
Why Are Most Humans Right-Handed? The Modified Fighting Hypothesis

Abstract
:1. Explaining Handedness
2. The Fighting Hypothesis of Handedness
2.1. Does Left-Handedness Have Health Costs?
2.1.1. Neurodevelopmental, Cognitive, and Psychiatric Disorders
2.1.2. Is Left-Handedness Associated with Other General Health Problems?
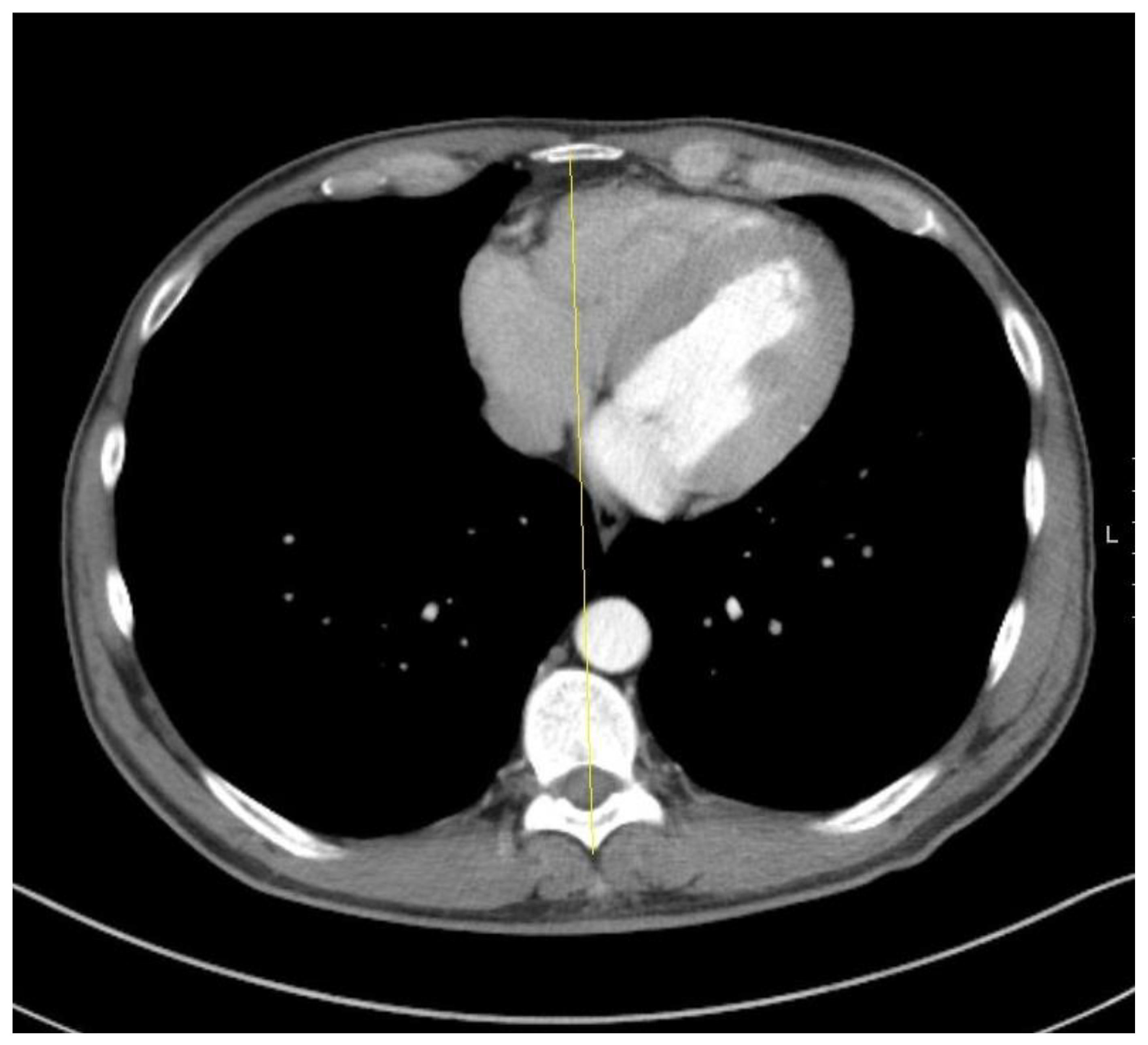
3. The Modified Fighting Hypothesis
3.1. Why Should Right-Handers Have a Fighting Advantage?
3.2. Could Weapon Use Have Caused Population-Level Handedness?
4. The Cause of Population-Level Lateralization May Facilitate the Co-Ordination of Other Laterality Phenotypes in Individuals
5. Novel Predictions
Differences between the Original Fighting Hypothesis and Modified Fighting Hypotheses: Contrasting Predictions
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Niven, J.E.; Frasnelli, E. Insights into the evolution of lateralization from the insects. Prog. Brain Res. 2018, 238, 3–31. [Google Scholar] [PubMed]
- Rogers, L.J. A matter of degree: Strength of brain asymmetry and behaviour. Symmetry 2017, 9, 57. [Google Scholar] [CrossRef]
- Vallortigara, G.; Rogers, L. Survival with an asymmetrical brain: Advantages and disadvantages of cerebral lateralization. Behav. Brain Sci. 2005, 28, 575–633. [Google Scholar] [CrossRef]
- Vallortigara, G.; Rogers, L.J. A function for the bicameral mind. Cortex 2020, 124, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Levy, J. The mammalian brain and the adaptive advantage of cerebral asymmetry. Ann. N. Y. Acad. Sci. 1977, 299, 264–272. [Google Scholar] [CrossRef]
- Rogers, L.J.; Vallortigara, G.; Andrew, R.J. Divided Brains: The Biology and Behaviour of Brain Asymmetries; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Gotts, S.J.; Jo, H.J.; Wallace, G.L.; Saad, Z.S.; Cox, B.C.; Martin, A. Two distinct forms of functional lateralization in the human brain. Proc. Natl. Acad. Sci. USA 2013, 110, E3435–E3444. [Google Scholar] [CrossRef]
- Güntürkün, O.; Ströckens, F.; Ocklenburg, S. Brain lateralization: A comparative perspective. Physiol. Rev. 2020, 100, 1019–1063. [Google Scholar] [CrossRef]
- Rogers, L.J. Manual bias, behavior, and cognition in common marmosets and other primates. Prog. Brain Res. 2018, 238, 91–113. [Google Scholar] [CrossRef]
- Raymond, M.; Pontier, D.; Dufour, A.-B.; Moller, A.P. Frequency-dependent maintenance of left handedness in humans. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1996, 263, 1627–1633. [Google Scholar] [CrossRef]
- Wiper, M.L. Evolutionary and mechanistic drivers of laterality: A review and new synthesis. Laterality Asymmetries Body Brain Cogn. 2017, 22, 740–770. [Google Scholar] [CrossRef]
- Corballis, M.C. From mouth to hand: Gesture, speech, and the evolution of right-handedness. Behav. Brain Sci. 2003, 26, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Corballis, M.C. Lateralization of the human brain. Prog. Brain Res. 2012, 195, 103–121. [Google Scholar] [CrossRef]
- Corballis, M.C. What’s left in language? Beyond the classical model. Ann. N. Y. Acad. Sci. 2015, 1359, 14–29. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Russell, J.; Freeman, H.; Buehler, N.; Reynolds, E.; Schapiro, S.J. The distribution and development of handedness for manual gestures in captive chimpanzees (Pan troglodytes). Psychol. Sci. 2005, 16, 487–493. [Google Scholar] [CrossRef]
- Vauclair, J. Lateralization of communicative signals in nonhuman primates and the hypothesis of the gestural origin of language. Interact. Stud. 2004, 5, 365–386. [Google Scholar] [CrossRef]
- Bakan, P.; Dibb, G.; Reed, P. Handedness and birth stress. Neuropsychologia 1973, 11, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Geschwind, N.; Behan, P. Left-handedness: Association with immune disease, migraine, and developmental learning disorder. Proc. Natl. Acad. Sci. USA 1982, 79, 5097–5100. [Google Scholar] [CrossRef]
- Llaurens, V.; Raymond, M.; Faurie, C. Why are some people left-handed? An evolutionary perspective. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 881–894. [Google Scholar] [CrossRef]
- Faurie, C.; Raymond, M. Handedness, homicide and negative frequency-dependent selection. Proc. R Soc. B Biol. Sci. 2005, 272, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Faurie, C.; Raymond, M. The fighting hypothesis as an evolutionary explanation for the handedness polymorphism in humans: Where are we? Ann. N. Y. Acad. Sci. 2013, 1288, 110–113. [Google Scholar] [CrossRef]
- Groothuis, T.G.; McManus, I.; Schaafsma, S.M.; Geuze, R.H. The fighting hypothesis in combat: How well does the fighting hypothesis explain human left-handed minorities? Ann. N. Y. Acad. Sci. 2013, 1288, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Coren, S.; Porac, C. Family patterns in four dimensions of lateral preference. Behav. Genet. 1980, 10, 333–348. [Google Scholar] [CrossRef]
- Schmitz, J.; Zheng, M.; Lui, K.F.H.; McBride, C.; Ho, C.S.-H.; Paracchini, S. Quantitative multidimensional phenotypes improve genetic analysis of laterality traits. Transl. Psychiatry 2022, 12, 68. [Google Scholar] [CrossRef]
- Papadatou-Pastou, M.; Martin, M.; Munafò, M.R.; Jones, G.V. Sex differences in left-handedness: A meta-analysis of 144 studies. Psychol. Bull. 2008, 134, 677. [Google Scholar] [CrossRef]
- Peters, M.; Reimers, S.; Manning, J.T. Hand preference for writing and associations with selected demographic and behavioral variables in 255,100 subjects: The BBC internet study. Brain Cogn. 2006, 62, 177–189. [Google Scholar] [CrossRef]
- Medland, S.E.; Perelle, I.; De Monte, V.; Ehrman, L. Effects of culture, sex, and age on the distribution of handedness: An evaluation of the sensitivity of three measures of handedness. Laterality Asymmetries Body Brain Cogn. 2004, 9, 287–297. [Google Scholar] [CrossRef]
- De Kovel, C.G.; Carrión-Castillo, A.; Francks, C. A large-scale population study of early life factors influencing left-handedness. Sci. Rep. 2019, 9, 584. [Google Scholar] [CrossRef]
- Papadatou-Pastou, M.; Ntolka, E.; Schmitz, J.; Martin, M.; Munafò, M.R.; Ocklenburg, S.; Paracchini, S. Human handedness: A meta-analysis. Psychol. Bull. 2020, 146, 481. [Google Scholar] [CrossRef] [PubMed]
- Fox, C.L.; Frayer, D.W. Non-dietary marks in the anterior dentition of the Krapina Neanderthals. Int. J. Osteoarchaeol. 1997, 7, 133–149. [Google Scholar] [CrossRef]
- Steele, J.; Uomini, N. Humans, tools and Handedness. In Stone Knapping: The Necessary Conditions for a Uniquely Hominin Behaviour; Roux, V., Bril, B., Eds.; MacDonald Institute for Archaeological Research: Cambridge, UK, 2005; pp. 217–239. [Google Scholar]
- Toth, N. Archaeological evidence for preferential right-handedness in the Lower and Middle Pleistocene, and its possible implications. J. Hum. Evol. 1985, 14, 607–614. [Google Scholar] [CrossRef]
- Uomini, N.T. Handedness in Neanderthals. In Neanderthal Lifeways, Subsistence and Technology: One Hundred Fifty Years of Neanderthal Study; Conard, N.J., Richter, J., Eds.; Vertebrate Paleobiology and Paleoanthropology Series; Springer: Dordrecht, The Netherlands, 2011; pp. 139–154. [Google Scholar] [CrossRef]
- Hepper, P.G. The developmental origins of laterality: Fetal handedness. Dev. Psychobiol. 2013, 55, 588–595. [Google Scholar] [CrossRef]
- Hepper, P.G.; Shahidullah, S.; White, R. Handedness in the human fetus. Neuropsychologia 1991, 29, 1107–1111. [Google Scholar] [CrossRef]
- Paracchini, S. Recent advances in handedness genetics. Symmetry 2021, 13, 1792. [Google Scholar] [CrossRef]
- Brandler, W.M.; Morris, A.P.; Evans, D.M.; Scerri, T.S.; Kemp, J.P.; Timpson, N.J.; Pourcain, B.S.; Smith, G.D.; Ring, S.M.; Stein, J.; et al. Common variants in left/right asymmetry genes and pathways are associated with relative hand skill. PLoS Genet. 2013, 9, e1003751. [Google Scholar] [CrossRef]
- McManus, I.C.; Davison, A.; Armour, J.A. Multilocus genetic models of handedness closely resemble single-locus models in explaining family data and are compatible with genome-wide association studies. Ann. N. Y. Acad. Sci. 2013, 1288, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Borod, J.C.; Caron, H.S.; Koff, E. Left-handers and right-handers compared on performance and preference measures of lateral dominance. Br. J. Psychol. 1984, 75, 177–186. [Google Scholar] [CrossRef]
- McManus, I.; Van Horn, J.D.; Bryden, P.J. The Tapley and Bryden test of performance differences between the hands: The original data, newer data, and the relation to pegboard and other tasks. Laterality Asymmetries Body Brain Cogn. 2016, 21, 371–396. [Google Scholar] [CrossRef]
- McManus, I.C. Eye-dominance, writing hand, and throwing hand. Laterality Asymmetries Body Brain Cogn. 1999, 4, 173–192. [Google Scholar] [CrossRef]
- Johnstone, L.T.; Karlsson, E.M.; Carey, D.P. Left-handers are less lateralized than right-handers for both left and right hemispheric functions. Cereb. Cortex 2021, 31, 3780–3787. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, E.M. Cerebral Asymmetries: Handedness and the Right Hemisphere; Bangor University: Bangor, UK, 2019. [Google Scholar]
- Rodway, P.; Thoma, V.; Schepman, A. The effects of sex and handedness on masturbation laterality and other lateralized motor behaviours. Laterality 2022, 27, 324–352. [Google Scholar] [CrossRef] [PubMed]
- Vingerhoets, G.; Acke, F.; Alderweireldt, A.-S.; Nys, J.; Vandemaele, P.; Achten, E. Cerebral lateralization of praxis in right-and left-handedness: Same pattern, different strength. Hum. Brain Mapp. 2012, 33, 763–777. [Google Scholar] [CrossRef]
- Vingerhoets, G. Phenotypes in hemispheric functional segregation? Perspectives and challenges. Phys. Life Rev. 2019, 30, 1–18. [Google Scholar] [CrossRef]
- Packheiser, J.; Schmitz, J.; Berretz, G.; Papadatou-Pastou, M.; Ocklenburg, S. Handedness and sex effects on lateral biases in human cradling: Three meta-analyses. Neurosci. Biobehav. Rev. 2019, 104, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Billiard, S.; Faurie, C.; Raymond, M. Maintenance of handedness polymorphism in humans: A frequency-dependent selection model. J. Theor. Biol. 2005, 235, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Frasnelli, E.; Vallortigara, G. Individual-level and population-level lateralization: Two sides of the same coin. Symmetry 2018, 10, 739. [Google Scholar] [CrossRef]
- Ghirlanda, S.; Vallortigara, G. The evolution of brain lateralization: A game-theoretical analysis of population structure. Proc. R Soc. London. Ser. B Biol. Sci. 2004, 271, 853–857. [Google Scholar] [CrossRef]
- Ghirlanda, S.; Frasnelli, E.; Vallortigara, G. Intraspecific competition and coordination in the evolution of lateralization. Philos. Trans. R Soc. B Biol. Sci. 2009, 364, 861–866. [Google Scholar] [CrossRef]
- Rogers, L.J. Laterality in animals. Int. J. Comp. Psychol. 1989, 3, 5–25. [Google Scholar] [CrossRef]
- Marzoli, D.; D’anselmo, A.; Malatesta, G.; Lucafò, C.; Prete, G.; Tommasi, L. The intricate web of asymmetric processing of social stimuli in humans. Symmetry 2022, 14, 1096. [Google Scholar] [CrossRef]
- Rogers, L.J.; Rigosi, E.; Frasnelli, E.; Vallortigara, G. A right antenna for social behaviour in honeybees. Sci. Rep. 2013, 3, 2045. [Google Scholar] [CrossRef]
- Stancher, G.; Sovrano, V.A.; Vallortigara, G. Motor asymmetries in fishes, amphibians, and reptiles. Prog. Brain Res. 2018, 238, 33–56. [Google Scholar] [CrossRef]
- Benelli, G.; Romano, D.; Stefanini, C.; Kavallieratos, N.G.; Athanassiou, C.G.; Canale, A. Asymmetry of mating behaviour affects copulation success in two stored-product beetles. J. Pest Sci. 2017, 90, 547–556. [Google Scholar] [CrossRef]
- Boukouvala, M.C.; Kavallieratos, N.G.; Canale, A.; Benelli, G. Functional Asymmetries Routing the Mating Behavior of the Rusty Grain Beetle, Cryptolestes ferrugineus (Stephens) (Coleoptera: Laemophloeidae). Insects 2022, 13, 699. [Google Scholar] [CrossRef]
- Vallortigara, G.; Bisazza, A. How ancient is brain lateralization. Comp. Vertebr. Lateralization 2002, 9, 69. [Google Scholar]
- Romano, D.; Canale, A.; Benelli, G. Do right-biased boxers do it better? Population-level asymmetry of aggressive displays enhances fighting success in blowflies. Behav. Process. 2015, 113, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Schnell, A.K.; Jozet-Alves, C.; Hall, K.C.; Radday, L.; Hanlon, R.T. Fighting and mating success in giant Australian cuttlefish is influenced by behavioural lateralization. Proc. R Soc. B 2019, 286, 20182507. [Google Scholar] [CrossRef] [PubMed]
- Karenina, K.; Giljov, A.; Ingram, J.; Rowntree, V.J.; Malashichev, Y. Lateralization of mother–infant interactions in a diverse range of mammal species. Nat. Ecol. Evol. 2017, 1, 0030. [Google Scholar] [CrossRef]
- Karenina, K.; Giljov, A. Mother and offspring lateralized social behavior across mammalian species. Prog. Brain Res. 2018, 238, 115–141. [Google Scholar] [CrossRef]
- Giljov, A.; Karenina, K. Positional biases in social behaviors: Humans vs. saiga antelopes. Front. Behav. Neurosci. 2022, 16, 1103584. [Google Scholar] [CrossRef]
- Tonello, L.; Vallortigara, G. Evolutionary models of lateralization: Steps toward stigmergy? Front. Behav. Neurosci. 2023, 17, 1121335. [Google Scholar] [CrossRef] [PubMed]
- Díaz, S.; Murray, L.; Roberts, S.G.; Rodway, P. Between-task consistency, temporal stability and the role of posture in simple reach and fishing hand preference in chimpanzees (Pan troglodytes). Appl. Anim. Behav. Sci. 2021, 242, 105417. [Google Scholar] [CrossRef]
- Ströckens, F.; Güntürkün, O.; Ocklenburg, S. Limb preferences in non-human vertebrates. Laterality Asymmetries Body Brain Cogn. 2013, 18, 536–575. [Google Scholar] [CrossRef] [PubMed]
- Soto, C.; Gázquez, J.M.; Llorente, M. Hand preferences in coordinated bimanual tasks in non-human primates: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2022, 141, 104822. [Google Scholar] [CrossRef]
- Chagnon, N.A. Life histories, blood revenge, and warfare in a tribal population. Science 1988, 239, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Puts, D.A. Beauty and the beast: Mechanisms of sexual selection in humans. Evol. Hum. Behav. 2010, 31, 157–175. [Google Scholar] [CrossRef]
- Kordsmeyer, T.L.; Hunt, J.; Puts, D.; Ostner, J.; Penke, L. The relative importance of intra-and intersexual selection on human male sexually dimorphic traits. Evol. Hum. Behav. 2018, 39, 424–436. [Google Scholar] [CrossRef]
- Richardson, T.; Gilman, R.T. Left-handedness is associated with greater fighting success in humans. Sci. Rep. 2019, 9, 15402. [Google Scholar] [CrossRef] [PubMed]
- Richardson, T. Is arm length a sexually selected trait in humans? Evidence from mixed martial arts. Evol. Behav. Sci. 2021, 15, 175. [Google Scholar] [CrossRef]
- Pollet, T.V.; Stulp, G.; Groothuis, T.G. Born to win? Testing the fighting hypothesis in realistic fights: Left-handedness in the Ultimate Fighting Championship. Anim. Behav. 2013, 86, 839–843. [Google Scholar] [CrossRef]
- Grouios, G.; Tsorbatzoudis, H.; Alexandris, K.; Barkoukis, V. Do left-handed competitors have an innate superiority in sports? Percept. Mot. Ski. 2000, 90 (Suppl. S3), 1273–1282. [Google Scholar] [CrossRef]
- Ziyagil, M.A.; Gursoy, R.; Dane, S.; Yuksel, R. Left-Handed Wrestlers Are More Successful. Percept. Mot. Ski. 2010, 111, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Loffing, F.; Hagemann, N.; Strauss, B. Left-handedness in professional and amateur tennis. PLoS ONE 2012, 7, e49325. [Google Scholar] [CrossRef] [PubMed]
- Witkowski, M.; Tomczak, M.; Karpowicz, K.; Solnik, S.; Przybyla, A. Effects of fencing training on motor performance and asymmetry vary with handedness. J. Mot. Behav. 2019, 52, 50–57. [Google Scholar] [CrossRef]
- Aggleton, J.P.; Wood, C.J. Is there a left-handed advantage in “ballistic” sports? Int. J. Sport Psychol. 1990, 21, 46–57. [Google Scholar]
- Loffing, F.; Hagemann, N. Pushing through evolution? Incidence and fight records of left-oriented fighters in professional boxing history. Laterality Asymmetries Body Brain Cogn. 2015, 20, 270–286. [Google Scholar] [CrossRef]
- Porac, C.; Coren, S. Lateral Preferences and human Behavior; Springer: Berlin/Heidelberg, Germany, 1981. [Google Scholar]
- Baker, J.; Kungl, A.-M.; Pabst, J.; Strauß, B.; Büsch, D.; Schorer, J. Your fate is in your hands? Handedness, digit ratio (2D: 4D), and selection to a national talent development system. Laterality Asymmetries Body Brain Cogn. 2013, 18, 710–718. [Google Scholar] [CrossRef]
- Loffing, F.; Schorer, J.; Hagemann, N.; Baker, J. On the advantage of being left-handed in volleyball: Further evidence of the specificity of skilled visual perception. Atten. Percept. Psychophys. 2012, 74, 446–453. [Google Scholar] [CrossRef]
- Hagemann, N. The advantage of being left-handed in interactive sports. Atten. Percept. Psychophys. 2009, 71, 1641–1648. [Google Scholar] [CrossRef]
- Akpinar, S.; Sainburg, R.L.; Kirazci, S.; Przybyla, A. Motor asymmetry in elite fencers. J. Mot. Behav. 2015, 47, 302–311. [Google Scholar] [CrossRef]
- Schaafsma, S.M.; Geuze, R.H.; Riedstra, B.; Schiefenhövel, W.; Bouma, A.; Groothuis, T.G. Handedness in a nonindustrial society challenges the fighting hypothesis as an evolutionary explanation for left-handedness. Evol. Hum. Behav. 2012, 33, 94–99. [Google Scholar] [CrossRef]
- Zickert, N.; Geuze, R.H.; van der Feen, F.E.; Groothuis, T.G. Fitness costs and benefits associated with hand preference in humans: A large internet study in a Dutch sample. Evol. Hum. Behav. 2018, 39, 235–248. [Google Scholar] [CrossRef]
- Porac, C. Laterality: Exploring the Enigma of Left-Handedness; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Backwell, P.; Matsumasa, M.; Double, M.; Roberts, A.; Murai, M.; Keogh, J.S.; Jennions, M.D. What are the consequences of being left-clawed in a predominantly right-clawed fiddler crab? Proc. R Soc. B Biol. Sci. 2007, 274, 2723–2729. [Google Scholar] [CrossRef] [PubMed]
- van der Feen, F.E.; Zickert, N.; Groothuis, T.G.G.; Geuze, R.H. Does hand skill asymmetry relate to creativity, developmental and health issues and aggression as markers of fitness? Laterality 2020, 25, 53–86. [Google Scholar] [CrossRef]
- Hamaoui, J. Development of Laterality: Asymmetrical Human Behaviors and Perceptual Biases; Université de Strasbourg: Strasbourg, France, 2022. [Google Scholar]
- McManus, I. Handedness and birth stress. Psychol. Med. 1981, 11, 485–496. [Google Scholar] [CrossRef]
- Bailey, L.; McKeever, W. A large-scale study of handedness and pregnancy/birth risk events: Implications for genetic theories of handedness. Laterality Asymmetries Body Brain Cogn. 2004, 9, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, J.; Stratford, R.; Bailey, B.; McCaughey, E.; Betts, P. Hormones and handedness. Horm. Res. Paediatr. 2001, 56, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, M.E.; Johnston, D.W.; Shields, M.A. Adverse birth factors predict cognitive ability, but not hand preference. Neuropsychology 2012, 26, 578. [Google Scholar] [CrossRef]
- Bishop, D.V. Handedness and Developmental Disorder; Cambridge University Press: Cambridge, UK, 1990; Volume 110. [Google Scholar]
- Richards, G.; Beking, T.; Kreukels, B.P.; Geuze, R.H.; Beaton, A.A.; Groothuis, T. An examination of the influence of prenatal sex hormones on handedness: Literature review and amniotic fluid data. Horm. Behav. 2021, 129, 104929. [Google Scholar] [CrossRef]
- Grimshaw, G.M.; Bryden, M.P.; Finegan, J.-A.K. Relations between prenatal testosterone and cerebral lateralization in children. Neuropsychology 1995, 9, 68. [Google Scholar] [CrossRef]
- Pfannkuche, K.A.; Bouma, A.; Groothuis, T.G. Does testosterone affect lateralization of brain and behaviour? A meta-analysis in humans and other animal species. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 929–942. [Google Scholar] [CrossRef]
- Richardson, T. No association between adult sex steroids and hand preference in humans. Am. J. Hum. Biol. 2022, 34, e23605. [Google Scholar] [CrossRef] [PubMed]
- Berretz, G.; Wolf, O.T.; Güntürkün, O.; Ocklenburg, S. Atypical lateralization in neurodevelopmental and psychiatric disorders: What is the role of stress? Cortex 2020, 125, 215–232. [Google Scholar] [CrossRef] [PubMed]
- Malatesta, G.; Marzoli, D.; Tommasi, L. Environmental and genetic determinants of sensorimotor asymmetries in mother-infant interaction. Front. Behav. Neurosci. 2022, 16, 1080141. [Google Scholar] [CrossRef]
- Abbondanza, F.; Dale, R.S.; Wang, C.; Hayiou-Thomas, M.E.; Toseeb, U.; Koomar, T.; Wigg, K.; Feng, Y.; Price, K.; Kerr, E.N.; et al. Non-right handedness is associated with language and reading impairments. Child Dev. 2023, 1–16. [Google Scholar]
- Papadatou-Pastou, M.; Tomprou, D.-M. Intelligence and handedness: Meta-analyses of studies on intellectually disabled, typically developing, and gifted individuals. Neurosci. Biobehav. Rev. 2015, 56, 151–165. [Google Scholar] [CrossRef]
- Rodriguez, A.; Kaakinen, M.; Moilanen, I.; Taanila, A.; McGough, J.J.; Loo, S.; Järvelin, M.-R. Mixed-handedness is linked to mental health problems in children and adolescents. Pediatrics 2010, 125, e340–e348. [Google Scholar] [CrossRef]
- Björk, T.; Brus, O.; Osika, W.; Montgomery, S. Laterality, hand control and scholastic performance: A British birth cohort study. BMJ Open 2012, 2, e000314. [Google Scholar] [CrossRef]
- Papadatou-Pastou, M. Handedness and cognitive ability: Using meta-analysis to make sense of the data. Prog. Brain Res. 2018, 238, 179–206. [Google Scholar] [CrossRef]
- Bryden, P.J.; Bruyn, J.; Fletcher, P. Handedness and health: An examination of the association between different handedness classifications and health disorders. Laterality Asymmetries Body Brain Cogn. 2005, 10, 429–440. [Google Scholar] [CrossRef]
- Satz, P. Pathological left-handedness: An explanaory model. Cortex 1972, 8, 121–135. [Google Scholar] [CrossRef]
- McManus, C. Half a century of handedness research: Myths, truths; fictions, facts; backwards, but mostly forwards. Brain Neurosci. Adv. 2019, 3, 2398212818820513. [Google Scholar] [CrossRef]
- Coren, S.; Halpern, D.F. Left-handedness: A marker for decreased survival fitness. Psychol. Bull. 1991, 109, 90. [Google Scholar] [CrossRef]
- Harris, L.J. Do left-handers die sooner than right-handers? Commentary on Coren and Halpern’s (1991) “Left-handedness: A marker for decreased survival fitness.” Psychol. Bull. 1993, 114, 203–234. [Google Scholar] [CrossRef]
- Harris, L.J. Reply to Halpern and coren. Psychol. Bull. 1993, 114, 242–247. [Google Scholar] [CrossRef]
- Cerhan, J.R.; Folsom, A.R.; Potter, J.D.; Prineas, R.J. Handedness and mortality risk in older women. Am. J. Epidemiol. 1994, 140, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Persson, P.-G.; Allebeck, P. Do left-handers have increased mortality? Epidemiology 1994, 5, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Steenhuis, R.E.; Østbye, T.; Walton, R. An examination of the hypothesis that left-handers die earlier: The Canadian study of health and aging. Laterality Asymmetries Body Brain Cogn. 2001, 6, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Fritschi, L.; Divitini, M.L.; Talbot-Smith, A.; Knuiman, M. Left-handedness and risk of breast cancer. Br. J. Cancer 2007, 97, 686–687. [Google Scholar] [CrossRef]
- Porac, C.; Searleman, A. The effects of hand preference side and hand preference switch history on measures of psychological and physical well-being and cognitive performance in a sample of older adult right-and left-handers. Neuropsychologia 2002, 40, 2074–2083. [Google Scholar] [CrossRef]
- Wysocki, C.; McManus, I. Left-handers have a lower prevalence of arthritis and ulcer. Laterality Asymmetries Body Brain Cogn. 2005, 10, 97–102. [Google Scholar] [CrossRef]
- Packheiser, J.; Schmitz, J.; Stein, C.C.; Pfeifer, L.S.; Berretz, G.; Papadatou-Pastou, M.; Peterburs, J.; Ocklenburg, S. Handedness and depression: A meta-analysis across 87 studies. J. Affect. Disord. 2021, 294, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Schaafsma, S.M.; Geuze, R.H.; Lust, J.M.; Schiefenhovel, W.; Groothuis, T.G.G. The relation between handedness indices and reproductive success in a non-industrial society. PLoS ONE 2013, 8, e63114. [Google Scholar] [CrossRef] [PubMed]
- Larsson, M. Did heart asymmetry play a role in the evolution of human handedness? J. Cult. Cogn. Sci. 2017, 1, 65–76. [Google Scholar] [CrossRef]
- Churchill, S.E. Weapon technology, prey size selection, and hunting methods in modern hunter-gatherers: Implications for hunting in the Palaeolithic and Mesolithic. Archeol. Pap. Am. Anthropol. Assoc. 1993, 4, 11–24. [Google Scholar] [CrossRef]
- Harris, L.J. Left-Handedness: Early Theories, Facts, and Fancies. In Neuropsychology of Left-Handedness; Elsevier: Amsterdam, The Netherlands, 1980; pp. 3–78. [Google Scholar]
- Harris, L.J. In fencing, what gives left-handers the edge? Views from the present and the distant past. Laterality 2010, 15, 15–55. [Google Scholar] [CrossRef]
- McManus, I.C. Right Hand, Left Hand: The Origins of Asymmetry in Brains, Bodies, Atoms, and Cultures; Harvard University Press: Cambridge, MA, USA, 2002. [Google Scholar]
- Elias, L.J. Side Effects: How Left-Brain Right-Brain Differences Shape Everyday Behaviour; Dundurn: Toronto, ON, Canada, 2022. [Google Scholar]
- Kang, N.; Hsee, L.; Rizoli, S.; Alison, P. Penetrating cardiac injury: Overcoming the limits set by Nature. Injury 2009, 40, 919–927. [Google Scholar] [CrossRef]
- Muñoz, J.H.M.; Dussan, O.; Ruiz, F.; Rubiano, A.M.; Puyana, J.C. Penetrating cardiac trauma in stab wounds: A study of diagnostic accuracy of the cardiac area. Ulus Travma Acil Cerrahi Derg 2020, 26, 693–698. [Google Scholar] [CrossRef]
- Sandrasagra, F. Management of penetrating stab wounds of the chest: An assessment of the indications for early operation. Thorax 1978, 33, 474–478. [Google Scholar] [CrossRef]
- McGrew, W.C.; Marchant, L.F. On which side of the apes? Ethological study of laterality of hand use. Great Ape Soc. 1996, 255–272. [Google Scholar] [CrossRef]
- Goodall, J. Tool-using and aimed throwing in a community of free-living chimpanzees. Nature 1964, 201, 1264–1266. [Google Scholar] [CrossRef]
- Marchant, L.F.; McGrew, W.C.; Eibl-Eibesfeldt, I. Is human handedness universal? Ethological analyses from three traditional cultures. Ethology 1995, 101, 239–258. [Google Scholar] [CrossRef]
- Forrester, G.S.; Quaresmini, C.; Leavens, D.A.; Mareschal, D.; Thomas, M.S. Human handedness: An inherited evolutionary trait. Behav. Brain Res. 2013, 237, 200–206. [Google Scholar] [CrossRef]
- Caspar, K.R.; Pallasdies, F.; Mader, L.; Sartorelli, H.; Begall, S. The evolution and biological correlates of hand preferences in anthropoid primates. Elife 2022, 11, e77875. [Google Scholar] [CrossRef]
- McPherron, S.P.; Alemseged, Z.; Marean, C.W.; Wynn, J.G.; Reed, D.; Geraads, D.; Bobe, R.; Béarat, H.A. Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika, Ethiopia. Nature 2010, 466, 857–860. [Google Scholar] [CrossRef] [PubMed]
- Rolian, C.; Carvalho, S. Tool use and manufacture in the last common ancestor of Pan and Homo. In Chimpanzees and Human Evolution; Harvard University Press: Cambridge, MA, USA, 2017. [Google Scholar] [CrossRef]
- Wilkins, J.; Schoville, B.J.; Brown, K.S.; Chazan, M. Evidence for early hafted hunting technology. Science 2012, 338, 942–946. [Google Scholar] [CrossRef]
- Van Lawick-Goodall, J. Tool-Using in Primates and Other Vertebrates. In Advances in the Study of Behavior; Elsevier: Amsterdam, The Netherlands, 1971; pp. 195–249. [Google Scholar]
- Roffman, I.; Savage-Rumbaugh, S.; Rubert-Pugh, E.; Stadler, A.; Ronen, A.; Nevo, E. Preparation and use of varied natural tools for extractive foraging by bonobos (Pan paniscus). Am. J. Phys. Anthropol. 2015, 158, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Leutenegger, W.; Kelly, J.T. Relationship of sexual dimorphism in canine size and body size to social, behavioral, and ecological correlates in anthropoid primates. Primates 1977, 18, 117–136. [Google Scholar] [CrossRef]
- Greenfield, L.O. Origin of the human canine: A new solution to an old enigma. Am. J. Phys. Anthropol. 1992, 35, 153–185. [Google Scholar] [CrossRef]
- Suwa, G.; Kono, R.T.; Simpson, S.W.; Asfaw, B.; Lovejoy, C.O.; White, T.D. Paleobiological implications of the Ardipithecus ramidus dentition. Science 2009, 326, 69–99. [Google Scholar] [CrossRef]
- Lawrence, J.; Kimbel, W.H. Morphological integration of the canine region within the hominine alveolar arch. J. Hum. Evol. 2021, 154, 102942. [Google Scholar] [CrossRef]
- Kingdon, J. Lowly Origin: Where, When, and Why Humans First Stood Up; Princeton University Press: Princeton, NJ, USA, 2004. [Google Scholar]
- Ben-Dor, M.; Sirtoli, R.; Barkai, R. The evolution of the human trophic level during the Pleistocene. Am. J. Phys. Anthropol. 2021, 175, 27–56. [Google Scholar] [CrossRef]
- Alexander, R.D. How Did Humans Evolve? Reflections on the Uniquely Unique Species; University of Michigan Museum of Zoology: Ann Arbor, MI, USA, 1990. [Google Scholar]
- Flinn, M.V.; Geary, D.C.; Ward, C.V. Ecological dominance, social competition, and coalitionary arms races: Why humans evolved extraordinary intelligence. Evol. Hum. Behav. 2005, 26, 10–46. [Google Scholar] [CrossRef]
- Saladié, P.; Huguet, R.; Rodríguez-Hidalgo, A.; Cáceres, I.; Esteban-Nadal, M.; Arsuaga, J.L.; de Castro, J.M.B.; Carbonell, E. Intergroup cannibalism in the European Early Pleistocene: The range expansion and imbalance of power hypotheses. J. Hum. Evol. 2012, 63, 682–695. [Google Scholar] [CrossRef] [PubMed]
- Fibiger, L.; Ahlström, T.; Meyer, C.; Smith, M. Conflict, violence, and warfare among early farmers in Northwestern Europe. Proc. Natl. Acad. Sci. USA 2023, 120, e2209481119. [Google Scholar] [CrossRef] [PubMed]
- Briffa, M.; Lane, S.M. The role of skill in animal contests: A neglected component of fighting ability. Proc. R. Soc. B Biol. Sci. 2017, 284, 20171596. [Google Scholar] [CrossRef]
- Palaoro, A.V.; Peixoto, P.E.C. The hidden links between animal weapons, fighting style, and their effect on contest success: A meta-analysis. Biol. Rev. 2022, 97, 1948–1966. [Google Scholar] [CrossRef]
- Loffing, F.; Hagemann, N. Performance Differences between Left-and Right-Sided Athletes in One-on-One Interactive Sports. In Laterality in Sports; Elsevier: Amsterdam, The Netherlands, 2016; pp. 249–277. [Google Scholar]
- Hayashi, M.; Saito, D.N.; Aramaki, Y.; Asai, T.; Fujibayashi, Y.; Sadato, N. Hemispheric asymmetry of frequency-dependent suppression in the ipsilateral primary motor cortex during finger movement: A functional magnetic resonance imaging study. Cereb. Cortex 2008, 18, 2932–2940. [Google Scholar] [CrossRef] [PubMed]
- Tzourio-Mazoyer, N.; Petit, L.; Zago, L.; Crivello, F.; Vinuesa, N.; Joliot, M.; Jobard, G.; Mellet, E.; Mazoyer, B. Between-hand difference in ipsilateral deactivation is associated with hand lateralization: fMRI mapping of 284 volunteers balanced for handedness. Front. Hum. Neurosci. 2015, 9, 5. [Google Scholar] [CrossRef]
- Peters, M. Subclassification of non-pathological left-handers poses problems for theories of handedness. Neuropsychologia 1990, 28, 279–289. [Google Scholar] [CrossRef]
- Peters, M. Prevalence and classification. Brain Asymmetry 1996, 183–214. [Google Scholar]
- de Schotten, M.T.; Dell’acqua, F.; Forkel, S.; Simmons, A.; Vergani, F.; Murphy, D.G.; Catani, M. A lateralized brain network for visuo-spatial attention. Nat. Preced. 2011, 14, 1245–1246. [Google Scholar] [CrossRef]
- McCourt, M.E.; Garlinghouse, M. Asymmetries of visuospatial attention are modulated by viewing distance and visual field elevation: Pseudoneglect in peripersonal and extrapersonal space. Cortex 2000, 36, 715–731. [Google Scholar] [CrossRef]
- Annett, M.; Annett, J. Individual differences in right and left reaction time. Br. J. Psychol. 1979, 70, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Goodin, D.S.; Aminoff, M.J.; Ortiz, T.A.; Chequer, R.S. Response times and handedness in simple reaction-time tasks. Exp. Brain Res. 1996, 109, 117–126. [Google Scholar] [CrossRef]
- Gilbert, A.N.; Wysocki, C.J. Hand preference and age in the United States. Neuropsychologia 1992, 30, 601–608. [Google Scholar] [CrossRef]
- Portal, J. Patterns of eye-hand dominance in baseball players. N. Engl. J. Med. 1988, 319, 655–656. [Google Scholar]
- Gorynia, I.; Egenter, D. Intermanual coordination in relation to handedness, familial sinistrality and lateral preferences. Cortex 2000, 36, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Judge, J.; Stirling, J. Fine motor skill performance in left-and right-handers: Evidence of an advantage for left-handers. Laterality Asymmetries Body Brain Cogn. 2003, 8, 297–306. [Google Scholar] [CrossRef]
- Badzakova-Trajkov, G.; Häberling, I.S.; Roberts, R.P.; Corballis, M. Cerebral asymmetries: Complementary and independent processes. PLoS ONE 2010, 5, e9682. [Google Scholar] [CrossRef] [PubMed]
- Badzakova-Trajkov, G.; Corballis, M.; Häberling, I. Complementarity or independence of hemispheric specializations? A brief review. Neuropsychologia 2016, 93, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Bryden, M. Choosing sides: The left and right of the normal brain. Can. Psychol. Psychol. Can. 1990, 31, 297. [Google Scholar] [CrossRef]
- Karlsson, E.M.; Johnstone, L.T.; Carey, D.P. Reciprocal or independent hemispheric specializations: Evidence from cerebral dominance for fluency, faces, and bodies in right-and left-handers. Psychol. Neurosci. 2022, 15, 89. [Google Scholar] [CrossRef]
- McManus, C. Cerebral polymorphisms for lateralisation: Modelling the genetic and phenotypic architectures of multiple functional modules. Symmetry 2022, 14, 814. [Google Scholar] [CrossRef]
- Parker, A.J.; Woodhead, Z.V.J.; Thompson, P.A.; Bishop, D.V.M. Assessing the reliability of an online behavioural laterality battery: A pre-registered study. Laterality 2021, 26, 359–397. [Google Scholar] [CrossRef]
- Corballis, M.C. Humanity and the left hemisphere: The story of half a brain. Laterality 2021, 26, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Knecht, S.; Dräger, B.; Deppe, M.; Bobe, L.; Lohmann, H.; Flöel, A.; Ringelstein, E.-B.; Henningsen, H. Handedness and hemispheric language dominance in healthy humans. Brain 2000, 123, 2512–2518. [Google Scholar] [CrossRef]
- Carey, D.P.; Johnstone, L.T. Quantifying cerebral asymmetries for language in dextrals and adextrals with random-effects meta analysis. Front. Psychol. 2014, 5, 1128. [Google Scholar] [CrossRef]
- Mazoyer, B.; Zago, L.; Jobard, G.; Crivello, F.; Joliot, M.; Perchey, G.; Mellet, E.; Petit, L.; Tzourio-Mazoyer, N. Gaussian mixture modeling of hemispheric lateralization for language in a large sample of healthy individuals balanced for handedness. PLoS ONE 2014, 9, e101165. [Google Scholar] [CrossRef]
- Hiraoka, K.; Igawa, K.; Kashiwagi, M.; Nakahara, C.; Oshima, Y.; Takakura, Y. The laterality of stop and go processes of the motor response in left-handed and right-handed individuals. Laterality Asymmetries Body Brain Cogn. 2018, 23, 51–66. [Google Scholar] [CrossRef]
- Abrams, R.A.; Davoli, C.C.; Du, F.; Knapp, W.H.; Paull, D. Altered vision near the hands. Cognition 2008, 107, 1035–1047. [Google Scholar] [CrossRef]
- Reed, C.L.; Betz, R.; Garza, J.P.; Roberts, R.J. Grab it! Biased attention in functional hand and tool space. Atten. Percept. Psychophys. 2010, 72, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Le Bigot, N.; Grosjean, M. Effects of handedness on visual sensitivity in perihand space. PLoS ONE 2012, 7, e43150. [Google Scholar] [CrossRef]
- Shioiri, S.; Sasada, T.; Nishikawa, R. Visual attention around a hand location localized by proprioceptive information. Cereb. Cortex Commun. 2022, 3, tgac005. [Google Scholar] [CrossRef]
- Buckingham, G.; Main, J.C.; Carey, D.P. Asymmetries in motor attention during a cued bimanual reaching task: Left and right handers compared. Cortex 2011, 47, 432–440. [Google Scholar] [CrossRef]
- Marzoli, D.; Prete, G.; Tommasi, L. Perceptual asymmetries and handedness: A neglected link? Front. Psychol. 2014, 5, 163. [Google Scholar] [CrossRef]
- Coren, S.; Porac, C. Fifty centuries of right-handedness: The historical record. Science 1977, 198, 631–632. [Google Scholar] [CrossRef] [PubMed]
- McManus, I.C.; Moore, J.; Freegard, M.; Rawles, R. Science in the Making: Right Hand, Left Hand. III: Estimating historical rates of left-handedness. Laterality 2010, 15, 186–208. [Google Scholar] [CrossRef] [PubMed]
- McManus, I.C.; Hartigan, A. Declining left-handedness in Victorian England seen in the films of Mitchell and Kenyon. Curr. Biol. 2007, 17, R793–R794. [Google Scholar] [CrossRef]
- Rodway, P.; Schepman, A. Who goes where in couples and pairs? Effects of sex and handedness on side preferences in human dyads. Laterality 2022, 27, 415–442. [Google Scholar] [CrossRef]
- Güneş, E.; Nalçaci, E. Directional preferences in turning behavior of girls and boys. Percept. Mot. Ski. 2006, 102, 352–357. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Original Fighting Hypothesis (Raymond et al. [10]) | Modified Fighting Hypothesis | |
|---|---|---|
| Primary explicandum | Why are some people left-handed? | Why are most people right-handed? |
| Assumption | Right-handedness is the default; left-handedness needs an explanation. | Right-handedness and left-handedness both need explanations. |
| Hypothesis | Left-handers persist because they have a negative frequency-dependent fighting advantage. They are in the minority because they have a health cost. Right-handers are in the majority because they do not have a health cost. | Early male hominins fought with sharp weapons. Right-handers are in the majority because they had a fighting advantage due to the position of the heart when fighting with sharp weapons. Left-handers persist because they have a negative frequency-dependent fighting advantage. |
| Prediction: Left-handers over time | Left-handedness has a health cost. Frequency of left-handers will decline over time due to this health cost (as the fighting advantage no longer applies). | Left-handedness has no health cost. Frequency of left-handedness will remain stable or increase over time (as fighting with sharp weapons no longer applies). |
| Prediction: Relationships between the lateralization of different functions | Specific predictions not originally made. | The lateralization of different functions, that are related to fighting (e.g., handedness, footedness, and attention) will correlate more strongly with each other than with functions unrelated to fighting (e.g., language). |
| Predictions: Genetic factors | All lateralization is polygenetic, but theory makes no specific predictions about genetic clustering for different functions. | Greater genetic overlap for fighting-related functions than for other functions. Greater genetic differences between left- and right-handers for fighting-related functions. Greater genetic similarities for functions unrelated to fighting (e.g., language). |
| Prediction: Aggression | Left-handed males are more aggressive than right-handed males. | Left-handed males and right-handed males do not show different levels of aggression. |
| Prediction: Behaviours related to fighting (e.g., positioning and turning biases) | Predictions not originally specified. Would predict stronger behavioural biases in males than females. Could predict stronger biases in left-handed males than right-handed males. | Predicts stronger biases in males than females. Predicts that the biases will be equally strong in male right-handers and male left-handers. |
| Prediction: Fighting advantage | Left-handers have a fighting advantage in hand-to-hand combat and when using weapons | Right-handers have a fighting advantage when using sharp weapons. Left-handers could have an advantage in hand-to-hand combat. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Larsson, M.; Schepman, A.; Rodway, P. Why Are Most Humans Right-Handed? The Modified Fighting Hypothesis. Symmetry 2023, 15, 940. https://doi.org/10.3390/sym15040940
Larsson M, Schepman A, Rodway P. Why Are Most Humans Right-Handed? The Modified Fighting Hypothesis. Symmetry. 2023; 15(4):940. https://doi.org/10.3390/sym15040940
Chicago/Turabian StyleLarsson, Matz, Astrid Schepman, and Paul Rodway. 2023. "Why Are Most Humans Right-Handed? The Modified Fighting Hypothesis" Symmetry 15, no. 4: 940. https://doi.org/10.3390/sym15040940