
Terahertz Waves Enhance the Permeability of Sodium Channels

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Method
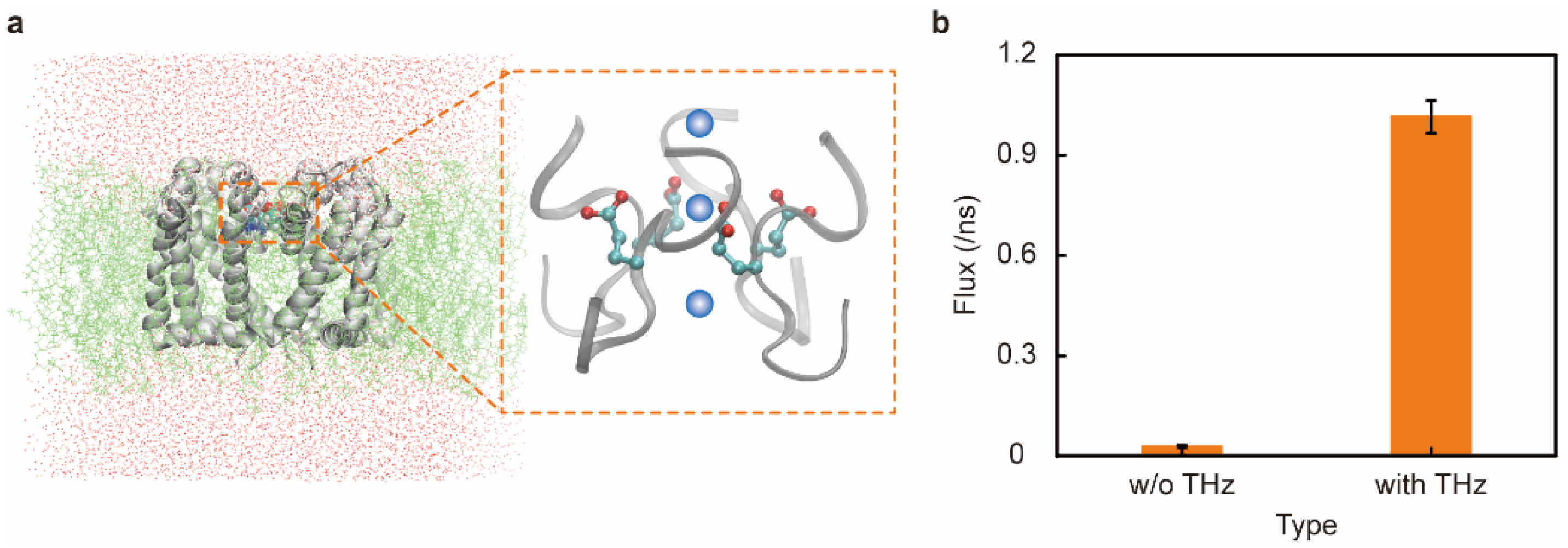
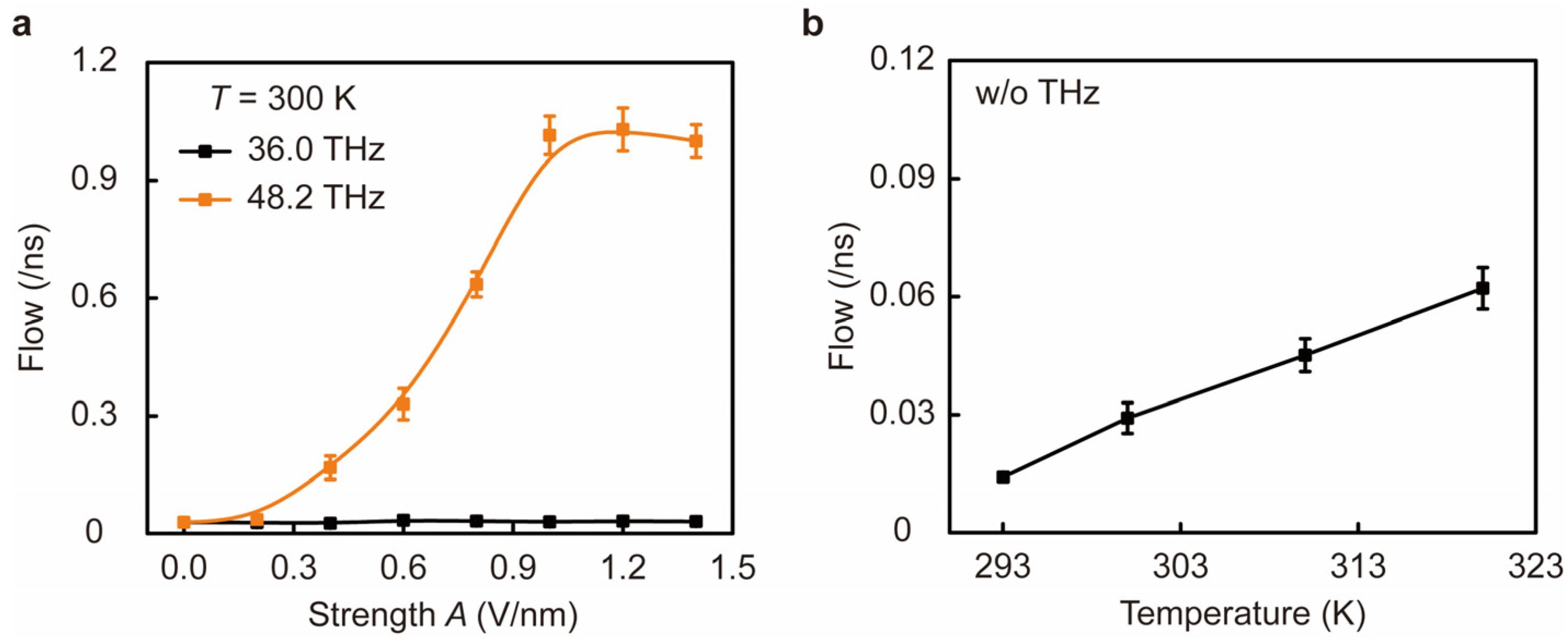
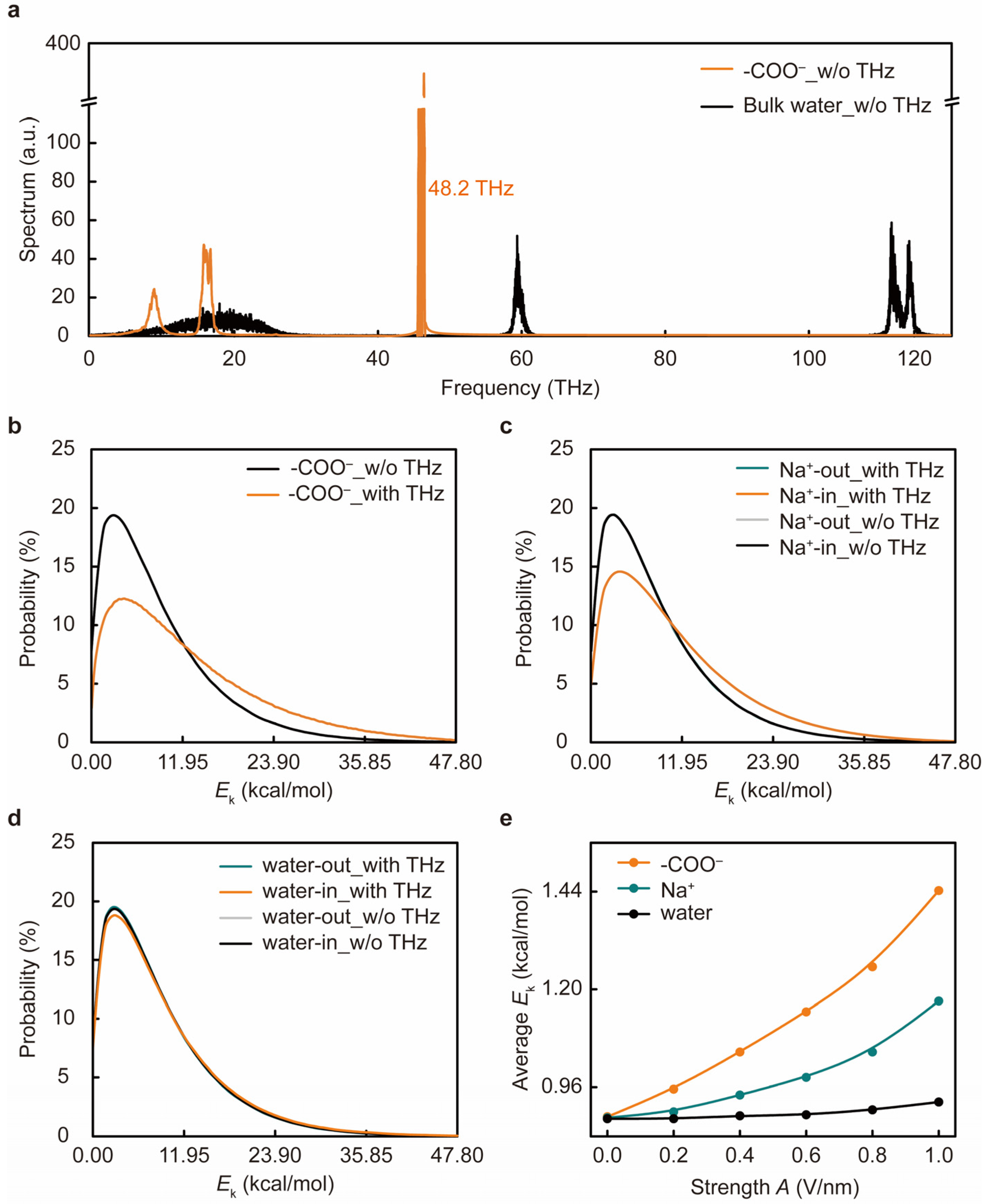
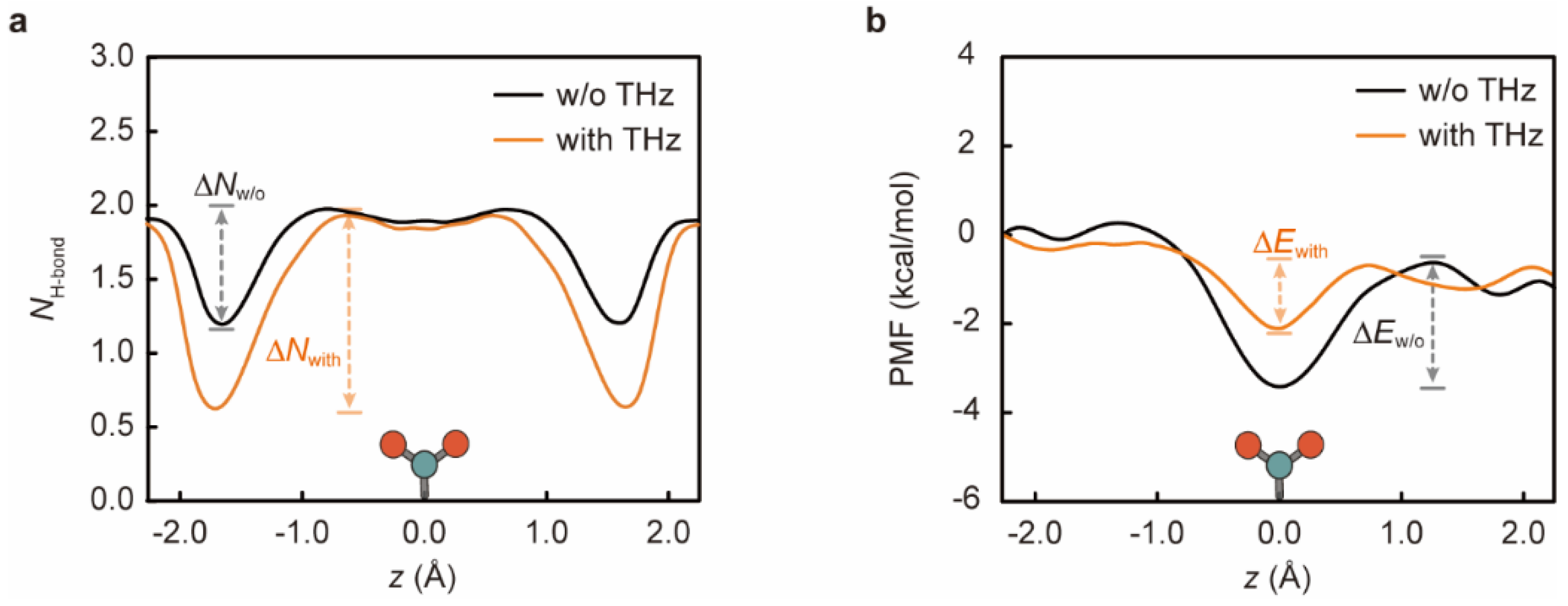
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanukoglu, I.; Hanukoglu, A. Epithelial sodium channel (ENaC) family: Phylogeny, structure–function, tissue distribution, and associated inherited diseases. Gene 2016, 579, 95–132. [Google Scholar] [CrossRef] [PubMed]
- Remme, C.A.; Bezzina, C.R. Sodium channel (dys) function and cardiac arrhythmias. Cardiovasc. Ther. 2010, 28, 287–294. [Google Scholar] [CrossRef]
- Sun, P.; Zheng, F.; Zhu, M.; Wang, K.; Zhong, M.; Wu, D.; Zhu, H. Realizing synchronous energy harvesting and ion separation with graphene oxide membranes. Sci. Rep. 2014, 4, 5528. [Google Scholar] [CrossRef] [PubMed]
- Corry, B. Designing carbon nanotube membranes for efficient water desalination. J. Phys. Chem. B 2008, 112, 1427–1434. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Zhou, J.; Lu, X.; Corry, B. Bioinspired graphene nanopores with voltage-tunable ion selectivity for Na+ and K+. ACS Nano 2013, 7, 10148–10157. [Google Scholar] [CrossRef] [PubMed]
- Koch, S.; Bierbrauer, J.; Haas, K.; Wolter, S.; Grosskreutz, J.; Luft, F.; Spies, C.; Fielitz, J.; Weber-Carstens, S. Critical illness polyneuropathy in ICU patients is related to reduced motor nerve excitability caused by reduced sodium permeability. Intensive Care Med. Exp. 2016, 4, 10. [Google Scholar] [CrossRef]
- Daviet, F.; Blin, M.G.; Fallague, K.; Bachelier, R.; Laforêt, M.; Carré, M.; Poitevin, S.; Dignat-George, F.; Blot-Chabaud, M.; Bardin, N.; et al. Sera from patients with minimal change disease increase endothelial permeability to sodium. Kidney Int. Rep. 2020, 5, 1071–1075. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Shi, H.; Tonggu, L.; El-Din, T.M.G.; Lenaeus, M.; Zhao, Y.; Yoshioka, C.; Zheng, N.; Catterall, W.A. Structure of the cardiac sodium channel. Cell 2020, 180, 122–134. [Google Scholar] [CrossRef]
- Fozzard, H.A.; Makielski, J.C. The electrophysiology of acute myocardial ischemia. Annu. Rev. Med. 1985, 36, 275–284. [Google Scholar] [CrossRef]
- Rush, A.M.; Dib-Hajj, S.D.; Liu, S.; Cummins, T.R.; Black, J.A.; Waxman, S.G. A single sodium channel mutation produces hyper-or hypoexcitability in different types of neurons. Proc. Natl. Acad. Sci. USA 2006, 103, 8245–8250. [Google Scholar] [CrossRef] [Green Version]
- Payandeh, J.; El-Din, T.M.G.; Scheuer, T.; Zheng, N.; Catterall, W.A. Crystal structure of a voltage-gated sodium channel in two potentially inactivated states. Nature 2012, 486, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Naylor, C.E.; Bagnéris, C.; DeCaen, P.G.; Sula, A.; Scaglione, A.; Clapham, D.E.; Wallace, B. Molecular basis of ion permeability in a voltage-gated sodium channel. EMBO J. 2016, 35, 820–830. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhao, X.; Yang, K.; Liu, Y.; Liu, Y.P.; Fu, W.; Luo, Y. Biomedical applications of terahertz spectroscopy and imaging. Trends Biotechnol. 2016, 34, 810–824. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.S.; Callebaut, H.; Kumar, S.; Hu, Q. 3.4-THz quantum cascade laser based on longitudinal-optical-phonon scattering for depopulation. Appl. Phys. Lett. 2003, 82, 1015–1017. [Google Scholar] [CrossRef]
- Wei, L.; Yu, L.; Jiaoqi, H.; Guorong, H.; Yang, Z.; Weiling, F. Application of terahertz spectroscopy in biomolecule detection. Front. Lab. Med. 2018, 2, 127–133. [Google Scholar] [CrossRef]
- Xiang, Z.; Tang, C.; Chang, C.; Liu, G. A new viewpoint and model of neural signal generation and transmission: Signal transmission on unmyelinated neurons. Nano Res. 2021, 14, 590–600. [Google Scholar] [CrossRef]
- Liu, G.; Chang, C.; Qiao, Z.; Wu, K.; Zhu, Z.; Cui, G.; Peng, W.; Tang, Y.; Li, J.; Fan, C. Myelin sheath as a dielectric waveguide for signal propagation in the mid-infrared to terahertz spectral range. Adv. Funct. 2019, 29, 1807862. [Google Scholar] [CrossRef]
- Sun, L.; Li, Y.; Yu, Y.; Wang, P.; Zhu, S.; Wu, K.; Liu, Y.; Wang, R.; Min, L.; Chang, C. Inhibition of cancer cell migration and glycolysis by terahertz wave modulation via altered chromatin accessibility. Research 2022, 2022, 986067. [Google Scholar] [CrossRef]
- Li, N.; Peng, D.; Zhang, X.; Shu, Y.; Zhang, F.; Jiang, L.; Song, B. Demonstration of biophoton-driven DNA replication via gold nanoparticle-distance modulated yield oscillation. Nano Res. 2021, 14, 40–45. [Google Scholar] [CrossRef]
- Tan, X.; Zhong, Y.; Li, R.; Chang, C. Neuromodulation of Chemical Synaptic Transmission Driven by THz Photons. Research 2022, 2022, 0010. [Google Scholar] [CrossRef]
- Zhang, Q.L.; Yang, R.Y.; Wang, C.L.; Hu, J. Ultrafast active water pump driven by terahertz electric fields. Phys. Rev. Fluid 2022, 7, 114202. [Google Scholar] [CrossRef]
- Pan, Z.; Liu, W.; Yu, L.; Xie, Z.; Sun, Q.; Zhao, P.; Chen, D.; Fang, W.; Liu, B. Resonance-Induced Reduction of Interfacial Tension of Water-Methane and Improvement of Methane Solubility in Water. Langmuir 2022, 38, 13594–13601. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.Y.; Huang, Z.Q.; Wei, S.N.; Zhang, Q.L.; Jiang, W.Z. The resonant heating of heavy water solutions under the terahertz pulse irradiation. J. Mol. Liq. 2017, 229, 148–152. [Google Scholar] [CrossRef]
- Wu, K.; Qi, C.; Zhu, Z.; Wang, C.; Song, B.; Chang, C. Terahertz wave accelerates DNA unwinding: A molecular dynamics simulation study. J. Phys. Chem. Lett. 2020, 11, 7002–7008. [Google Scholar] [CrossRef] [PubMed]
- Cherkasova, O.P.; Serdyukov, D.S.; Nemova, E.F.; Ratushnyak, A.S.; Kucheryavenko, A.S.; Dolhanova, I.N.; Xu, G.; Skorobogatiy, M.; Reshetov, I.V.; Timashev, P.S. Cellular effects of terahertz waves. J. Biomed. Opt. 2021, 26, 090902. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Hao, Y.H.; Peng, R.Y. Advances in the biological effects of terahertz wave radiation. Mil. Med. Res. 2014, 1, 26. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Chang, C.; Shu, Y.; Song, B. Transition to a superpermeation phase of confined water induced by a terahertz electromagnetic wave. J. Phys. Chem. Lett. 2019, 11, 256–262. [Google Scholar] [CrossRef]
- Zhu, Z.; Chen, C.; Chang, C.; Song, B. Terahertz-Light Induced Structural Transition and Superpermeation of Confined Monolayer Water. ACS Photonics 2020, 8, 781–786. [Google Scholar] [CrossRef]
- Sun, T.; Zhu, Z. Light resonantly enhances the permeability of functionalized membranes. J. Membr. Sci. 2022, 662, 121026. [Google Scholar] [CrossRef]
- Liu, X.; Qiao, Z.; Chai, Y.; Zhu, Z.; Wu, K.; Ji, W.; Li, D.; Xiao, Y.; Mao, L.; Chang, C.; et al. Nonthermal and reversible control of neuronal signaling and behavior by midinfrared stimulation. Proc. Natl. Acad. Sci. USA 2021, 118, e2015685118. [Google Scholar] [CrossRef]
- Hu, Z.H.; Lv, W.P.; Hui, D.X.; Wang, X.J.; Wang, Y.N. Permeability enhancement of the KcsA channel under radiation of a terahertz wave. Phys. Rev. E 2022, 105, 024104. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chang, C.; Zhu, Z.; Sun, L.; Fan, C. Terahertz wave enhances permeability of the voltage-gated calcium channel. J. Am. Chem. Soc. 2021, 143, 4311–4318. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Bo, W.; Wang, K.; Wang, S. Theoretical investigation on the effect of terahertz wave on Ca2+ transport in the calcium channel. Iscience 2022, 25, 103561. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, Z.; Sun, L.; Xiang, Z.; Chang, C.; Fan, C. Physicochemical Insights on Terahertz Wave Diminished Side Effects of Drugs from Slow Dissociation. ACS Nano 2022, 16, 8419–8426. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.L.; Yang, R.Y.; Jiang, W.Z.; Huang, Z.Q. Fast water channeling across carbon nanotubes in far infrared terahertz electric fields. Nanoscale 2016, 8, 1886–1891. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.L.; Jiang, W.Z.; Liu, J.; Miao, R.D.; Sheng, N. Water transport through carbon nanotubes with the radial breathing mode. Phys. Rev. Lett. 2013, 110, 254501. [Google Scholar] [CrossRef] [PubMed]
- Payandeh, J.; Scheuer, T.; Zheng, N.; Catterall, W.A. The crystal structure of a voltage-gated sodium channel. Nature 2011, 475, 353–358. [Google Scholar] [CrossRef]
- Kariev, A.M.; Green, M.E. Quantum calculations on ion channels: Why are they more useful than classical calculations, and for which processes are they essential? Symmetry 2021, 13, 655. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Pálla, S.; Smith, J.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef]
- Torrie, G.M.; Valleau, J.P. Monte Carlo free energy estimates using non-Boltzmann sampling: Application to the sub-critical Lennard-Jones fluid. Chem. Phys. Lett. 1974, 28, 578–581. [Google Scholar] [CrossRef]
- Torrie, G.M.; Valleau, J.P. Nonphysical sampling distributions in Monte Carlo free-energy estimation: Umbrella sampling. J. Comput. Phys. 1977, 23, 187–199. [Google Scholar] [CrossRef]
- Allen, T.W.; Andersen, O.S.; Roux, B. Molecular dynamics-potential of mean force calculations as a tool for understanding ion permeation and selectivity in narrow channels. Biophys. Chem. 2006, 124, 251–267. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.A. Einstein-Smoluchowski diffusion equation: A discussion. Phys. Scr. 2004, 70, 120. [Google Scholar] [CrossRef]
- Dill, K.; Bromberg, S. Molecular Driving Forces: Statistical Thermodynamics in Biology, Chemistry, Physics, and Nanoscience; Garland Science: New York, NY, USA, 2010. [Google Scholar]
- Boulard, B.; Kieffer, J.; Phifer, C.C.; Angell, C.A. Vibrational spectra in fluoride crystals and glasses at normal and high pressures by computer simulation. J. Non. Cryst. Solids 1992, 140, 350–358. [Google Scholar] [CrossRef]
- Lin, S.T.; Blanco, M.; Goddard, W.A., III. The two-phase model for calculating thermodynamic properties of liquids from molecular dynamics: Validation for the phase diagram of Lennard-Jones fluids. J. Chem. Phys. 2003, 119, 11792–11805. [Google Scholar] [CrossRef]
- Du, J.; Xiang, Y. Effect of strontium substitution on the structure, ionic diffusion and dynamic properties of 45S5 bioactive glasses. J. Non. Cryst. Solids 2012, 358, 1059–1071. [Google Scholar] [CrossRef]
- Caine, S.; Heraud, P.; Tobin, M.J.; McNaughton, D.; Bernard, C.C.A. The application of Fourier transform infrared microspectroscopy for the study of diseased central nervous system tissue. Neuroimage 2012, 59, 3624–3640. [Google Scholar] [CrossRef]
- Yang, R.Y.; Jiang, W.Z.; Huo, P.Y. Anisotropic energy absorption from mid-infrared laser pulses in constrained water systems. J. Mol. Liq. 2022, 366, 120286. [Google Scholar] [CrossRef]
- Wojtkowiak, K.; Jezierska, A.; Panek, J.J. Interactions between Artificial Channel Protein, Water Molecules, and Ions Based on Theoretical Approaches. Symmetry 2022, 14, 691. [Google Scholar] [CrossRef]
- Zhu, Z.; Guo, H.K.; Jiang, X.K.; Chen, Y.C.; Song, B.; Zhu, Y.; Zhuang, S. Reversible hydrophobicity-hydrophilicity transition modulated by surface curvature. J. Phys. Chem. Lett. 2018, 9, 2346–2352. [Google Scholar] [CrossRef]
- Guo, Y.W.; Qin, J.Y.; Hu, J.H.; Cao, J.H.; Zhu, Z.; Wang, C.L. Molecular rotation-caused autocorrelation behaviors of thermal noise in water. Nucl. Sci. Tech. 2020, 31, 53. [Google Scholar] [CrossRef]
- Baştuğ, T.; Chen, P.C.; Patra, S.M.; Kuyucak, S. Potential of mean force calculations of ligand binding to ion channels from Jarzynski’s equality and umbrella sampling. J. Chem. Phys. 2008, 128, 04B614. [Google Scholar] [CrossRef] [PubMed]
- Ke, S.; Zangerl, E.M.; Stary-Weinzinger, A. Distinct interactions of Na+ and Ca2+ ions with the selectivity filter of the bacterial sodium channel NaVAb. Biochem. Biophys. Res. Commun. 2013, 430, 1272–1276. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Huang, X.; Qian, Y.; Chen, W.C.; Wen, L.; Jiang, L. Engineering Smart Nanofluidic Systems for Artificial Ion Channels and Ion Pumps: From Single-Pore to Multichannel Membranes. Adv. Mater. 2020, 32, 1904351. [Google Scholar] [CrossRef]
- Wang, H.; Yu, H.; Kim, Y.; Chen, T. Asymmetry in Muscle Strength, Dynamic Balance, and Range of Motion in Adult Symptomatic Hip Dysplasia. Symmetry 2022, 14, 748. [Google Scholar] [CrossRef]
- Esgalhado, F.; Batista, A.; Vassilenko, V.; Russo, S.; Ortigueira, M. Peak Detection and HRV Feature Evaluation on ECG and PPG Signals. Symmetry 2022, 14, 1139. [Google Scholar] [CrossRef]
- Xie, C.; Jauhari, S.; Mora, A. Popularity and performance of bioinformatics software: The case of gene set analysis. BMC Bioinform. 2021, 22, 191. [Google Scholar] [CrossRef]
- Song, B.; Jiang, L. The macroscopic quantum state of ion channels: A carrier of neural information. Sci. China Mater. 2021, 64, 2572–2579. [Google Scholar] [CrossRef]
- Hummer, G.; Rasaiah, J.C.; Noworyta, J.P. Water conduction through the hydrophobic channel of a carbon nanotube. Nature 2001, 414, 188–190. [Google Scholar] [CrossRef]
- York, D.M.; Darden, T.A.; Pedersen, L.G. The effect of long-range electrostatic interactions in simulations of macromolecular crystals: A comparison of the Ewald and truncated list methods. J. Chem. Phys. 1993, 99, 8345–8348. [Google Scholar] [CrossRef]
- Guillot, B.; Guissani, Y. Boson peak and high frequency modes in amorphous silica. Phys. Rev. Lett. 1997, 78, 2401. [Google Scholar] [CrossRef]
- Praprotnik, M.; Janežič, D.; Mavri, J. Temperature dependence of water vibrational spectrum: A molecular dynamics simulation study. J. Phys. Chem. A 2004, 108, 11056–11062. [Google Scholar] [CrossRef]
- Zhu, F.; Hummer, G. Convergence and error estimation in free energy calculations using the weighted histogram analysis method. J. Comput. Chem. 2012, 33, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Rosenberg, J.M.; Bouzida, D.; Swendsen, R.H.; Kollman, P.A. The weighted histogram analysis method for free-energy calculations on biomolecules. I. The method. J. Comput. Chem. 1992, 13, 1011–1021. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Wang, L.; Li, Y.; Zhu, Z. Terahertz Waves Enhance the Permeability of Sodium Channels. Symmetry 2023, 15, 427. https://doi.org/10.3390/sym15020427
Zhao Y, Wang L, Li Y, Zhu Z. Terahertz Waves Enhance the Permeability of Sodium Channels. Symmetry. 2023; 15(2):427. https://doi.org/10.3390/sym15020427
Chicago/Turabian StyleZhao, Yan, Lei Wang, Yangmei Li, and Zhi Zhu. 2023. "Terahertz Waves Enhance the Permeability of Sodium Channels" Symmetry 15, no. 2: 427. https://doi.org/10.3390/sym15020427