1. Introduction
Theoretical ecologists are focusing on the study of interactions between living organisms and their environments; this is because the study is important in the formation of an ecosystem. In the 1920s, Lotka and Volterra individually developed first-order ordinary differential equations to report on the interactions between two species. Since then, researchers have been interested in modeling and analyzing interactions between species; see [
1,
2] and references therein. Different types of mathematical models, including ordinary [
2], partial [
3], non-integer order [
4], and difference equation [
5,
6], have been developed in order to make use of different environmental factors in predator–prey models, such as Allee effects, prey refuges, stage structure, harvesting, toxic effects, and environmental fluctuations. On the other hand, the existence of chaos in dynamical systems is quite obvious due to the presence of nonlinear terms. Predicting the future evolution of chaotic systems remains a difficult task due to its sensitivity to the initial condition and system parameters. In order to narrow this gap, the problem of chaos control has become a hot topic among researchers in various fields, such as biomedical systems [
7], ecological models [
8], convection models [
9], etc. The chaos control approach leads a chaotic system to a limit cycle or an asymptotically stable state. In contrast, the problem of chaotification is becoming more popular because of its applications in the fields of secure communication, encryption, description, signal processing, etc., where chaotic behavior is a desired phenomenon; see [
10,
11,
12] for more details. In ecological systems, chaotic phenomena need to be controlled in order to predict how species will evolve in the future. The chaotic behaviors in the three-species food chain were initially noticed by Hastings and Powell [
13]. Since then, many academics have attempted to address the issue of how to use ecological factors to regulate chaos in food chain models (see [
14,
15,
16,
17,
18,
19,
20]). The authors of [
15] demonstrated that the food chain model exhibited chaos due to the parameters of prey growth rate and predator interference. Nath et al. [
17] showed that chaos can be controlled through prey refuge and the Allee effect in a food chain model. To regulate the system, the intermediate predator harvesting strategy was introduced in reference [
18]. The chaotic behavior of the prey–predator–parasite model can be controlled through a prey-harvesting strategy [
19]. Recently, Nitu and Vikas [
20] considered the tri-trophic food model, where the cannibalism effect was applied to middle predators to control the system dynamics.
From a biological perspective, interspecific killing among carnivores is a much more frequent issue. The interaction among species is symmetrical if both species kill each other, and the interaction is asymmetrical if one species kills another [
21]. On the other hand, the relationship between predator and prey is impacted not only by the method of direct encounters but also by an indirect effect (such as fear), which also alters the prey’s usual characteristics [
22,
23]. The altered characteristics could be associated with the prey’s nature, morphology, and habitat. To escape from the predation risk, the prey always attempts to change its usual habitat to a safer location [
24]. As a consequence, the short-term survivability of the prey is increasing; it has started to decrease in the long term. Numerous theoretical and experimental studies indicate that indirect effects can have notable impacts on the dynamics of predator–prey interactions [
25,
26,
27,
28,
29,
30,
31,
32,
33]. The growth rate of elk in the Greater Yellowstone Ecosystem is affected by their fear of wolves [
25], while mule deer shorten their feeding activity due to the risk of mountain lion predation. The authors of [
22] discovered that predator fear of song sparrows resulted in a 40% reduction in the number of offspring produced, even in the absence of direct contact with predators. Suraci et al. [
27] experimented for over a month on mesocarnivores (raccoons) by creating fear through the sounds of their predators and found that the fear of large carnivores reduced the raccoon’s foraging behavior by 66% and increased awareness. Based on the aforementioned ideas, Wang et al. [
23] mathematically modeled the impact of fear on the growth of the prey and observed that the cost of fear had a considerable impact on the dynamics of predator–prey interactions. Moreover, they noticed that fear can have a stabilizing effect on system dynamics. Panday et al. [
31] examined the dynamics of a food chain model with predation fear and they concluded that suitable fear effects can regularize the system from chaotic oscillation to stable. They also noticed that the top predator can lead to an extinction stage if the cost of fear of intermediate predators goes up. In [
29], Sasmal et al. dealt with the dynamics of the food chain model with the fear effect and group defense strategy among prey. In [
34], Mishra et al. studied the effect of fear in the agroecosystem. By utilizing the B-D functional response, Debnath et al. [
35] investigated the dynamics of a food chain model, and found that fear effects as well as mutual interference parameters among species can control the system dynamics.
The term “COE” often emerges from repetitive measures in clinical investigations. It has been recently applied to ecological and evolutionary concerns and applies to a wide range of circumstances. From an ecological point of view, COEs measure an individual species’ past learning, and can influence the current performance [
36]. The COE has a positive influence on the species community when past habitation is lost because some fatal or non-fatal effects are of poor quality, and it has a negative influence when the lost habitation is of high quality [
37]. COEs can occur over several seasons or even one season (i.e., changes in physiological behavior within a season). Experiments conducted in previous works [
38,
39,
40] show that amphibians, fish, marine insects, marine invertebrates, and other animals can experience carry-over effects in short periods of time and within a single season. Thus, emerging perspectives among ecologists show the importance of COEs in mathematical modeling; see [
37,
41,
42,
43]. Sasmal and Takeuchi [
42] incorporated predation fear and its COEs into models of prey–predator interactions and showed that fear and COE parameters can have significant impacts on system stability. Dubey and Sasmal [
43] considered a phytoplankton–zooplankton fish system, where zooplankton growth rates are influenced by fish-induced fear and the COEs. In addition, they pointed out that the system exhibited chaotic behavior for medium values of COE parameters, and the system became stable or periodic dynamics occurred for lower and higher values. Thus, incorporating COEs into an ecological model showed more insight into the factors that affected the species in the ecosystem.
Time delays in ecology are unavoidable because of some lags observed in ecological processes, such as maturation time, gestation, and handling time. Such time lags in ecological systems may produce more complex system dynamics. Thus, studying the dynamical features of ecological models with time lags has become an important topic among theoretical ecologists in the last decade, (see [
32,
44,
45,
46,
47,
48,
49] and references therein). To explore the dynamics of a food chain model, Pal et al. [
45] and Upadhyay et al. [
46], respectively, have taken the gestation delays of top predators only and both predators into account; they pointed out that delays significantly affected system stability. Recently, two different delays were established and analyzed in a tri-trophic food chain model by Surosh et al. in reference [
48]. To construct a more appropriate ecological model, it is important to include time lag in it.
Population ecologists are usually interested in forecasting the future population density of a species in an ecological system to maintain a healthy ecosystem. Most ecological systems are nonlinear, resulting in chaotic behaviors that need to be controlled in order to predict how the ecological systems will evolve in the future. In this connection, researchers have attempted to control chaos in ecological systems through various control methods. Controlling chaotic behavior in an ecological system must involve methods that are easy to use and have no negative influence on the real ecosystem. The artificial induction of fear among prey species by utilizing the sounds or vocal cues of the appropriate predators in real ecological systems is a control strategy that is crucial in preventing the extinction stage of interacting species. This fear lets the prey know to stay away from the predator and protects the prey species, as well as causes changes in its own characteristics. However, the prey learns about the fear induced and carries this information into future generations or upcoming generations, which makes it act again with its usual characteristics. As a result, induced fear among prey species fails to control the system’s chaotic dynamics. To overcome this situation, a non-chemical method, the concept of fear and fear-induced COE, is incorporated into ecological models. Moreover, there is still enough room to explore the dynamics of ecological models by introducing various ecological factors. To the best of our knowledge, there has been no work devoted to the three-species food chain model with fear-induced COEs in the growth terms of both prey and middle predators. This prompted our current investigation.
Inspired by the ideas of a fear-induced COE given in reference [
42,
43], we considered the chaos control of a tri-trophic food chain model in which the growth terms of prey and the middle predator were influenced by a fear-induced COE due to the predation risk. The relationships between the species followed the Holling type-II functional response. The developed model is an asymmetric food chain model since middle predators kill only prey and special predators kill only middle predators. To control the chaotic dynamics of the proposed system, a novel non-chemical method was imposed through varying fear and fear-induced COE parameters. The proposed method is based on the idea that vocal cues or sounds can be used to induce fear and that this process is carried over by prey species to the upcoming season or generation. Furthermore, we consider gestation delay in the growth terms of special predators. We derived local and global stability criteria and the Hopf bifurcation analysis of the integrated model. In addition, we incorporated gestation delay into the proposed and analyzed the local stability and bifurcation of the delayed model. The numerical results demonstrate the effectiveness of the novel method and confirm the theoretical results. The main contributions of this paper are as follows:
- i.
We propose a chaos control strategy based on inducing fear and its COEs in the prey and middle predator growth terms.
- ii.
We analyze how induced fear and its COE parameters affect the population densities of the species involved in the proposed model.
- iii.
Fear-induced COEs in the growth terms of both prey and middle predators are taken into account in this paper. This approach is different from other works in the literature.
- iv.
We determine the impact of the gestation delay of a special predator on the dynamics of the system.
The model considered in this paper generalizes the models investigated in reference [
13,
31,
45]. If the prey can learn about artificial vocal cues, then our theoretical findings clearly provide mechanisms to protect species in ecological systems. Moreover, the model suggested in this paper works better in places where there are three levels of food chains. It is also essential for developing optimized harvesting strategies in fisheries and pest management in agriculture.
The structure of the article is as follows. We provide a three-species food chain model with fear and its COEs in
Section 2.
Section 3 derives the preliminaries of a nonlinear food chain model. The chaos control of the tri-trophic food chain model is covered in
Section 4. In
Section 5, the food chain model is modified and examined with time delay. Selected numerical results are presented in
Section 6 to support our proposed theoretical findings. Finally, in
Section 7, we present the conclusions of the proposed study and suggest future research.
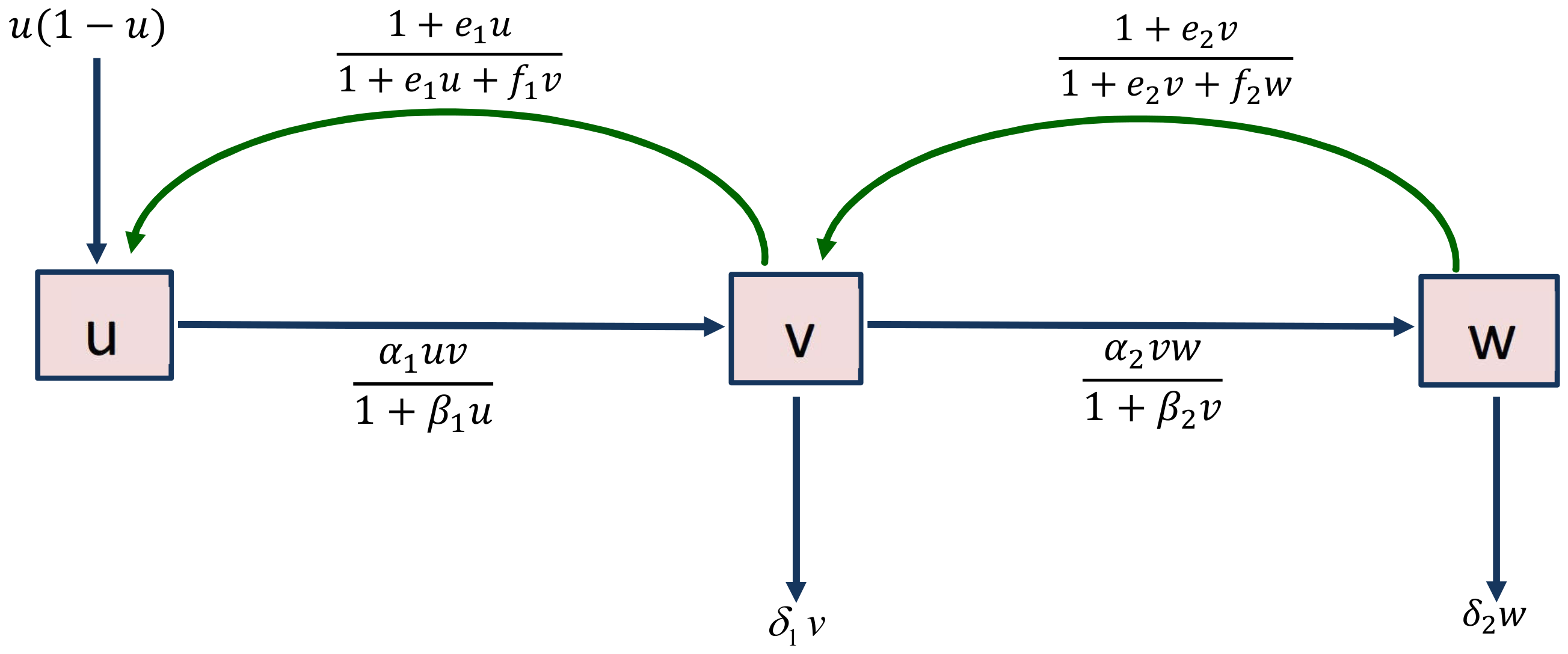
2. Description of the Tri-Trophic Food Chain Model
Mathematical models that describe the dynamics of a population can be categorized according to a continuous-time domain or a discrete-time domain. A continuous-time model is typically more appropriate if the species involved in the system have generations that overlap and births that are spaced throughout the year. Hastings–Powell [
13] initially discovered chaos control in a continuous food chain model, and since then, several authors have attempted to control chaos by introducing new strategies [
16,
31,
35]. In general, tritrophic food chains consist of prey, middle, and special predators at the bottom, middle, and top trophic levels. Hastings–Powell [
13] developed a food chain model in the form:
where
, and
are the respective densities of prey, middle, and special predators at time
T. The system parameters are all considered to be positive [
13].
and
K are the growth rates of the prey and environmental support capacity.
(or
) is the maximum attack rate of the middle (or special) predator.
(or
) represents the half-saturation coefficient of the prey (or middle predator).
(or
) indicates the conversion efficiencies of the middle (or special) predator.
(or
) is the death rate of the middle (or special) predator. To simplify the notation, the state variables
and
are denoted by
and
respectively. By introducing the effect of fear in model (
1), Panday et al. [
31] extended the model given (
1) into the following form
where
and
represent the intensity of fear in the prey and middle predator population, respectively. However, the prey carries over some information from previous predation attacks to subsequent generations. The carry-over information affects the growth rate of the species [
42,
43]. Taking into account the above viewpoints, we extend model (
2) by introducing fear-induced COEs. Then the system is given as follows:
where
and
are the carry-over effect parameters due to the fear
and
, respectively. In model (
3), we introduce the functions
and
, which represent fear and its COE in the interaction between the species. The functions
and
satisfy the following characteristics associated with fear and its COE [
42]:
and
and
and
and
and
Further, and satisfy the following characteristics associated with COE:
and , which indicates the positive effects of COE as a result of lessons learned from earlier seasons or experiences.
and , as the population densities of the prey and middle predator increase, respectively, increasing their growth rates.
and i.e., if the density of the prey’s population or COE becomes large, then there is no change in its growth rate.
and i.e., if the density of the middle predator’s population or COE becomes large, then there is no change in its growth rate.
If COE parameters and are zero, then the functions and only reveal the fear effect.
Based on the ideas given in reference [
13], to minimize the complexity of model (
3), we devise a non-dimensional scheme as follows:
,
,
,
and let
,
,
,
,
,
,
,
,
and
Then system (
3) becomes
subject to the initial conditions
. A conceptual diagram of model (
4) is represented in
Figure 1.
Remark 1. In this paper, we consider the delayed three-species food chain model with fear due to predation and its carry-over effect. If there is no fear-induced carry-over effect (i.e., ) in the proposed non-delayed model, then the model is reduced to the model studied by Pandey et al. in reference [31]. If there is no fear (i.e., ) in the proposed non-delayed model, then the model is reduced to the model dealt with by Hastings and Powell [13]. The proposed delayed model was the model discussed by the authors in reference [45] when there is no fear effect on predation growth. The results derived in this paper are general cases of the models investigated in reference [13,31,45]. Panday et al. [33] considered the problem of chaos control in a three-species food chain model by introducing fear to the middle predator only. The chaotic dynamics of the food chain model can be controlled through fear of predation and mutual interference among species [35]. It should be noted that artificially introduced fear plays a crucial role in controllin chaotic dynamics [13,33,35,45]. However, in [13,33,35,45], fear-induced COE is missing, which stimulated the present study. The model proposed in this paper is more applicable in places where there are three-level food chains. Moreover, the model is appropriate to food chain fisheries where the prey includes species, such as yellow perch, sunfish, and herring; the middle predators include species, such as walleye, tuna, and catfish; and the special predators are species, such as sharks, whales, and dolphins. Remark 2. The model proposed in this paper is based on the continuous-time model because the species involved in the system have generations that overlap and births that are spaced throughout the year. By using the standard finite difference schemes, one can obtain discrete-time models from the continuous ones, where the step size represents the generation time. In the absence of middle and special predators in model (4), the prey species grows logistically and the model takes the following formEquation (5) represents a single-species growth model (a logistic model). The discrete version of model (5) tends to be the logistic map, which exhibits a variety of complex behaviors, including unstable, stable, period-doubling, and chaotic oscillations. Chaotic dynamics can be seen in discrete-time models with only one species, but at least three species are needed for continuous-time systems. There has been a lot of research on the dynamical properties, including stability and bifurcation, of both continuous-time models [13,31,43] and discrete-time models [5,6] in independent ways. 6. Numerical Simulations
In this section, we conduct numerical simulations by using MATLAB and XPPAUT to explore various complex dynamics, including stable, unstable, periodic, and chaotic solutions of the derived model by varying the system parameters. The growth rates of the prey and middle predators will be influenced by the fear and COEs of their predators, which will result in complex dynamics of the food chain model. We should note that the model without the impact of COEs in system (
4) is considered in reference [
31]. In order to visualize the complex dynamics of the proposed model (
4), we present a bifurcation diagram, time trajectories, the largest Lyapunov exponent (LLE), and a phase portrait of the species. The algorithm for calculating LLE is given in
Appendix A. All of the parameters except fear and fear-induced COE are chosen from the reference [
13]. The ranges for the fear parameter and the fear-induced COE parameter are chosen from [
31,
43]. We consider the empirical values of the system parameters as follows.
With the parameters given in (
29), system (
4) having
and
The figures for system (
4) are drawn using the initial populations
. Among them, the only coexistence equilibrium point
is stable.
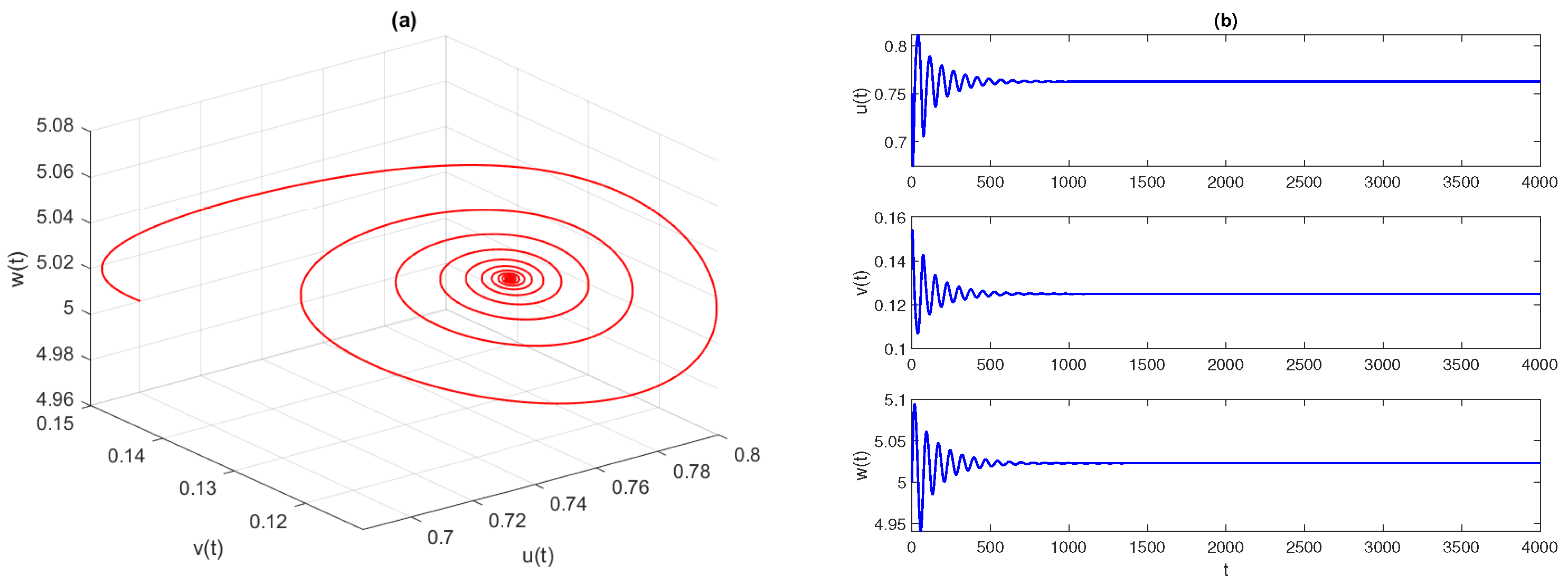
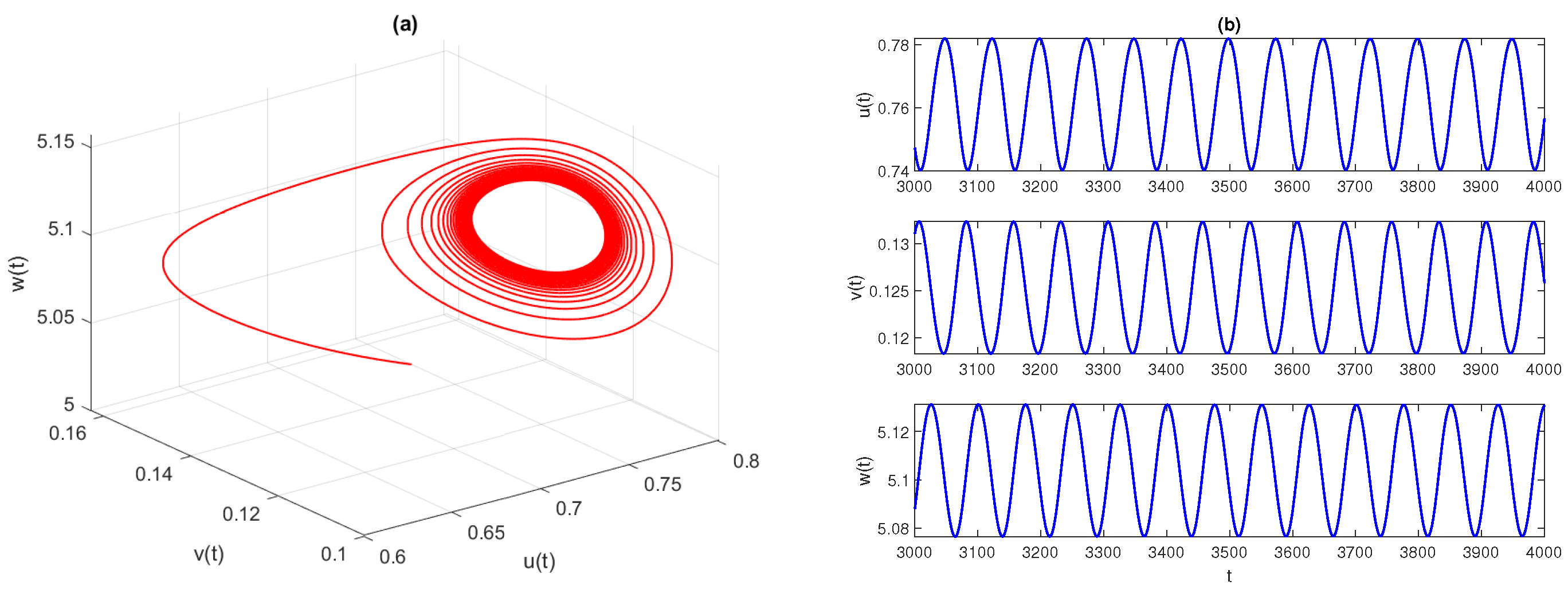
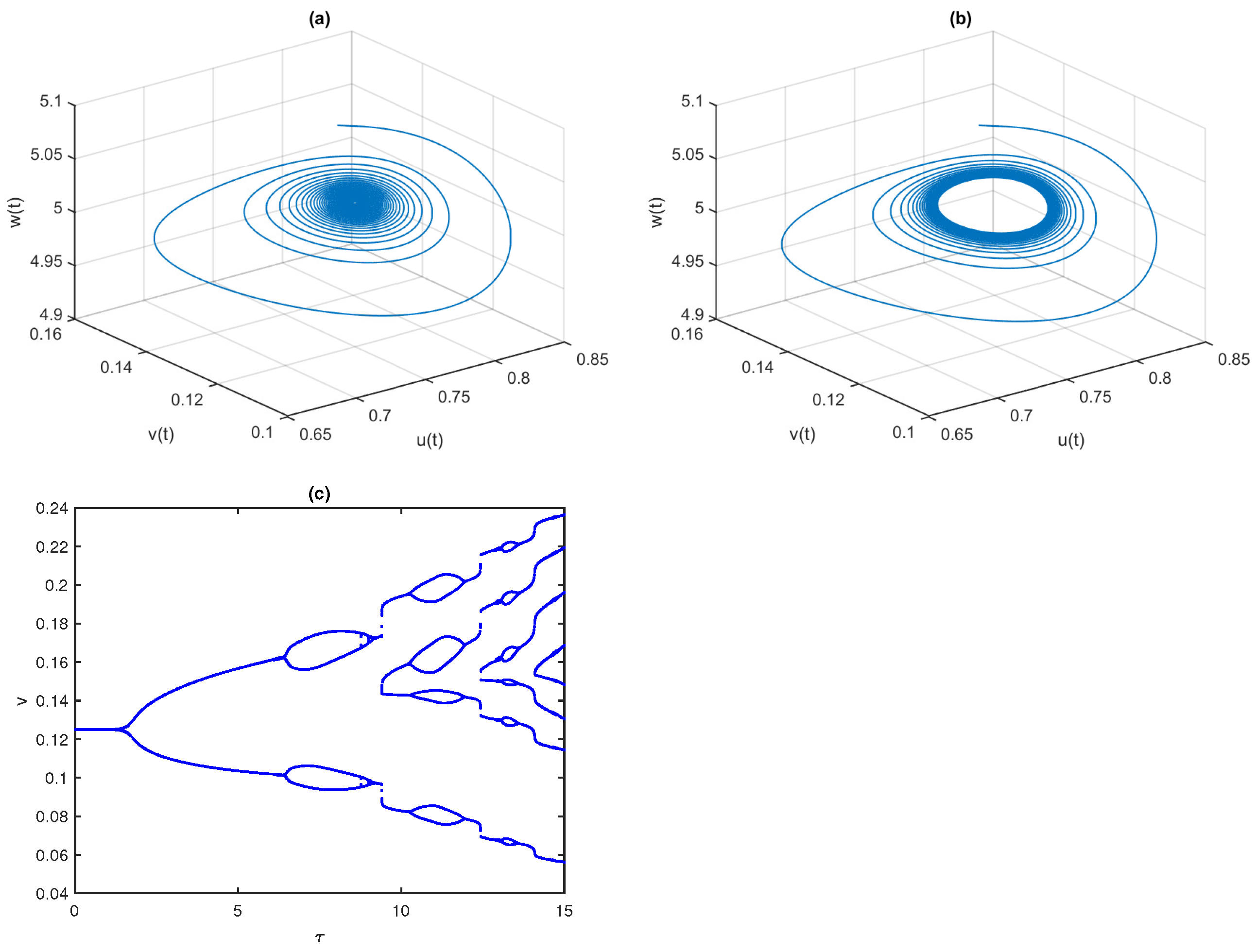
Figure 2 displays the phase space diagram and time-series solutions of model (
4). They show that solution trajectories converge asymptotically to the stable equilibrium point
Further, we observe that the equilibrium
is LAS for
. At the critical value of parameters
,
, the equilibrium point
loses its stability and the system experiences a Hopf bifurcation. The conditions given in Theorem 9 are well satisfied, as
. In
Figure 3, we draw the phase space diagram and time-series solutions of model (
4) with
and other parameters as given in Equation (
29). Furthermore, we examine the direction and stability properties of positive periodic solutions emerging from the coexistence equilibrium
. Applying the results obtained in
Section 4.4, we obtain that an occurrence of the Hopf bifurcation is subcritical and the corresponding bifurcating periodic solution is unstable.
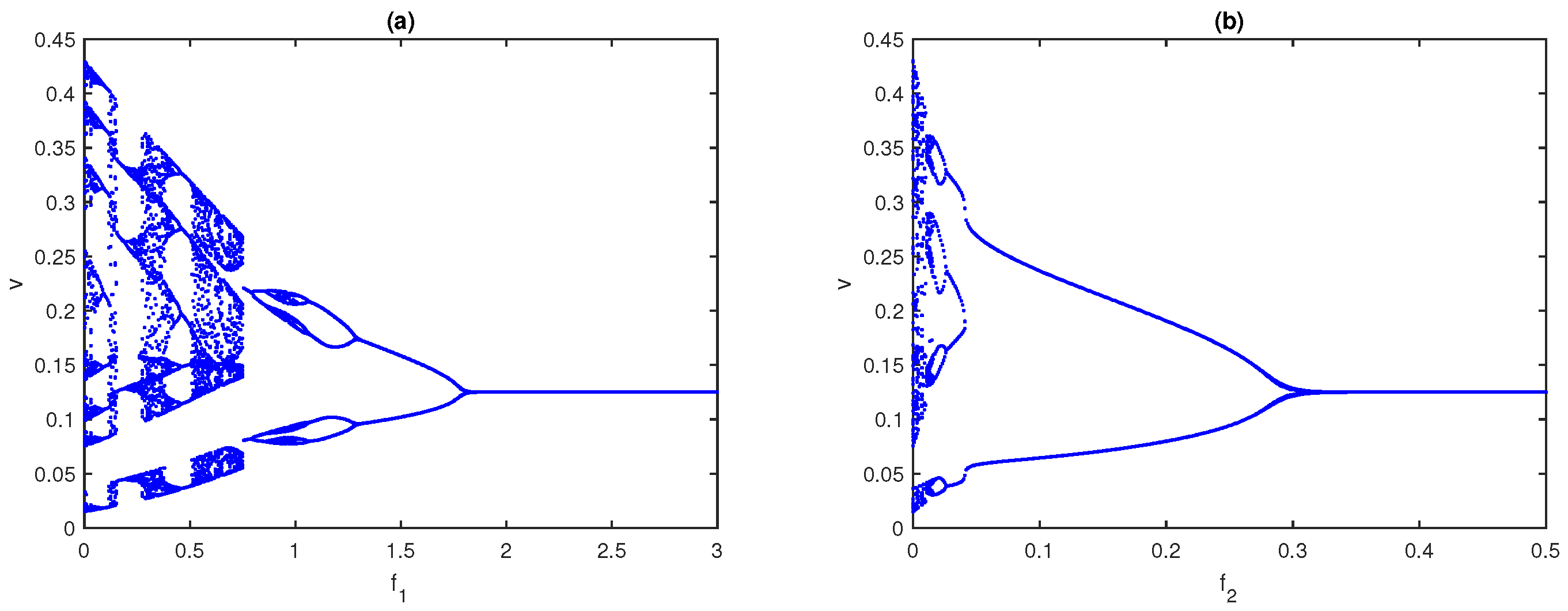
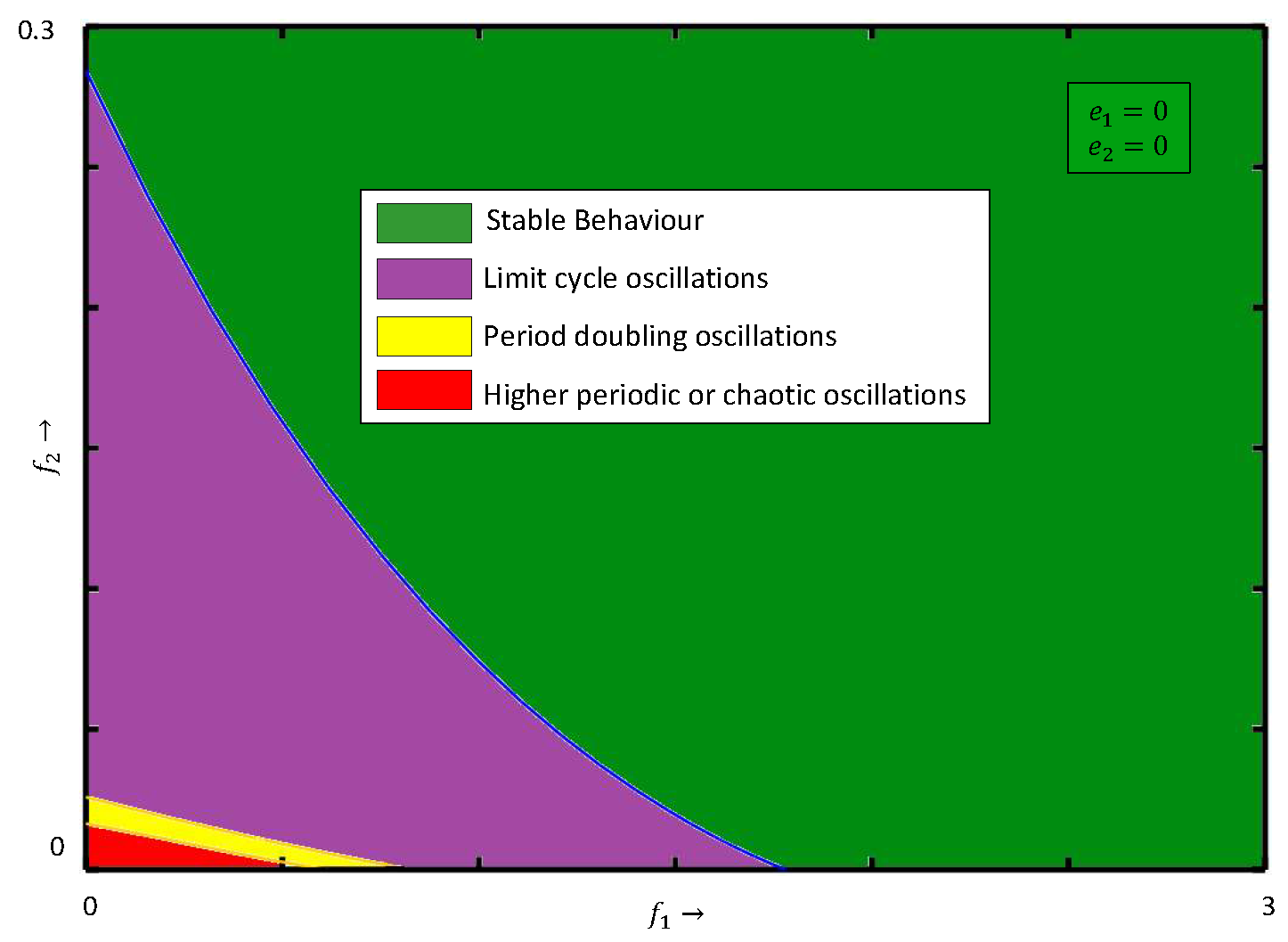
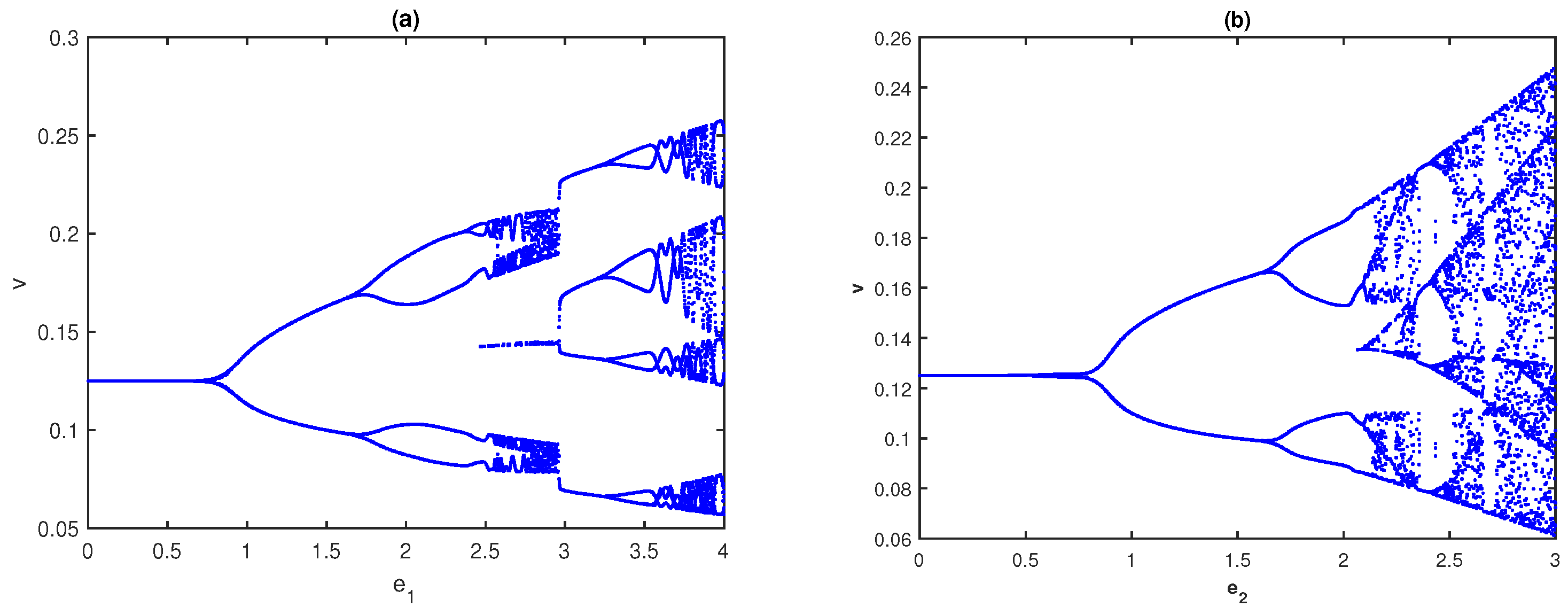
6.1. Impacts of Fear-Induced COEs
First, we explore the influence of fear in the absence of COEs. The bifurcation diagram of system (
4) with respect to
in the absence of
is presented in
Figure 4a, while the bifurcation diagram with respect to
in the absence of
is given in
Figure 4b. It is observed from
Figure 4a,b that the system, respectively, approaches a stable state from the chaotic nature through period-halving as fear parameters
and
increase. When both fear parameters are present, we plot a two-parameter bifurcation diagram in the
parametric plane; see
Figure 5. As we can see, for lower values of the
and
systems, (
4) exhibits chaotic behavior and then the system becomes stable for higher values. In an ecological sense, as fear parameters
and
increase, the growth rate of prey and middle predators decreases, resulting in less prey/middle predators being consumed by middle/special predators. As a result, all three species cannot become extinct in the system but they maintain the positive density. From the above discussions, fear parameters are extremely important in ecology because they may regulate the chaotic nature of the model and make it a stable one. However, the fear of predators also has some COE, which affects the growth rate of respective prey [
42].
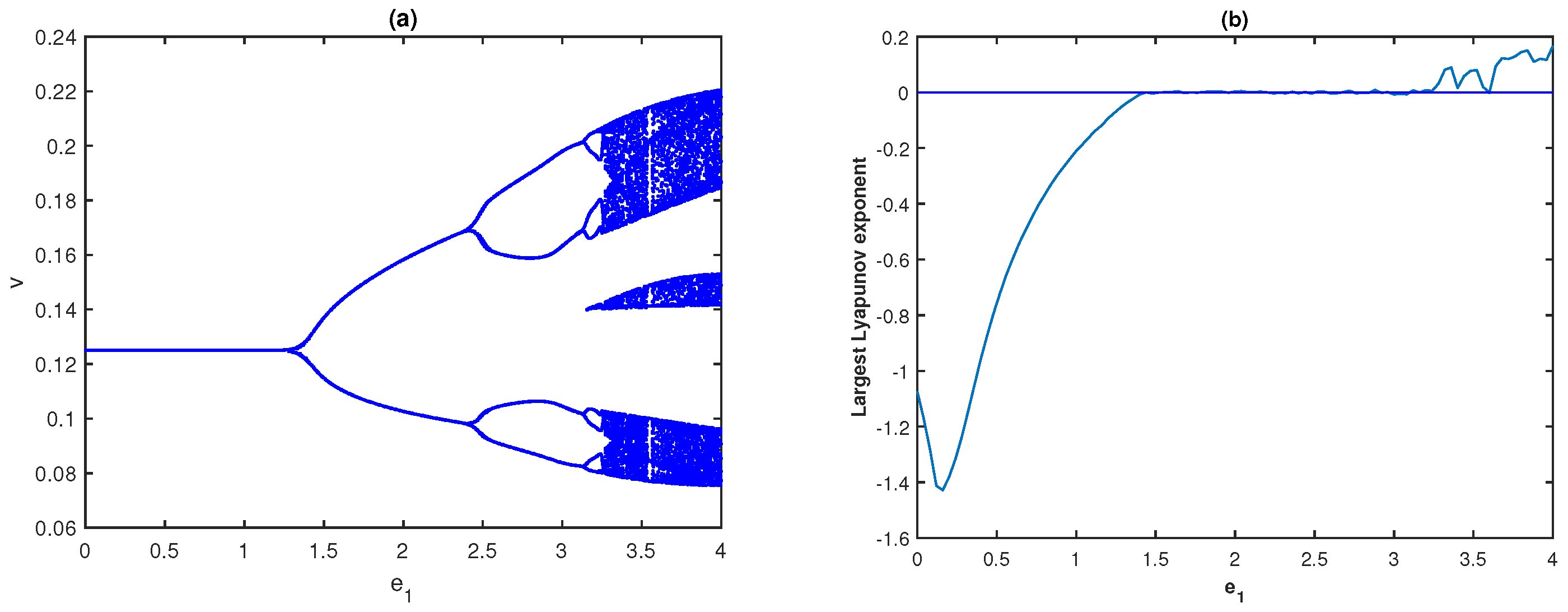
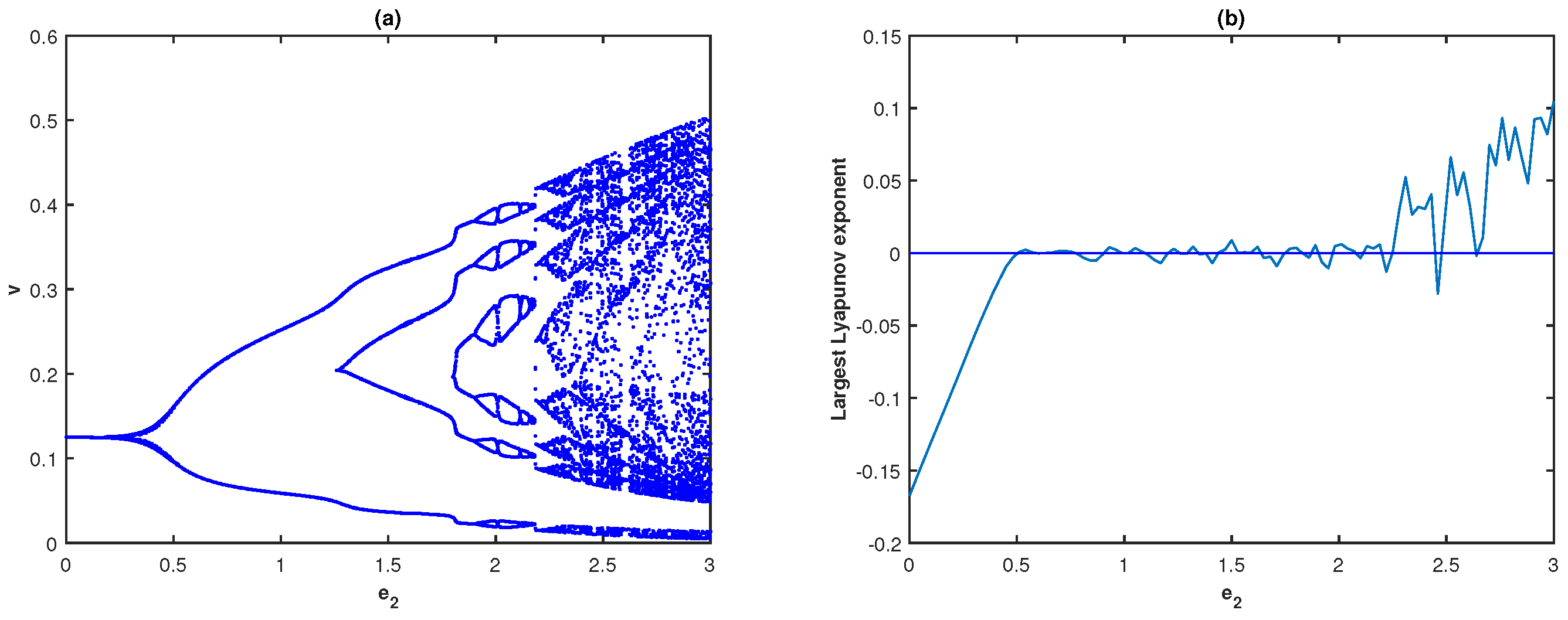
Next, we explore how fear-induced COE parameters are reflected in the system dynamics. First, we analyze the impact of the middle predator induced fear on the prey’s growth and its COE, while the fear of the special predator on the middle predator is absent
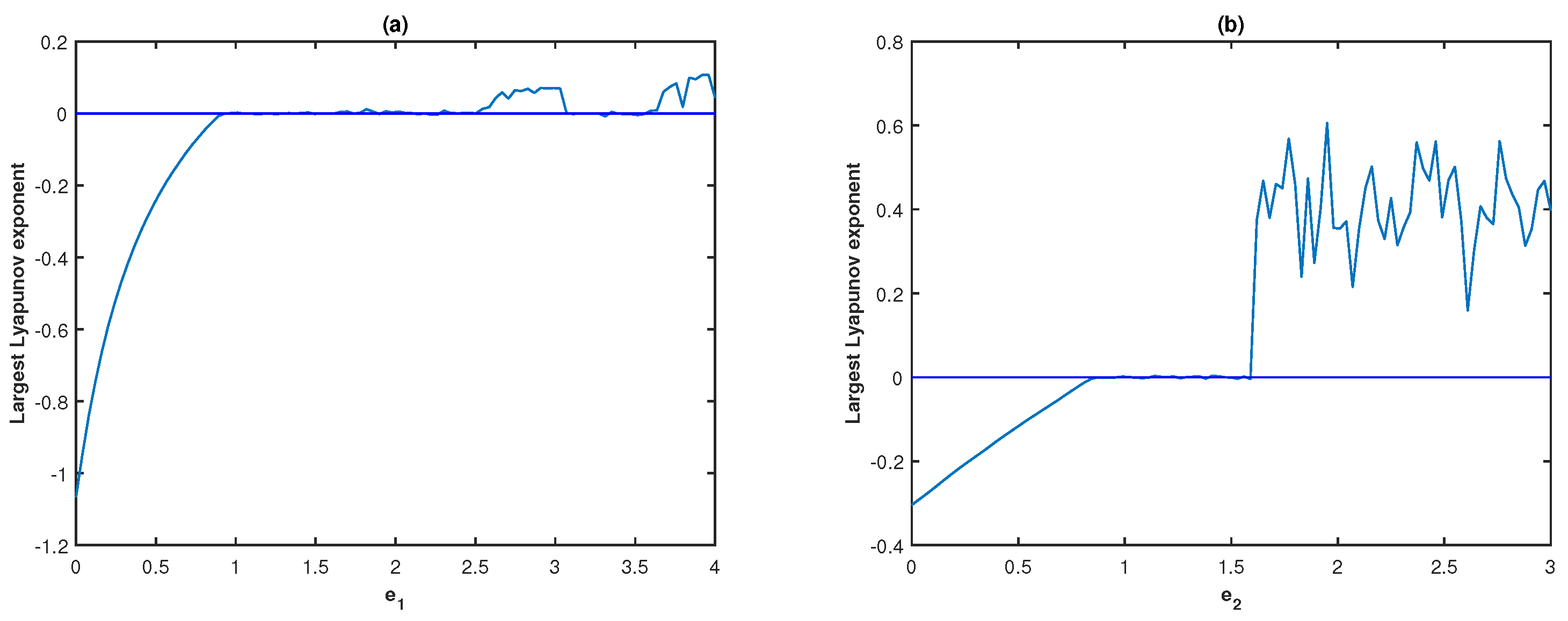
. The bifurcation diagram and fluctuations of LLE of system (
4) with respect to
when
are, respectively, presented in
Figure 6a,b. It can be seen in
Figure 6a that the system remains stable for the lower values of
, then the system turns chaotic through period-doubling as
increases.
Figure 6a illustrates that the system dynamics demonstrate stable behavior for
; limit cycle oscillations for
, and display higher periodic or chaotic oscillations for
In addition, we observe that the system remains chaotic for larger values of
To verify the chaotic behavior of system (
4), we draw the fluctuation of the LLE with respect to
in
Figure 6b. For different values of
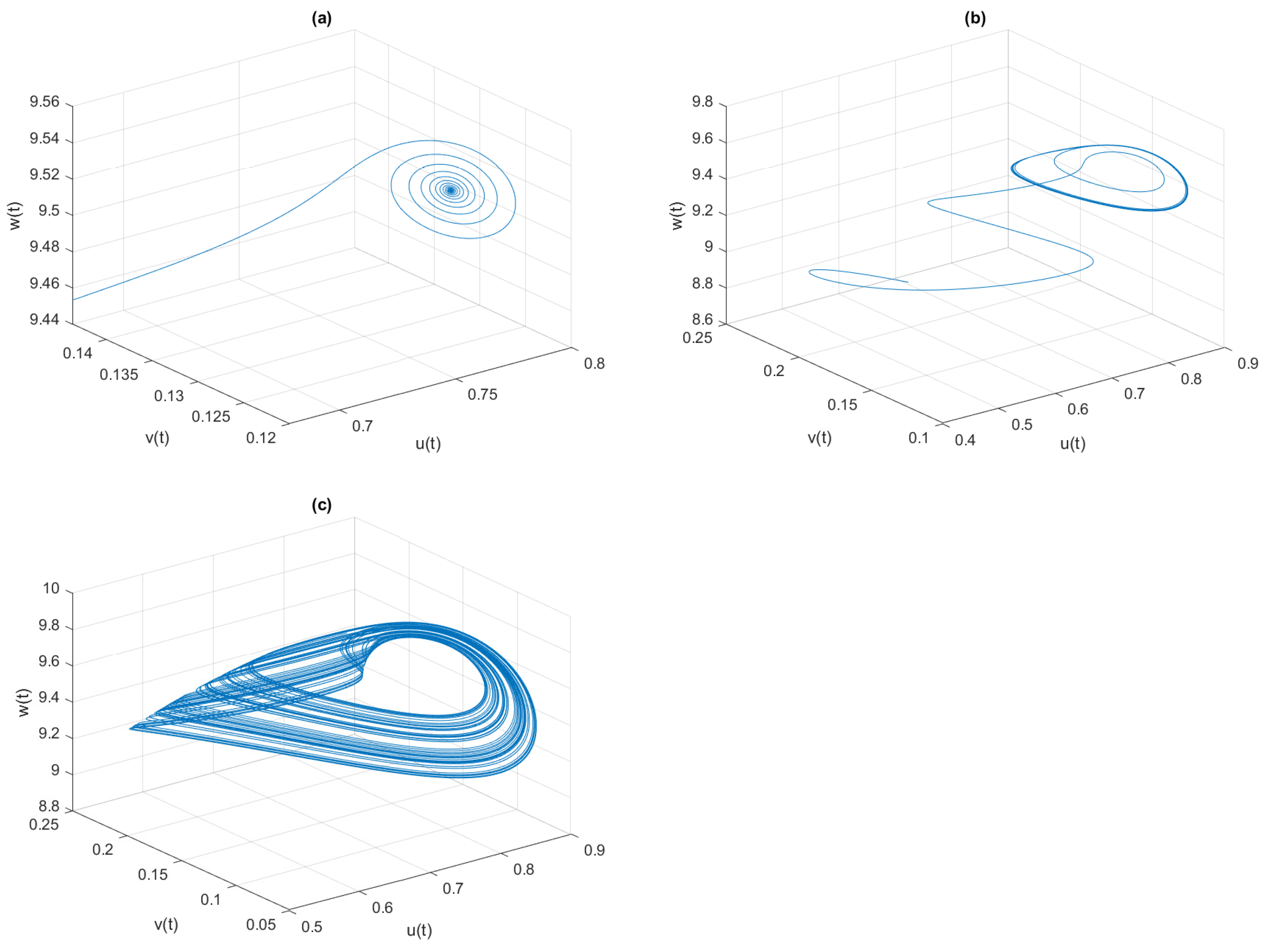
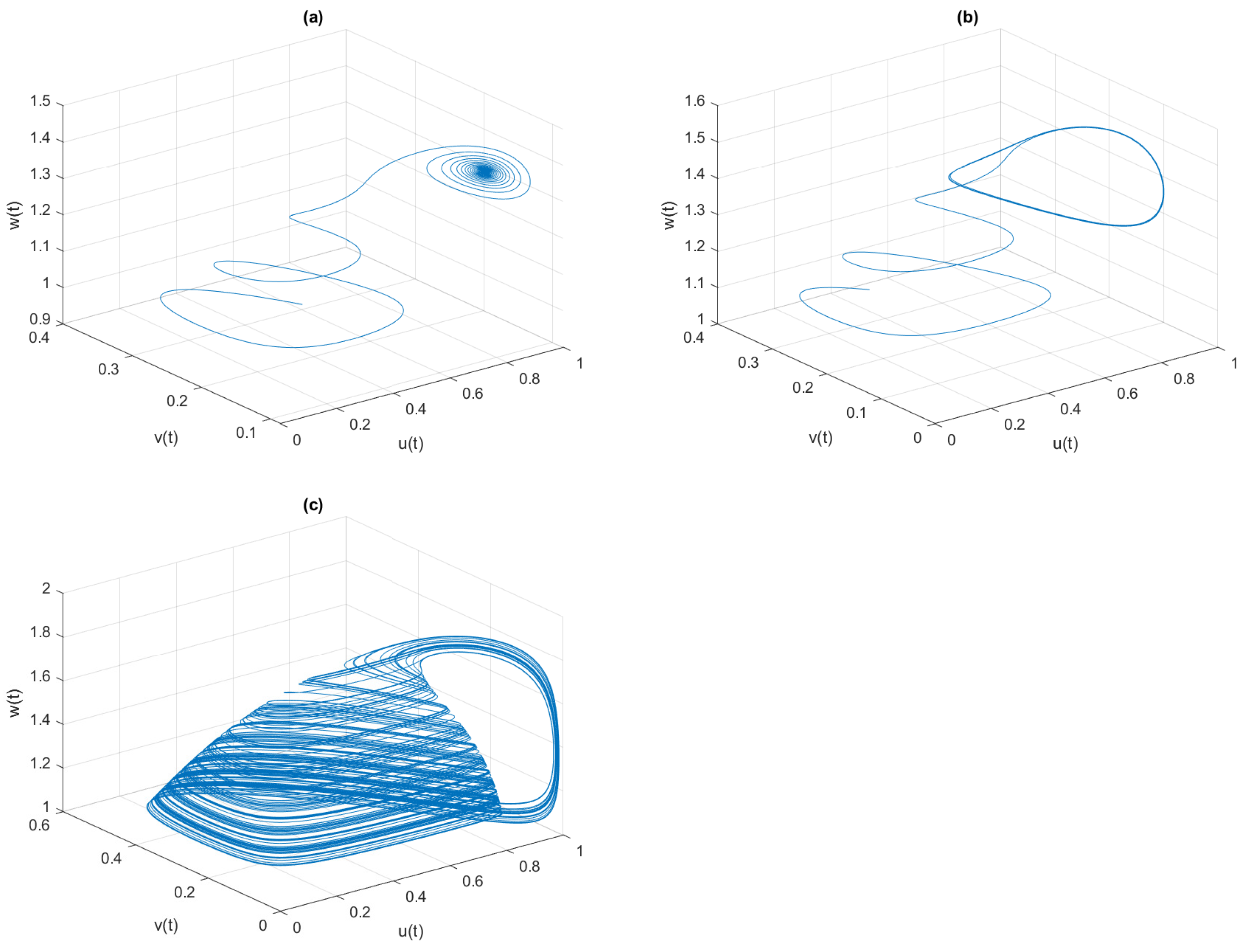
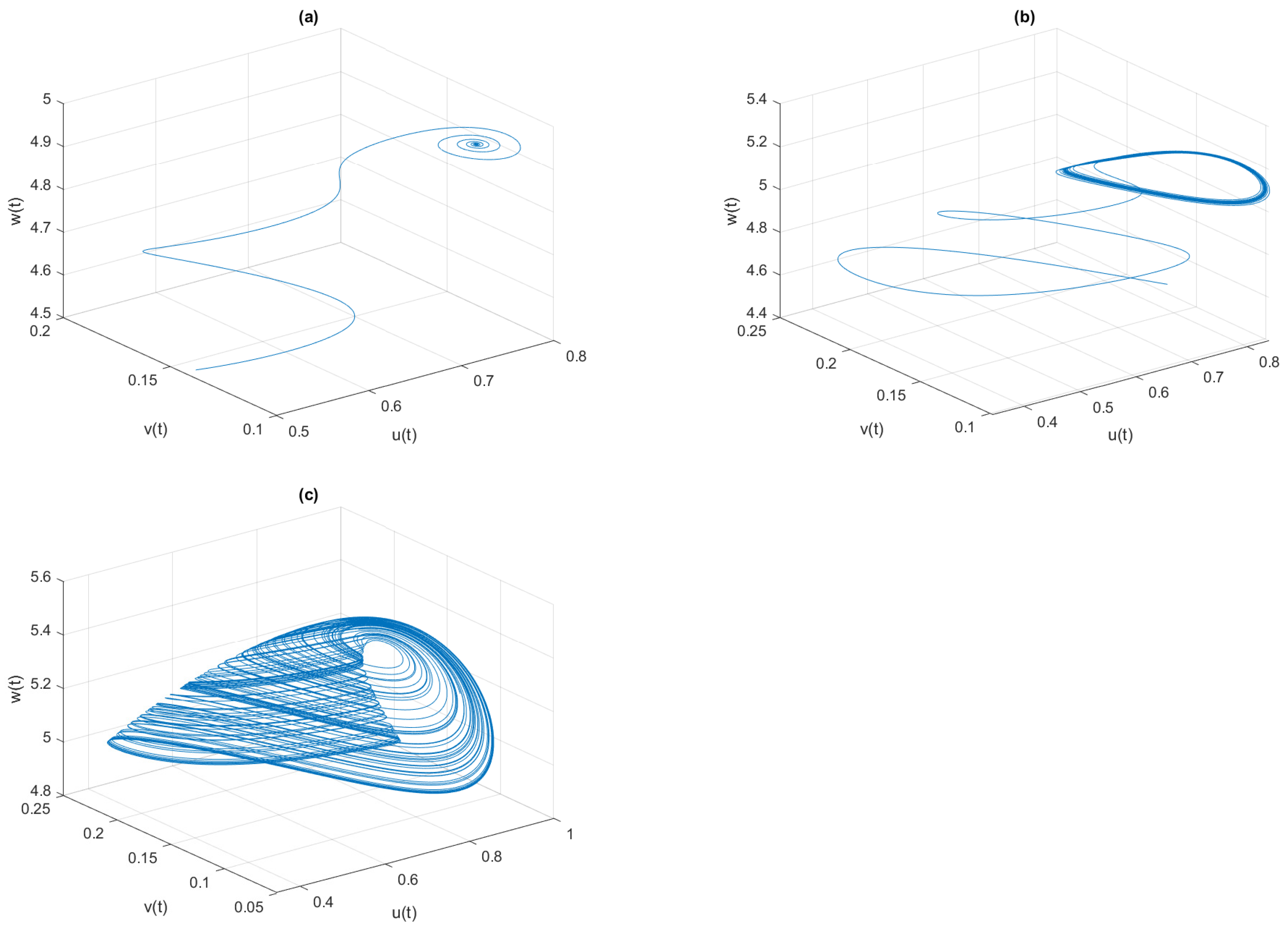
, the phase space diagrams of model (
4) are given in
Figure 7, where the system is LAS for
, periodic oscillation for
, and chaotic oscillation for
In
Figure 8, we draw a two-parameter bifurcation diagram between fear
and its COE parameter
. For smaller values of
, we observe that the system changes its state from chaotic oscillation to a stable one through period-halving with
increasing, while for higher values, the system remains chaotic. The chaotic phenomena can also be controlled by increasing fear parameter
. Similarly, we analyze the impact of the special predator-induced fear on the middle predator’s growth and its COE, while the fear of the middle predator on the prey is absent
. In this case,
Figure 9 displays the bifurcation diagram and LLE of system (
4) with respect to
when
We should note that the system becomes chaotic from stable oscillation through period-doubling as COE parameter
increases from 0 to 3. The phase space diagrams of model (
4) are drawn in
Figure 10 for different values of
, where the system is LAS for
, the periodic oscillation is for
, and the chaotic oscillation is for
Moreover, we notice that as
further increases, the system enters into periodic oscillation from chaotic oscillation.
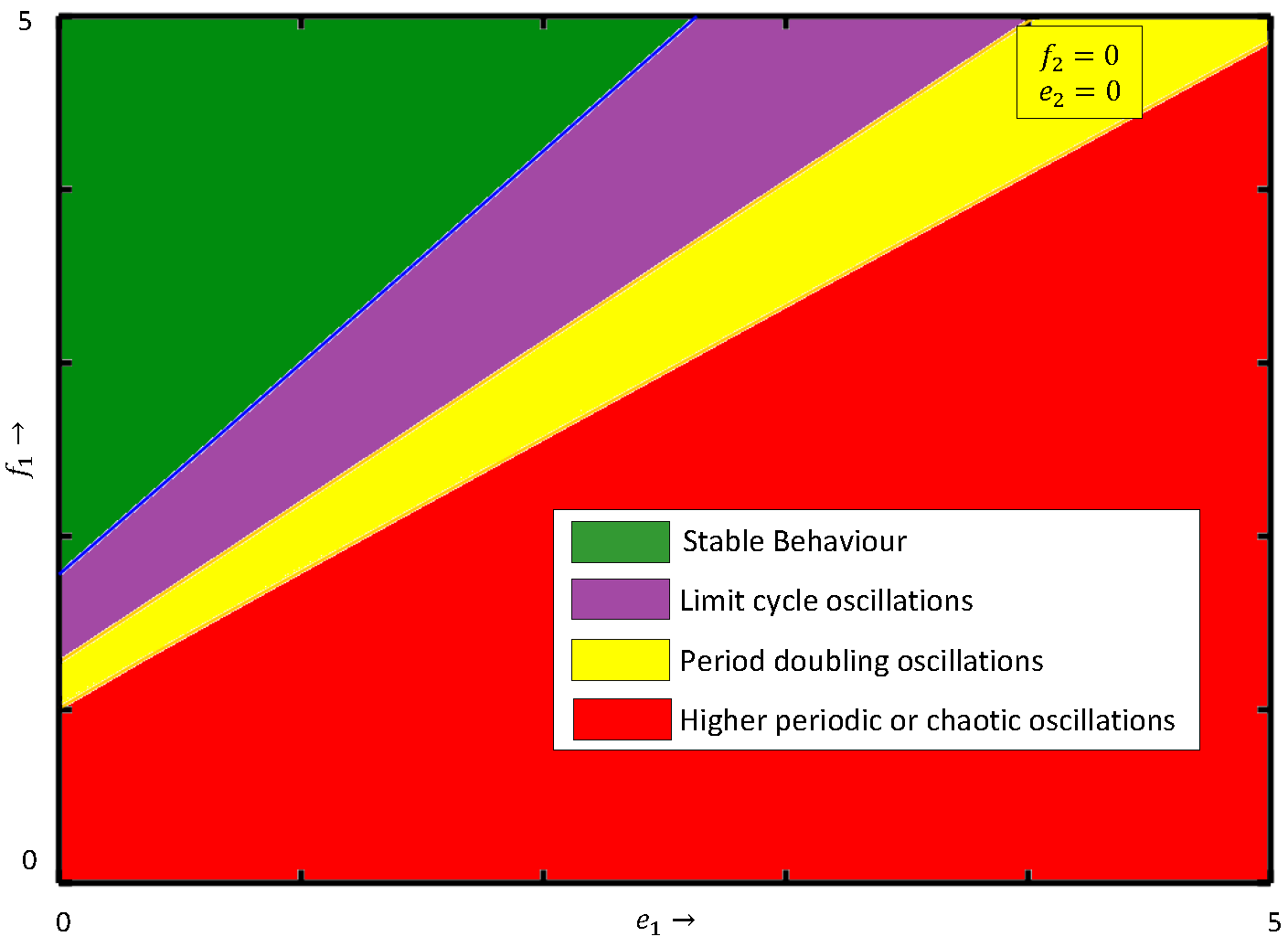
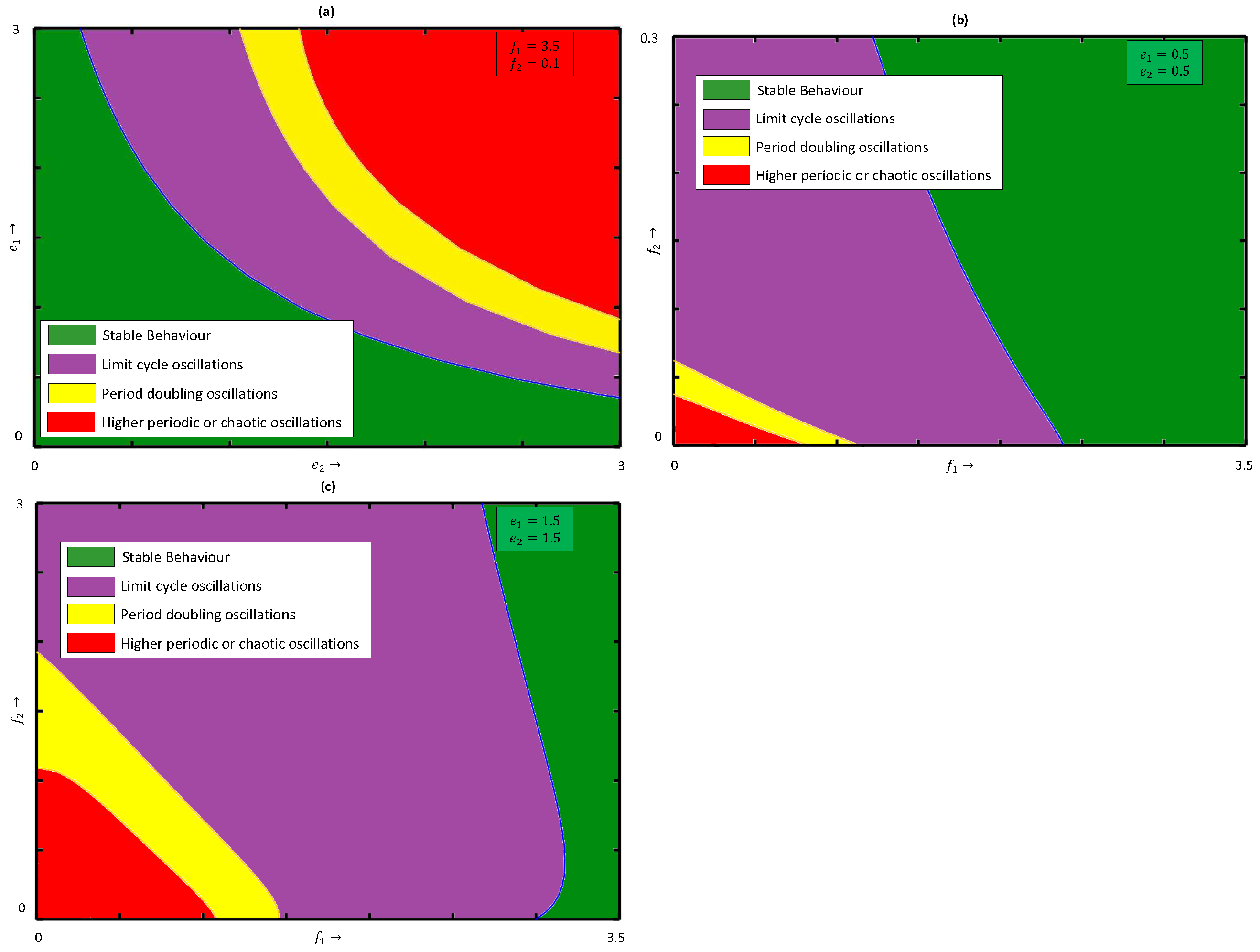
Next, we examine how the system dynamics are influenced when both fear-induced COE factors are present. To emphasize this, we draw the two-parameter bifurcation diagram in the
parametric plane of system (
4) in the presence of the fear-induced COE in the prey species as well as in the middle predator, see
Figure 11a. We observe that for lower values of COE parameters,
and
, system (
4) shows stable behavior and it becomes chaotic through period-doubling as
(
) increases, i.e., both
and
have destabilizing effects for fixed fear parameters
.
Figure 11b,c display the two-parameter bifurcation diagram in the
parametric plane of system (
4) with
and
, respectively. The figure clearly shows that as
or
increases, the stability region in the
parametric plane gradually decreases. To protect biodiversity, artificially introduced fear through vocalization plays a crucial role. In reality, species learn about the introduced artificial fear from past experiences and pass it on to future generations as fear-induced COE parameters
increase, and as a result, a greater amount of prey and middle predators are consumed by middle and special predators, respectively, in the ecosystem. This situation leads a species population to an extinction stage. It is clear that the system exhibits chaos for higher values of
, and it can be controlled by increasing the fear parameter. For a better understanding of the impacts of
and
on the dynamics of system (
4), we plot the bifurcation diagram with respect to
and
in
Figure 12. It can be observed from
Figure 12 that an increase in parameter
(
) makes the system chaotic from stable through period-doubling. If
we observe that the system exhibits stable behavior for
, limits cycle oscillations for
, and higher periodic or chaotic oscillations for
Similarly, if
we observe that the system exhibits stable behavior for
, limits cycle oscillations for
, and higher periodic or chaotic oscillations for
The chaotic behaviors occurring in
Figure 12 can be confirmed by the positive LLE values shown in
Figure 13. To show the impacts of parameters
and
on the system dynamics, we draw the phase space diagrams of model (
4) for different values of
in
Figure 14. The values of LLE presented in
Figure 6b,
Figure 9b and
Figure 13 are summarized in
Table 1, where the positive values confirm the chaotic dynamics of model (
4).
6.2. Effects of Time-Delays
Now, we explore the dynamics of system (
22) by varying delay parameters
and fixing other system parameters as given in (
29). We can obtain
and critical delay
According to Theorem 10, when
passes its critical value
, the interior equilibrium
loses its stability and a periodic solution occurs through the Hopf bifurcation, which is given in
Figure 15. For low values of
, the system is in a stable state and then the system loses its stability as
passes its critical value.
Figure 15c displays the bifurcation diagram of model (
22) with respect to
which indicates that the system (
22) is stable for
; limit cycle oscillations for
and higher periodic or chaotic oscillations for
Thus, the gestation delay in special predators has a destabilizing effect.
6.3. Discussion
System (
4) has four ecologically possible equilibria for system parameters as given in (
29). Coexistence equilibrium is the only one that is stable. This means that all solutions to system (
4) start with different initial populations that approach equilibrium. In the case without COEs among species, the system becomes stable from its chaotic nature as we increase the fear parameters
and
. Ecologically, middle and special predators can access lower numbers of prey. As a result, none of the species could become extinct in the system while maintaining a positive density level. In the absence of induced fear on the middle predator
, the system maintains stability for lower values of
, and the system shows periodic or chaotic oscillations as
increases further. Similarly, in the absence of induced fear on the prey,
, the system transitions from a stable state to a chaotic state as
increases and shuttles down in periodic oscillation as
increases further. When both fear factors are present constantly, we observe that for lower parameters,
and
, the system shows stable behavior and it becomes chaotic through period-doubling as
and/or
increase. A greater number of prey and middle predators are consumed by middle and special predators, respectively, in the ecosystem as the COE effect among species increases, and this situation leads to an extinction stage of the species. Thus, both
and
have stabilizing effects, while both
and
have destabilizing effects. For a lower time delay, the system is stable. When the time delay increases, the system exhibits higher periodic or chaotic oscillations. The bifurcation diagram of the middle predator is only used because all three species in the system have bifurcation diagrams that are symmetric. To protect biodiversity and manage ecosystems, we can use fear through artificial vocalization.




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}