Cell 2014, 29, 34–46. [Google Scholar]
- Wallingford, J.B.; Harland, R.M. Neural Tube Closure Requires Dishevelled-Dependent Convergent Extension of the Midline. Development 2002, 129, 5815–5825. [Google Scholar]
- Shi, W.; Peyrot, S.M.; Munro, E.; Levine, M. FGF3 in the Floor Plate Directs Notochord Convergent Extension in the Ciona Tadpole. Development 2009, 136, 23. [Google Scholar]
- Nishimura, T.; Honda, H.; Takeichi, M. Planar Cell Polarity Links Axes of Spatial Dynamics in Neural-Tube Closure. Cell 2012, 149, 1084–1097. [Google Scholar] [CrossRef] [PubMed]
- Irvine, K.D.; Wieschaus, E. Cell Intercalation during Drosophila Germband Extension and Its Regulation by Pair-Rule Segmentation Genes. Development 1994, 120, 841–872. [Google Scholar] [CrossRef]
- Vincent, J.P.; O’Farrell, P.H. The State of Engrailed Expression Is Not Clonally Transmitted during Early Drosophila Development. Cell 1992, 68, 923–931. [Google Scholar] [PubMed] [Green Version]
- Monier, B.; Pélissier-Monier, A.; Brand, A.H.; Sanson, B. An Actomyosin-Based Barrier Inhibits Cell Mixing at Compartmental Boundaries in Drosophila Embryos. Nat. Cell Biol. 2009, 12, 60–65. [Google Scholar] [CrossRef] [Green Version]
- Bertet, C.; Sulak, L.; Lecuit, T. Myosin-Dependent Junction Remodelling Controls Planar Cell Intercalation and Axis Elongation. Nature 2004, 429, 667–671. [Google Scholar] [CrossRef]
- Zallen, J.A.; Wieschaus, E. Patterned Gene Expression Directs Bipolar Planar Polarity in Drosophila. Dev. Cell 2004, 6, 343–355. [Google Scholar]
- Kasza, K.E.; Farrell, D.L.; Zallen, J.A. Spatiotemporal Control of Epithelial Remodeling by Regulated Myosin Phosphorylation. Proc. Natl. Acad. Sci. USA 2014, 111, 11732–11737. [Google Scholar]
- de Matos Simões, S.; Blankenship, J.T.; Weitz, O.; Farrell, D.L.; Tamada, M.; Fernandez-Gonzalez, R.; Zallen, J.A. Rho-Kinase Directs Bazooka/Par-3 Planar Polarity during Drosophila Axis Elongation. Dev. Cell 2010, 19, 377–388. [Google Scholar]
- Rauzi, M.; Lenne, P.-F.; Lecuit, T. Planar Polarized Actomyosin Contractile Flows Control Epithelial Junction Remodelling. Nature 2010, 468, 1110–1114. [Google Scholar]
- Amano, M.; Ito, M.; Kimura, K.; Fukata, Y.; Chihara, K.; Nakano, T.; Matsuura, Y.; Kaibuchi, K. Phosphorylation and Activation of Myosin by Rho-Associated Kinase (Rho- Kinase). J. Biol. Chem. 1996, 271, 20246–20249. [Google Scholar] [CrossRef] [Green Version]
- de Matos Simões, S.; Mainieri, A.; Zallen, J.A. Rho GTPase and Shroom Direct Planar Polarized Actomyosin Contractility during Convergent Extension. J. Cell Biol. 2014, 204, 575–589. [Google Scholar] [CrossRef]
- Garcia De Las Bayonas, A.; Philippe, J.-M.; Lellouch, A.C.; Lecuit, T. Distinct RhoGEFs Activate Apical and Junctional Contractility under Control of G Proteins during Epithelial Morphogenesis. Curr. Biol. 2019, 29, 3370–3385.e7. [Google Scholar] [PubMed] [Green Version]
- Pickering, K.; Alves-Silva, J.; Goberdhan, D.; Millard, T.H. Par3/Bazooka and Phosphoinositides Regulate Actin Protrusion Formation during Drosophila Dorsal Closure and Wound Healing. Development 2013, 140, 800–809. [Google Scholar]
- Tamada, M.; Zallen, J.A. Square Cell Packing in the Drosophila Embryo through Spatiotemporally Regulated EGF Receptor Signaling. Dev. Cell 2015, 35, 151–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.C.; Balaghi, N.; Erdemci-Tandogan, G.; Castle, V.; Fernandez-Gonzalez, R. Myosin Cables Control the Timing of Tissue Internalization in the Drosophila Embryo. Cells Dev. 2021, 168, 203721. [Google Scholar] [CrossRef]
- Strutt, H.; Strutt, D. Asymmetric Localisation of Planar Polarity Proteins: Mechanisms and Consequences. Semin. Cell Dev. Biol. 2009, 20, 957–963. [Google Scholar] [PubMed]
- Goodrich, L.V.; Strutt, D. Principles of Planar Polarity in Animal Development. Development 2011, 138, 1877–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devenport, D. The Cell Biology of Planar Cell Polarity. J. Cell Biol. 2014, 207, 171–179. [Google Scholar]
- Yang, Y.; Mlodzik, M. Wnt-Frizzled/Planar Cell Polarity Signaling: Cellular Orientation by Facing the Wind (Wnt). Annu. Rev. Cell Dev. Biol. 2015, 31, 623–646. [Google Scholar]
- Paré, A.C.; Vichas, A.; Fincher, C.T.; Mirman, Z.; Farrell, D.L.; Mainieri, A.; Zallen, J.A. A Positional Toll Receptor Code Directs Convergent Extension in Drosophila. Nature 2014, 515, 523–527. [Google Scholar]
- Nüsslein-volhard, C.; Wieschaus, E. Mutations Affecting Segment Number and Polarity in Drosophila. Nature 1980, 287, 795–801. [Google Scholar] [PubMed]
- Kimelman, D.; Martin, B.L. Anterior-Posterior Patterning in Early Development: Three Strategies. Wiley Interdiscip. Rev. Dev. Biol. 2012, 1, 253–266. [Google Scholar]
- Clark, E. Dynamic Patterning by the Drosophila Pair-Rule Network Reconciles Long-Germ and Short-Germ Segmentation. PLoS Biol. 2017, 15, e2002439. [Google Scholar] [CrossRef] [Green Version]
- Akam, M. The Molecular Basis for Metameric Pattern in the Drosophila Embryo. Development 1987, 101, 1–22. [Google Scholar] [CrossRef]
- Eldon, E.; Kooyer, S.; D’Evelyn, D.; Duman, M.; Lawinger, P.; Botas, J.; Bellen, H. The Drosophila 18 Wheeler Is Required for Morphogenesis and Has Striking Similarities to Toll. Development 1994, 120, 885–899. [Google Scholar] [PubMed]
- Chiang, C.; Beachy, P.A. Expression of a Novel Toll-like Gene Spans the Parasegment Boundary and Contributes to Hedgehog Function in the Adult Eye of Drosophila. Mech. Dev. 1994, 47, 225–239. [Google Scholar]
- Kambris, Z.; Hoffmann, J.A.; Imler, J.L.; Capovilla, M. Tissue and Stage-Specific Expression of the Tolls in Drosophila Embryos. Gene Expr. Patterns 2002, 2, 311–317. [Google Scholar]
- Paré, A.C.; Naik, P.; Shi, J.; Mirman, Z.; Palmquist, K.H.; Zallen, J.A. An LRR Receptor-Teneurin System Directs Planar Polarity at Compartment Boundaries. Dev. Cell 2019, 51, 208–221.e6. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.; Price, B.D.; Bockheim, S.; Boedigheimer, M.J.; Smith, R.; Laughon, A. Molecular and Genetic Characterization of the Drosophila Tartan Gene. Dev. Biol. 1993, 160, 315–332. [Google Scholar] [CrossRef] [PubMed]
- Graham, P.L.; Anderson, W.R.; Brandt, E.A.; Xiang, J.; Pick, L. Dynamic Expression of Drosophila Segmental Cell Surface-Encoding Genes and Their Pair-Rule Regulators. Dev. Biol. 2019, 447, 147–156. [Google Scholar]
- Tetley, R.J.; Blanchard, G.B.; Fletcher, A.G.; Adams, R.J.; Sanson, B. Unipolar Distributions of Junctional Myosin II Identify Cell Stripe Boundaries That Drive Cell Intercalation throughout Drosophila Axis Extension. eLife 2016, 5, e12094. [Google Scholar] [CrossRef]
- Sharrock, T.E.; Evans, J.; Blanchard, G.B.; Sanson, B. Different Temporal Requirements for Tartan and Wingless in the Formation of Contractile Interfaces at Compartmental Boundaries. Development 2022, 149, dev200292. [Google Scholar] [CrossRef] [PubMed]
- Farrell, D.L.; Weitz, O.; Magnasco, M.O.; Zallen, J.A. SEGGA: A Toolset for Rapid Automated Analysis of Epithelial Cell Polarity and Dynamics. Development 2017, 144, 1725–1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Gonzalez, R.; de Matos Simões, S.; Röper, J.C.; Eaton, S.; Zallen, J.A. Myosin II Dynamics Are Regulated by Tension in Intercalating Cells. Dev. Cell 2009, 17, 736–743. [Google Scholar] [PubMed] [Green Version]
- Yu, J.C.; Fernandez-Gonzalez, R. Local Mechanical Forces Promote Polarized Junctional Assembly and Axis Elongation in Drosophila. eLife 2016, 5, e10757. [Google Scholar] [CrossRef]
- Bella, J.; Hindle, K.L.; McEwan, P.A.; Lovell, S.C. The Leucine-Rich Repeat Structure. Cell. Mol. Life Sci. 2008, 65, 2307–2333. [Google Scholar]
- Botos, I.; Segal, D.M.; Davies, D.R. The Structural Biology of Toll-Like Receptors. Structure 2011, 19, 447. [Google Scholar]
- Parthier, C.; Stelter, M.; Ursel, C.; Fandrich, U.; Lilie, H.; Breithaupt, C.; Stubbs, M.T. Structure of the Toll-Spatzle Complex, a Molecular Hub in Drosophila Development and Innate Immunity. Proc. Natl. Acad. Sci. USA 2014, 111, 6281–6286. [Google Scholar]
- Özkan, E.; Carrillo, R.A.; Eastman, C.L.; Weiszmann, R.; Waghray, D.; Johnson, K.G.; Zinn, K.; Celniker, S.E.; Garcia, K.C. An Extracellular Interactome of Immunoglobulin and LRR Proteins Reveals Receptor-Ligand Networks. Cell 2013, 154, 228–239. [Google Scholar]
- Lavalou, J.; Mao, Q.; Harmansa, S.; Kerridge, S.; Lellouch, A.C.; Philippe, J.M.; Audebert, S.; Camoin, L.; Lecuit, T. Formation of Polarized Contractile Interfaces by Self-Organized Toll-8/Cirl GPCR Asymmetry. Dev. Cell 2021, 56, 1574–1588.e7. [Google Scholar]
- Iijima, N.; Sato, K.; Kuranaga, E.; Umetsu, D. Differential Cell Adhesion Implemented by Drosophila Toll Corrects Local Distortions of the Anterior-Posterior Compartment Boundary. Nat. Commun. 2020, 11, 6320. [Google Scholar]
- Leulier, F.; Lemaitre, B. Toll-like Receptors—Taking an Evolutionary Approach. Nat. Rev. Genet. 2008, 9, 165. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Akira, S. Signaling to NF-KappaB by Toll-like Receptors. Trends Mol. Med. 2007, 13, 460–469. [Google Scholar]
- Anderson, K.V.; Bokla, L.; Nüsslein-Volhard, C. Establishment of Dorsal-Ventral Polarity in the Drosophila Embryo: The Induction of Polarity by the Toll Gene Product. Cell 1985, 42, 791–798. [Google Scholar]
- Anderson, K.V.; Jürgens, G.; Nüsslein-Volhard, C. Establishment of Dorsal-Ventral Polarity in the Drosophila Embryo: Genetic Studies on the Role of the Toll Gene Product. Cell 1985, 42, 779–789. [Google Scholar] [PubMed]
- Nüsslein-Volhard, C. The Toll Gene in Drosophila Pattern Formation. Trends Genet. 2022, 38, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.V. Toll Signaling Pathways in the Innate Immune Response. Curr. Opin. Immunol. 2000, 12, 13–19. [Google Scholar] [PubMed]
- Kleve, C.D.; Siler, D.A.; Syed, S.K.; Eldon, E.D. Expression of 18-Wheeler in the Follicle Cell Epithelium Affects Cell Migration and Egg Morphology in Drosophila. Dev. Dyn. 2006, 235, 1953–1961. [Google Scholar]
- Kim, S.; Chung, S.; Yoon, J.; Choi, K.-W.; Yim, J. Ectopic Expression of Tollo/Toll-8 Antagonizes Dpp Signaling and Induces Cell Sorting in the Drosophila Wing. Genesis 2006, 44, 541–549. [Google Scholar]
- Kolesnikov, T.; Beckendorf, S.K. 18 Wheeler Regulates Apical Constriction of Salivary Gland Cells via the Rho-GTPase-Signaling Pathway. Dev. Biol. 2007, 307, 53–61. [Google Scholar]
- Yagi, Y.; Nishida, Y.; Ip, Y.T. Functional Analysis of Toll-Related Genes in Drosophila. Dev. Growth Differ. 2010, 52, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Benton, M.A.; Pechmann, M.; Frey, N.; Stappert, D.; Conrads, K.H.; Chen, Y.T.; Stamataki, E.; Pavlopoulos, A.; Roth, S. Toll Genes Have an Ancestral Role in Axis Elongation. Curr. Biol. 2016, 26, 1609–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, A.E.; Shamardani, K.; Lugo, K.A.; Deguine, J.; Roberts, A.W.; Lee, B.L.; Barton, G.M. A Map of Toll-like Receptor Expression in the Intestinal Epithelium Reveals Distinct Spatial, Cell Type-Specific, and Temporal Patterns. Immunity 2018, 49, 560–575.e6. [Google Scholar] [PubMed] [Green Version]
- McIlroy, G.; Foldi, I.; Aurikko, J.; Wentzell, J.S.; Lim, M.A.; Fenton, J.C.; Gay, N.J.; Hidalgo, A. Toll-6 and Toll-7 Function as Neurotrophin Receptors in the Drosophila Melanogaster CNS. Nat. Neurosci. 2013, 16, 1248–1256. [Google Scholar] [CrossRef] [Green Version]
- Ballard, S.L.; Miller, D.L.; Ganetzky, B. Retrograde Neurotrophin Signaling through Tollo Regulates Synaptic Growth In Drosophila. J. Cell Biol. 2014, 204, 1157–1172. [Google Scholar]
- Li, G.; Forero, M.G.; Wentzell, J.S.; Durmus, I.; Wolf, R.; Anthoney, N.C.; Parker, M.; Jiang, R.; Hasenauer, J.; Strausfeld, N.J.; et al. A Toll-Receptor Map Underlies Structural Brain Plasticity. eLife 2020, 9, e52743. [Google Scholar] [CrossRef]
- Kaul, D.; Habbel, P.; Derkow, K.; Krüger, C.; Franzoni, E.; Wulczyn, F.G.; Bereswill, S.; Nitsch, R.; Schott, E.; Veh, R.; et al. Expression of Toll-like Receptors in the Developing Brain. PLoS ONE 2012, 7, e37767. [Google Scholar] [CrossRef] [Green Version]
- Okun, E.; Griffioen, K.J.; Mattson, M.P. Toll-like Receptor Signaling in Neural Plasticity and Disease. Trends Neurosci. 2011, 34, 269–281. [Google Scholar]
- Carvalho, L.; Jacinto, A.; Matova, N. The Toll/NF-KappaB Signaling Pathway Is Required for Epidermal Wound Repair in Drosophila. Proc. Natl. Acad. Sci. USA 2014, 111, E5373–E5382. [Google Scholar]
- Capilla, A.; Karachentsev, D.; Patterson, R.A.; Hermann, A.; Juarez, M.T.; McGinnis, W. Toll Pathway Is Required for Wound-Induced Expression of Barrier Repair Genes in the Drosophila Epidermis. Proc. Natl. Acad. Sci. USA 2017, 114, E2682–E2688. [Google Scholar] [CrossRef]
- Meyer, S.N.; Amoyel, M.; Bergantinos, C.; de la Cova, C.; Schertel, C.; Basler, K.; Johnston, L.A. An Ancient Defense System Eliminates Unfit Cells from Developing Tissues during Cell Competition. Science 2014, 346, 1258236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamada, M.; Shi, J.; Bourdot, K.S.; Supriyatno, S.; Palmquist, K.H.; Gutierrez-Ruiz, O.L.; Zallen, J.A. Toll Receptors Remodel Epithelia by Directing Planar-Polarized Src and PI3K Activity. Dev. Cell 2021, 56, 1589–1602.e9. [Google Scholar] [CrossRef] [PubMed]
- Wymann, M.P.; Pirola, L. Structure and Function of Phosphoinositide 3-Kinases. Biochim. Biophys. Acta 1998, 1436, 127–150. [Google Scholar] [CrossRef] [PubMed]
- Ridley, A.J. Rho Family Proteins: Coordinating Cell Responses. Trends Cell Biol. 2001, 11, 471–477. [Google Scholar]
- Campa, C.C.; Ciraolo, E.; Ghigo, A.; Germena, G.; Hirsch, E. Crossroads of PI3K and Rac Pathways. Small GTPases 2015, 6, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Kerridge, S.; Munjal, A.; Philippe, J.M.; Jha, A.; De Las Bayonas, A.G.; Saurin, A.J.; Lecuit, T. Modular Activation of Rho1 by GPCR Signalling Imparts Polarized Myosin II Activation during Morphogenesis. Nat. Cell Biol. 2016, 18, 261–270. [Google Scholar] [CrossRef]
- Dahmann, C.; Oates, A.C.; Brand, M. Boundary Formation and Maintenance in Tissue Development. Nat. Rev. Genet. 2011, 12, 43–55. [Google Scholar] [CrossRef]
- Monier, B.; Pélissier-Monier, A.; Sanson, B. Establishment and Maintenance of Compartmental Boundaries: Role of Contractile Actomyosin Barriers. Cell. Mol. Life Sci. 2011, 68, 1897–1910. [Google Scholar]
- Pujades, C. The Multiple Functions of Hindbrain Boundary Cells: Tinkering Boundaries? Semin. Cell Dev. Biol. 2020, 107, 179–189. [Google Scholar]
- Garcia-Bellido, A.; Ripoll, P.; Morata, G. Developmental Compartmentalisation of the Wing Disk of Drosophila. Nat. New Biol. 1973, 245, 251–253. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, P.A. A Clonal Analysis of Segment Development in Oncopeltus (Hemiptera). J. Embryol. Exp. Morphol. 1973, 30, 681–699. [Google Scholar] [CrossRef]
- Crick, F.H.; Lawrence, P.A. Compartments and Polyclones in Insect Development. Science 1975, 189, 340–347. [Google Scholar] [CrossRef]
- Fraser, S.; Keynes, R.; Lumsden, A. Segmentation in the Chick Embryo Hindbrain Is Defined by Cell Lineage Restrictions. Nature 1990, 344, 431–435. [Google Scholar] [CrossRef]
- Zeltser, L.M.; Larsen, C.W.; Lumsden, A. A New Developmental Compartment in the Forebrain Regulated by Lunatic Fringe. Nat. Neurosci. 2001, 4, 683–684. [Google Scholar] [CrossRef]
- Zervas, M.; Millet, S.; Ahn, S.; Joyner, A.L. Cell Behaviors and Genetic Lineages of the Mesencephalon and Rhombomere 1. Neuron 2004, 43, 345–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langenberg, T.; Brand, M. Lineage Restriction Maintains a Stable Organizer Cell Population at the Zebrafish Midbrain-Hindbrain Boundary. Development 2005, 132, 3209–3216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez-Guri, E.; Udina, F.; Colas, J.-F.; Sharpe, J.; Padrón-Barthe, L.; Torres, M.; Pujades, C. Clonal Analysis in Mice Underlines the Importance of Rhombomeric Boundaries in Cell Movement Restriction during Hindbrain Segmentation. PLoS ONE 2010, 5, e10112. [Google Scholar] [CrossRef]
- Kulesa, P.M.; Fraser, S.E. Cell Dynamics during Somite Boundary Formation Revealed by Time-Lapse Analysis. Science 2002, 298, 991–995. [Google Scholar] [CrossRef] [Green Version]
- Altabef, M.; Clarke, J.D.; Tickle, C. Dorso-Ventral Ectodermal Compartments and Origin of Apical Ectodermal Ridge in Developing Chick Limb. Development 1997, 124, 4547–4556. [Google Scholar] [CrossRef]
- Arques, C.G.; Doohan, R.; Sharpe, J.; Torres, M. Cell Tracing Reveals a Dorsoventral Lineage Restriction Plane in the Mouse Limb Bud Mesenchyme. Development 2007, 134, 3713–3722. [Google Scholar]
- Pearse, R.V., 2nd; Scherz, P.J.; Campbell, J.K.; Tabin, C.J. A Cellular Lineage Analysis of the Chick Limb Bud. Dev. Biol. 2007, 310, 388–400. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Q.; Chen, H.; Johnson, R.L. Lmx1b-Expressing Cells in the Mouse Limb Bud Define a Dorsal Mesenchymal Lineage Compartment. Genesis 2009, 47, 224–233. [Google Scholar] [CrossRef]
- Smith, D.M.; Tabin, C.J. Clonally Related Cells Are Restricted to Organ Boundaries Early in the Development of the Chicken Gut to Form Compartment Boundaries. Dev. Biol. 2000, 227, 422–431. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Pomares, J.M.; Foty, R.A. Tissue Fusion and Cell Sorting in Embryonic Development and Disease: Biomedical Implications. Bioessays 2006, 28, 809–821. [Google Scholar]
- Batlle, E.; Wilkinson, D.G. Molecular Mechanisms of Cell Segregation and Boundary Formation in Development and Tumorigenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008227. [Google Scholar] [CrossRef] [Green Version]
- Twigg, S.R.F.; Kan, R.; Babbs, C.; Bochukova, E.G.; Robertson, S.P.; Wall, S.A.; Morriss-Kay, G.M.; Wilkie, A.O.M. Mutations of Ephrin-B1 (EFNB1), a Marker of Tissue Boundary Formation, Cause Craniofrontonasal Syndrome. Proc. Natl. Acad. Sci. USA 2004, 101, 8652. [Google Scholar] [CrossRef]
- Scarpa, E.; Finet, C.; Blanchard, G.B.; Sanson, B. Actomyosin-Driven Tension at Compartmental Boundaries Orients Cell Division Independently of Cell Geometry In Vivo. Dev. Cell 2018, 47, 727–740.e6. [Google Scholar] [CrossRef] [Green Version]
- del Álamo, D.; Rouault, H.; Schweisguth, F. Mechanism and Significance of Cis-Inhibition in Notch Signalling. Curr. Biol. 2011, 21, R40–R47. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.H.; Axelrod, J.D.; Simon, M.A. Regulation of Frizzled by Fat-like Cadherins during Planar Polarity Signaling in the Drosophila Compound Eye. Cell 2002, 108, 675–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, Y.; Kerr, M.; Freeman, M. Modulation of Drosophila Retinal Epithelial Integrity by the Adhesion Proteins Capricious and Tartan. PLoS ONE 2008, 3, e1827. [Google Scholar] [CrossRef] [Green Version]
- Milán, M.; Pérez, L.; Cohen, S.M. Boundary Formation in the Drosophila Wing: Functional Dissection of Capricious and Tartan. Dev. Dyn. 2005, 233, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, K.T.; Kojima, T.; Aigaki, T.; Hayashi, S. Differential Control of Cell Affinity Required for Progression and Refinement of Cell Boundary during Drosophila Leg Segmentation. Dev. Biol. 2007, 309, 126–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krause, C.; Wolf, C.; Hemphälä, J.; Samakovlis, C.; Schuh, R. Distinct Functions of the Leucine-Rich Repeat Transmembrane Proteins Capricious and Tartan in the Drosophila Tracheal Morphogenesis. Dev. Biol. 2006, 296, 253–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucker, R.P. Teneurins: Domain Architecture, Evolutionary Origins, and Patterns of Expression. Front. Neurosci. 2018, 12, 938. [Google Scholar] [CrossRef] [Green Version]
- Cheung, A.; Schachermayer, G.; Biehler, A.; Wallis, A.; Missaire, M.; Hindges, R. Teneurin Paralogues Are Able to Localise Synaptic Sites Driven by the Intracellular Domain and Have the Potential to Form Cis-Heterodimers. Front. Neurosci. 2022, 16, 915149. [Google Scholar] [CrossRef]
- Hong, W.; Mosca, T.J.; Luo, L. Teneurins Instruct Synaptic Partner Matching in an Olfactory Map. Nature 2012, 484, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Mosca, T.J.; Hong, W.; Dani, V.S.; Favaloro, V.; Luo, L. Trans-Synaptic Teneurin Signalling in Neuromuscular Synapse Organization and Target Choice. Nature 2012, 484, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Mosca, T.J. On the Teneurin Track: A New Synaptic Organization Molecule Emerges. Front. Cell. Neurosci. 2015, 9, 204. [Google Scholar]
- Baumgartner, S.; Wides, R. Discovery of Teneurins. Front. Neurosci. 2019, 13, 230. [Google Scholar] [CrossRef] [Green Version]
- Leamey, C.A.; Merlin, S.; Lattouf, P.; Sawatari, A.; Zhou, X.; Demel, N.; Glendining, K.A.; Oohashi, T.; Sur, M.; Fässler, R. Ten_m3 Regulates Eye-Specific Patterning in the Mammalian Visual Pathway and Is Required for Binocular Vision. PLoS Biol. 2007, 5, e241. [Google Scholar] [CrossRef]
- Dharmaratne, N.; Glendining, K.A.; Young, T.R.; Tran, H.; Sawatari, A.; Leamey, C.A. Ten-M3 Is Required for the Development of Topography in the Ipsilateral Retinocollicular Pathway. PLoS ONE 2012, 7, e43083. [Google Scholar] [CrossRef]
- Antinucci, P.; Nikolaou, N.; Meyer, M.P.; Hindges, R. Teneurin-3 Specifies Morphological and Functional Connectivity of Retinal Ganglion Cells in the Vertebrate Visual System. Cell Rep. 2013, 5, 582–592. [Google Scholar] [CrossRef] [Green Version]
- Berns, D.S.; DeNardo, L.A.; Pederick, D.T.; Luo, L. Teneurin-3 Controls Topographic Circuit Assembly in the Hippocampus. Nature 2018, 554, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Nunes, S.M.; Ferralli, J.; Choi, K.; Brown-Luedi, M.; Minet, A.D.; Chiquet-Ehrismann, R. The Intracellular Domain of Teneurin-1 Interacts with MBD1 and CAP/Ponsin Resulting in Subcellular Codistribution and Translocation to the Nuclear Matrix. Exp. Cell Res. 2005, 305, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Keith, F.J.; Gay, N.J. The Drosophila Membrane Receptor Toll Can Function to Promote Cellular Adhesion. EMBO J. 1990, 9, 4299–4306. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.S.; Kim, S.E.; Heo, J.Y.; Lee, M.E.; Kim, H.M.; Paik, S.-G.; Lee, H.; Lee, J.-O. Crystal Structure of the TLR1-TLR2 Heterodimer Induced by Binding of a Tri-Acylated Lipopeptide. Cell 2007, 130, 1071–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.Y.; Nan, X.; Jin, M.S.; Youn, S.-J.; Ryu, Y.H.; Mah, S.; Han, S.H.; Lee, H.; Paik, S.-G.; Lee, J.-O. Recognition of Lipopeptide Patterns by Toll-like Receptor 2-Toll-like Receptor 6 Heterodimer. Immunity 2009, 31, 873–884. [Google Scholar] [CrossRef] [Green Version]
- Bugga, L.; Ratnaparkhi, A.; Zinn, K. The Cell Surface Receptor Tartan Is a Potential in Vivo Substrate for the Receptor Tyrosine Phosphatase Ptp52F. Mol. Cell. Biol. 2009, 29, 3390–3400. [Google Scholar]
- Zarember, K.A.; Godowski, P.J. Tissue Expression of Human Toll-like Receptors and Differential Regulation of Toll-like Receptor MRNAs in Leukocytes in Response to Microbes, Their Products, and Cytokines. J. Immunol. 2002, 168, 554–561. [Google Scholar] [CrossRef]
- Huhta, H.; Helminen, O.; Kauppila, J.H.; Salo, T.; Porvari, K.; Saarnio, J.; Lehenkari, P.P.; Karttunen, T.J. The Expression of Toll-like Receptors in Normal Human and Murine Gastrointestinal Organs and the Effect of Microbiome and Cancer. J. Histochem. Cytochem. 2016, 64, 470–482. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuebler, C.A.; Paré, A.C. Striped Expression of Leucine-Rich Repeat Proteins Coordinates Cell Intercalation and Compartment Boundary Formation in the Early Drosophila Embryo. Symmetry 2023, 15, 1490. https://doi.org/10.3390/sym15081490
Kuebler CA, Paré AC. Striped Expression of Leucine-Rich Repeat Proteins Coordinates Cell Intercalation and Compartment Boundary Formation in the Early Drosophila Embryo. Symmetry. 2023; 15(8):1490. https://doi.org/10.3390/sym15081490
Chicago/Turabian StyleKuebler, Chloe A., and Adam C. Paré. 2023. "Striped Expression of Leucine-Rich Repeat Proteins Coordinates Cell Intercalation and Compartment Boundary Formation in the Early Drosophila Embryo" Symmetry 15, no. 8: 1490. https://doi.org/10.3390/sym15081490