
Structural Brain Asymmetries for Language: A Comparative Approach across Primates


1
Laboratoire de Psychologie Cognitive, CNRS/Université Aix-Marseille UMR7290, 13003 Marseille, France
2
Station de Primatologie CNRS-CELPHEDIA UAR846, 13790 Rousset, France
*
Author to whom correspondence should be addressed.
Symmetry 2022, 14(5), 876; https://doi.org/10.3390/sym14050876
Submission received: 26 November 2021
/
Revised: 8 April 2022
/
Accepted: 10 April 2022
/
Published: 25 April 2022
(This article belongs to the Special Issue Cognitive and Neurophysiological Models of Brain Asymmetry)
{kind=link}
Abstract
:Humans are the only species that can speak. Nonhuman primates, however, share some ‘domain-general’ cognitive properties that are essential to language processes. Whether these shared cognitive properties between humans and nonhuman primates are the results of a continuous evolution [homologies] or of a convergent evolution [analogies] remain difficult to demonstrate. However, comparing their respective underlying structure—the brain—to determinate their similarity or their divergence across species is critical to help increase the probability of either of the two hypotheses, respectively. Key areas associated with language processes are the Planum Temporale, Broca’s Area, the Arcuate Fasciculus, Cingulate Sulcus, The Insula, Superior Temporal Sulcus, the Inferior Parietal lobe, and the Central Sulcus. These structures share a fundamental feature: They are functionally and structurally specialised to one hemisphere. Interestingly, several nonhuman primate species, such as chimpanzees and baboons, show human-like structural brain asymmetries for areas homologous to key language regions. The question then arises: for what function did these asymmetries arise in non-linguistic primates, if not for language per se? In an attempt to provide some answers, we review the literature on the lateralisation of the gestural communication system, which may represent the missing behavioural link to brain asymmetries for language area’s homologues in our common ancestor.
1. Introduction
“Human being: n. a man, woman, or child of the species Homo sapiens, distinguished from other animals by superior mental development and language…” [The Oxford Pocket Dictionary of Current English, Oxford University Press 2009].
This definition states the obvious: humans are the only species able to speak, and thus, this fascinating cognitive faculty is considered as a key feature which divides us from other animals [1]. However, in recent years, the conception of one holistic language faculty has been contrasted with an alternative view that language may be the result of an assembly of cognitive properties that are domain-general and not specific to language [2,3]. Therefore, since nonhuman animals, especially primates, have been shown to share some of these domain-general cognitive properties, the research on nonhuman primates was reconsidered as a critical model to investigate language evolution [3,4,5]. Whether these shared cognitive properties between humans and nonhuman primates are the result of a continuous evolution (homologies) or of a convergent evolution (analogies) remains difficult to demonstrate. However, comparing their respective underlying structure—the brain—to determinate their similarity or divergence across species is critical to help increase the probability of either of the two hypotheses, respectively. In humans, language models congruently describe a few key hubs of language processing, namely Broca’s and Wernicke’s area, and their interconnection, the Arcuate Fasciculus [6,7,8,9]. However, in recent years, data have expanded to more distributed models, taking into account several different fibre tracts and regions crucial for language processing, as demonstrated by clinical cases [10] and questioned a localisationist viewpoint of language specialisation [11]. For example, the importance of regions in the interface between social cognition and communication like the Superior Temporal Sulcus and Geschwind’s territory in the Inferior Parietal Lobe are highlighted [12], as well as the anterior Insula cortex [13] and the anterior and mid-Cingulate Sulci [14]. Most of these structures share one fundamental feature: they are functionally and structurally specialised towards one hemisphere, mostly the left [7,13,14]. Specifically, ‘functional lateralisation’ refers to a more pronounced activation in one hemisphere than the other. ‘Structural lateralisation’ refers to a more pronounced volume, size, surface measurements, or neuron density of a given region in one hemisphere than the other. Although the link between structural and functional asymmetry remains in question [15,16], most authors usually consider that structural asymmetry (the size of the roads) of these regions might reflect the functional asymmetry (the traffic) for language tasks [13,17,18].
Thus, the fundamental question of ‘how language lateralisation has evolved’ is more relevant than ever in the light of the aforementioned findings about language organisation in the brain. This is where comparative studies on our primate cousins are of importance. In this view of language evolution, the different cognitive components of language could have evolved gradually, in opposition to a saltatory emergence [19]. In this evolutionary process, it is not excluded that ‘exaptation’ phenomena, where an opportunistic selective adaptation is piggybacked onto preexisting structures, could have played a crucial role [3]. Because brain tissue hardly fossilises, one fruitful way to look for intermediate steps is the comparison of brains between primate species, including humans, in order to infer potential features inherited from their shared common ancestors [20]. In other words, one can determine which brain architectures and behaviours are shared between us and other primates, suggesting its pre-existence before the emergence of our species. Comparative brain approaches for primate species have particularly focussed on neuroanatomy, given the well-known limitation for functional studies in apes and monkeys (e.g., techniques, ethics, sample size, reproductivity). The advent of non-invasive techniques such as Magnetic Resonance Imaging (MRI) favours in-vivo acquisitions on anesthetised subjects, allowing no limitation in terms of sample sizes and species diversity [21]. As a result, primate brain research has benefited in recent years from considerable increase in available MRI databases on large cohorts in many primate species, including macaques: Prime-De [22] and chimpanzees: www.chimpanzeebrain.org (accessed on 12 April 2022) [23]. Just as in human brain research, research on primate brain anatomy allows morphological quantification of regions in each hemisphere and determination of possible interhemispheric asymmetries. In this quest, an increased body of evidence highlights that different nonhuman primate species, such as chimpanzees or baboons, show human-like structural brain asymmetries for areas homologous to key language regions [20,24].
Thus, the question arises: for which function have these asymmetries in non-linguistic primates developed, if not for language per se? In humans, handedness for manual actions was for a long time considered a behavioural reflection of language-related brain lateralisation [25]. However, more recent studies indicate that the direction of handedness for manual actions poorly predicts language lateralisation, especially in left-handed humans. Indeed, 96% of right-handers and 70% of left-handers have their left hemisphere functionally specialised for most language functions [25]. In fact, handedness for manual actions was shown to be associated with the contralateral lateralisation of the motor hand area within the Central Sulcus, rather than key language areas [26]. Altogether, it is now acknowledged that direction of handedness might be independent from direction of language lateralisation [27,28,29]. As a result, comparative research on handedness for manual actions across primates might not be suitable for investigating phylogenetical origins of hemispheric language specialisation. To do so, it has been suggested that studying the manual lateralisation of gestural communication in nonhuman primates—and not handedness for manipulative actions—might constitute a more fruitful approach [30]. In fact, following the evolutionary framework on the gestural origins of language [31], the gestural communication system in nonhuman primates was found to share key features of domain-general processes, important for language, such as intentionality, referentiality, and learning flexibility [32,33,34]. Interestingly, production of communicative manual gestures has been found highly lateralised in favour of the right hand in both baboons and chimpanzees. In contrast, the handedness patterns for non-communicative manual actions in chimpanzees and baboons were found to be different from those found for communicative gestures at both the populational and individual levels [30]. These findings supported the idea that gestural communication in nonhuman primates may be related to a specific lateralised system for communication, which might be different from handedness for manipulative actions [33].
In addition, the sequential and hierarchical motor actions that underlie tool-use behaviours have been used as the basis for the emergence of communicative systems [35,36,37,38], and share neural substrates with the human language network [39]. A recent functional MRI study with human subjects showed that the use of learning tools improved a language syntax task and there is neural overlap of both behaviours in the basal ganglia [40].
Therefore, there is the necessity to take stock of what the work on neuroanatomical correlates of gestural communication and tool-use in nonhuman primates has provided, in order to test its supposed continuity with language lateralisation.
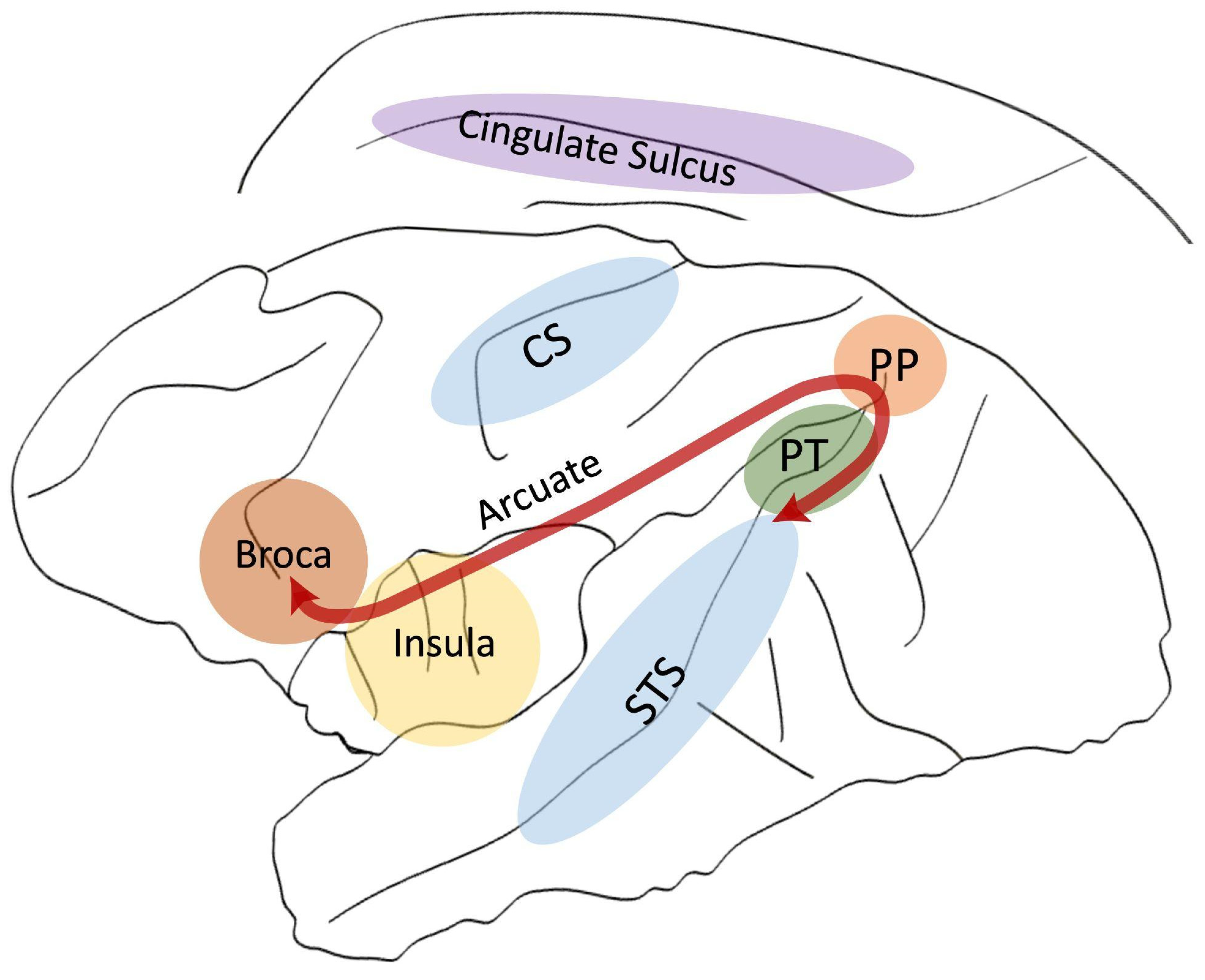
In this paper, we aimed to review the literature comparing structural brain asymmetries across primates for areas related to language in humans. We focus on the classical perisylvian language regions, namely the Planum Temporale, Broca’s Area, and the white matter tract that interconnects these two regions—The Arcuate Fasciculus. In this review, it is of importance to take also into account other key regions of the large, distributed language network beyond the perisylvian regions, such the Insula, the Cingulate Sulcus, and the Superior Temporal Sulcus (STS) as novel grey matter areas of interest. Finally, we consider the Central Sulcus (CS), which delimitates the primary motor from the primary somato-sensory cortex and include thus the mouth and lips motor areas as well as the motor hand area related to handedness (see Figure 1). After briefly describing their functions, we first review their structural and functional lateralisation in humans, including infants, to discuss whether structural markers can predict the functional lateralisation of language. In the next step, we compare these findings with nonhuman primates and discuss whether this asymmetric organisation is shared between species. Finally, we address the following question: If such brain lateralisation is shared with our non-linguistic primate cousins, for what behavioural functions did it evolve, if not for language? In an attempt to provide some elements of responses, we propose to review the literature about the lateralisation of the gestural communicative system, which could potentially constitute the ideal missing behavioural link with brain asymmetries for language in our common ancestor.
2. Planum Temporale
The most emblematic marker of the lateralised language organisation is the Planum Temporale (PT), which is located within the Sylvian fissure and is part of the auditory association cortex [41] and Wernicke’s area [42]. In adults, left hemispheric lesions in this region result in severe language comprehension and production deficits [43,44,45]. Therefore, many studies have shown the particular functional significance of the PT in the left hemisphere for a variety of auditory language processing [46], including the main perception component of the audio–motor loop for phonological processing [47]. Interestingly, in preverbal newborns, the functional implication of the left PT was highlighted from birth [17,48,49].
In a pioneering work by Geschwind and Levitsky [50], the PT was shown to be anatomically asymmetric: in 100 post-mortem brains, 65% of the left PT was larger than the right, which was confirmed by in-vivo MRI studies [46] and also highlighted in early development (Post-mortem Infants: [51,52]; in-vivo MRI infants: [53,54,55,56]; foetuses: [57,58]). These asymmetries later increase during development and are associated with language development [17,48,49,51]. PT asymmetry, is, therefore seen as a marker for human unique innate readiness to acquire language [17].
Indeed, in adults, a direct relationship was shown between the left PT’s size and functional asymmetry of language tasks [59]. In addition, an absence or reversed PT asymmetry has been linked to several language-related pathologies like dyslexia [60,61]. This function-structure relationship is however debated [13,15,16,62,63,64,65,66,67,68]. For example, Greve et al. [65] showed that regardless of the functional hemispheric dominance for language, the structural PT volume asymmetry is left biased. In contrast, Ocklenburg et al. [68] found that a higher density at a microstructural level of the left PT was associated with faster processing of auditory speed in the same area, as shown in EEG. More recently, Tzourio-Mazoyer et al. [16] demonstrated that, although the structural PT asymmetry is not predictive of its functional counterpart in a language task, an adjacent auditory area at the end of the Sylvian fissure is.
Several studies in nonhuman primates also showed striking human-like PT asymmetries in their homologous regions. Manual delineation of post-mortem brains [69], in-vivo MRI scans [70,71] as well as voxel-based morphology on MRI scans [23], showed larger left PTs in chimpanzees and in apes in general [72]. Recently, in-vivo MRI studies on adult and newborn baboons with manual PT delineation extended this finding to a shared feature between Old World monkeys and humans [24,73,74]. Interestingly, the asymmetry strength increased with age in this longitudinal study [74]. These results are questioning the PT asymmetry: (1) to be unique in humans and (2) to be marker for language development in newborns. Rather, the PT asymmetry might have evolved for a cognitive function shared between Old World monkeys, apes, and humans, which is at the core of language processing in humans. A potential candidate related to such a function may be communicative gesture. Indeed, Meguerditchian et al. [75] highlighted a relation between Planum Temporale grey matter volume asymmetry and hand preference for communicative gesture in chimpanzee. Moreover, the left PT asymmetry was also found related to handedness for tool-use but not for handedness for manipulative actions in chimpanzees, which required no structured sequence of motor actions [76].
3. Broca’s Area
Broca area and its left hemispheric specialisation was historically considered as the center of speech production [77]. This modular view of the neuronal basis of language was progressively questioned by the view that language involves a plastic and large distributed network [11,78] and even implicates the two hemispheres. However, it is still well acknowledged that Broca’s area in the left hemisphere remains a key component for language specialisation within its distributed neural network [79]. Interestingly, complementary work thereby highlighted Broca’s area as a lateralised interface between speech and multimodal motor integration, including gesture and mouth movements [80]. Broca’s area is also known for its involvement in motor planning, sequential and hierarchical organisation of behaviours, including syntax [81], tool-use [39,82], and sign language production, thus including manual and oro–facial gestures [83,84]. In infants, speech perception activates Broca’s area from very early development on as highlighted in MEG or functional MRI studies [48,85,86]. This activation before the babbling stage suggested that activity of this area is not due to motor learning but might drive the learning of complex sequences [86].
In contrast to the PT [see section above], a clear structural leftward asymmetry has not been reproducibly demonstrated [87,88], which may be due to natural variability between subjects in sulcal contours defining this area [87,89]. Cytoarchitectonic analyses, however, reported a leftward asymmetry of some parts of area 44 and/or area 45, which together form Broca’s area [90,91,92]. In contrast, other accounts state a rightward asymmetry for area 45 grey matter, which gets reduced during aging, especially due to a loss in the right hemisphere [93]. In development, an early structural primacy of right-sided dendrite systems shortly after birth and a progressive shift to left-sided primacy during years three to six was highlighted and related to critical periods for language acquisition [94]. A second study found leftward asymmetries on the cellular level very early from 1-year old infants on [92], which increased into an adult-like leftward asymmetry at 5 years for area 45 and 11 years for area 44. This maturational effect was suggested to be influenced by language practice and thus, the interhemispheric asymmetry of this area would continue to change throughout life [92]. Interestingly, because area 45 supports semantic processes and area 44 subserves syntactic processes in adults, some see in this maturational difference a neural underpinning of the earlier onset of semantics than syntax in children [79].
In great apes, a homologous region was documented in several studies, which described the precentral inferior sulcus, the inferior frontal sulcus, and the fronto-orbital sulcus as common borders of Broca’s homologue [95,96,97]. Like in humans, inquiries about Broca’s area’s structural asymmetry on a population-level remain inconsistent in apes due to interindividual variation in location and cytoarchitecture [98]. Leftward lateralisation was found at a macrostructural level [95] but not at a cytoarchitectonic level in a relative smaller sample size ([98]; but see also [99]). In monkeys, no data of structural asymmetry for this region has been reported so far. In fact, determining Broca’s homologue is challenging because the common borders of Broca’s homologue in apes are absent in monkeys. Nevertheless, we know from few detailed cytoarchitectonic studies in macaques that the two parts of Broca’s area 44 and 45 are respectively located in the fundus and lower caudal/posterior bank, and on the rostral/anterior side, of the most ventral part of the inferior arcuate sulcus IAS [100,101,102]. Electric stimulation of this region elicited oro-facial and finger movements. Therefore, together with cytoarchitectonic similarities, the region anterior to the ventral part of the IAS was proposed as an equivalent area 44 in macaques [101]. Recent studies even reported that Broca’s homologue’s activation preceded voluntary trained production of a vocalisation after intensive operant conditioning in juvenile rhesus monkeys [103]. Moreover, the use of positron emission tomography (PET) in three captive chimpanzees has revealed that begging food from a human by using either gestures, novel species atypical attention-getting sounds, or both simultaneously, activated a homologous region of Broca’s area (IFG) predominantly in the left hemisphere [104]. Some functions associated with Broca’s homologue’s lateralisation in nonhuman primates have been proposed in relation to tool-use processing and communication gesture production. Regarding tool-use, chimpanzees that performed better a tool-use task with their right hand showed a greater left-lateralisation of Broca’s homologue [105]. Such a link might be attributed to the typical Broca’s function described in humans, namely hierarchical organisation of behaviours involved in tool-use [81,82]. Regarding gestural communication, Meguerditchian et al. [75] highlighted a relation between Broca’s homologue grey matter volume asymmetry and hand preference for communicative gesture in chimpanzees. Similarly, in a recent study in baboons, we showed that variation in hand preference for communicative gesture—but not for non-communicative manipulative actions—is related to the anatomical variation of Broca’s homologue. Indeed, the right Broca’s portion of the IAS is deeper than the left for baboons communicating with their left hand and vice versa [106].
4. Arcuate Fasciculus
The Arcuate Fasciculus (AF) is a bundle of white matter, which arches dorsally around the Sylvian fissure, interconnecting Broca’s area in the frontal lobe with the Planum Temporale in the temporal lobe [107]. It was highlighted that the connectivity between language areas, due to the AF, is crucial. For example, the integrity of the AF might be more important for lesion recovery (e.g., strokes) than the integrity of grey matter regions that it is connecting [11]. In addition, as neurological cases have shown, the AF plays a key role in language processing in the left hemisphere, with lesion of the direct pathway also causing conduction aphasia [10] in deaf signers [108]. The AF is already present at birth [109] but matures slowly until late childhood [109,110]. In contemporary language models, the AF (or also called Dorsal Pathway) is often opposed to the Ventral Pathway, which interconnects roughly the same regions, but travels ventrally around the Sylvian fissure [78]. In contrast to the AF, the Ventral Pathway matures more rapidly in development and was also described to be phylogenetically more ancient [79,110]. It is assumed that the late maturation of the AF is due to the frontal portion, which is connected to Broca’s area and not fully myelinated until the age of seven [110,111]. In fact, controversy persists whether this portion is also already present at birth [109,112,113,114,115].
Regarding lateralisation, the human AF was found larger in the left hemisphere for a number of macroscopic and microscopic measurements like the number of streamlines, volume of the tract, fibre density, and mean fractional anisotropy in 60% of normal adult humans. The remaining 40% of the adult population shows either a reduced lateralisation to the left (20%) or no lateralisation at all (20%) [18,116,117,118,119,120]. In early development, the leftward AF is the most asymmetrical region of the developing white matter ([113,121], but see also [122]). Interestingly, the early leftward asymmetry in newborns was correlated with later language capacities in children [123,124,125]. Catani et al. [18] argued that the AF’s asymmetry represents a better structural marker for functional language specialisation than PT asymmetry [18]. This structure–function relationship is, however, debated [15,126,127,128,129]. For example, Verhelst et al. [129] demonstrated in a fixel based analysis that structural AF asymmetry did not differ between subjects with either right or left functional language hemispheric dominance.
Axon tracing in monkey brains and diffusion MRI in chimpanzee and monkey brains have highlighted the existence of the Arcuate Fasciculus across primates that interconnects frontal and temporal areas [130,131,132,133,134,135,136,137,138,139]. It is debated to what extent the AF’s strength, lateralization, and frontal and temporal termination sites differ between primate species. In fact, recent findings speak for a rather conserved organisation across primates (for a review, see [9]). Therefore, latest functional results suggest that language abilities allowing humans to name, conceptualise, and thus better remember sound would be shared across primates [139]. In order to highlight anatomical differences across primates, which could explain the human uniqueness for language, several authors conclude that the left AF lateralisation is the crux of the human-specific distinction [136,138,139]. In fact, Rilling et al. [133] did not find any AF asymmetry in three macaque and four chimpanzee subjects. However, by adding more chimpanzee subjects, the authors were able to report a left lateralised AF, which was still weaker than in humans [134]. This result remained unique regarding AF lateralisation in nonhuman primates [136,138,139]. In fact, this inconsistency across the literature about the presence or not of population-level leftward AF bias might be explained by the small sample size (i.e., only few subjects) usually included in those AF studies in apes, which makes it difficult to infer any bias at the population-level. Only studies including an increased sample size would help elucidate this debate.
5. Insula
The Insula cortex lies in the depth of the Sylvian fissure, which separates the temporal lobe from the parietal and frontal lobes. The anterior part of the Insula is implicated for different language processing functions (General: [43,140,141]; auditory processing: [142]; motor aspects: [143]; syntax: [144,145]; sign language: [108]). Interestingly, the Insula seems to be particularly involved in motor planning of speech as seen in pathologies [140,143]. Further, Lœvenbrueck et al. [146,147] highlighted in adults that prosodic pointing gesture activates Broca’s area as well the left anterior Insula [146,147].
Moreover, at the structural level, the volume asymmetry of this region may be associated with hemispheric dominance for language. In fact, Keller et al. [13] found that the size of the Insula could predict functional lateralisation for language in the same hemisphere in the majority of individuals. Therefore, the Insula was proposed as a more reliable marker for functional language specialisation than the Planum Temporale [13,148]. Although little is known about the functional implication of the Insula during language development in newborns and infants, several studies highlighted an early lateralisation of the Insula towards the left hemisphere [56]. Thanks to all the aforementioned data, the (anterior) Insula was established as a region of interest for studying linguistic (motor) processing. Additionally, the anterior Insula comprises Von Economo (VEN) and Fork neurons [149] that were for a long time thought to be uniquely human and implicated in social awareness. Therefore, the insula VEN and Fork neurons are often used for theories about the social origins of language [150].
While larger in humans, the insular cortex also exists in apes and monkeys, where the anterior portion especially, expanded during primate brain evolution [151]. Von Economo neurons, which were thought to be exclusive to apes [152], are also present in the anterior insular cortex of monkeys [150]. The authors argue that two distinct insular regions could be implicated in monkey communication [150]. First, a specific sensory-motor organisation for body parts in one part of the anterior Insula (the Idfa region) was found. Electric stimulation of this region elicited vocal cord movements in macaques. This region happens to be juxtaposed to a dorsal region, which receives inputs from area 44 (Part of Broca’s Area). Together, the two regions could be homologous to the human anterior insula, implicated in several language processing functions (see above). Second, Von Economo and Fork neurons in another part of the anterior insula [the Ial region] project into a region of the thalamus (PAG) that is involved in vocalisations [150]. However, the structural lateralisation of this region was poorly investigated in nonhuman primates. One rare study comparing the Insula structure between a handful of different primate species subjects demonstrated that the anterior portion of the Insula, in which Von Economo neurons were found, displayed a human unique left asymmetry [151]. Further studies with a larger sample size are needed to investigate whether the Insula and particularly its anterior portion is structurally lateralised in nonhuman primates in relation to planification of communication, especially gesture.
6. Cingulate Cortex/Sulcus
The Cingulate Sulcus is in the medial part of the cerebral cortex delimitated ventrally by the Cingulate cortex and dorsally by the paracentral lobe and the superior frontal cortex. Its anterior part is considered a hub for domain-general cognitive processing, such as counterfactual thinking, mentalizing, and language, including cognitive control on signals production [153,154,155,156]. Pioneering studies regarding language processes have shown that for the Anterior and Midcingulate cortex, (1) stimulations evoke orofacial and tongue movements [157] and (2) lesioned patients experience akinetic mutism, associated with an absent motivation to speak [158]. Little is known about the Cingulate cortex concerning direct language development. Rare results come from Lœvenbruck et al. [147] highlighting, in adults, the functional neuroanatomical activation of the left anterior Cingulate cortex, besides Broca’s area and the Insula in communicative pointing gestures [147]. However, the anterior Cingulate cortex was prominently shown to be important for joint attention in both adults and infants [159]. Joint attention is considered a prerequisite of the theory of mind as well as a prelinguistic communication act [159,160]. Interestingly, the hand, mouth, and tongue motor representations are grouped together around the caudal end of each vertical sulci departing from the Cingulate Sulcus [161], suggesting its key implication for a multimodal language system. In addition, 50% of human subjects present a Paracingulate Sulcus, located more rostrally above the anterior and Midcingulate Sulcus [162].
The presence of the Paracingulate Sulcus is lateralised in the human brain with nearly 70% located in the left hemisphere [14,162]. This lateralisation is influenced by genetic factors and the in-womb environment [163]. At the functional level, this human asymmetry has been shown to be correlated with the involvement of the left Cingulate cortex in language tasks in right-handed subjects [153,164].
Some authors have suggested that the anterior and Midcingulate cortices might have also played a role in language evolution. Loh et al. [155] hypothesised the existence of an evolutionary conserved ventrolateral frontal (around Broca’s region) and dorsomedial frontal (roughly the Midcingulate cortex) network, which enables cognitive control of vocalisations. In fact, it is known that innate reflexive vocalisations such as ‘screams’ and ‘shrieks’, are associated with the ‘cingulate vocalization pathway’ [165]. In fact, the ‘cingulate vocalisation area’ in the anterior and Midcingulate cortex are connected (1) to the periaque–ductal gray, which directly projects to premotor nuclei in the brainstem and controls laryngeal motoneurons, which elicits vocalisations; and (2) to the facial motor nuclei to also produce affective facial movements [155]. In addition, in nonhuman primates, innate calls can be evoked by anterior and Midcingulate cortex stimulations [166] and lesions impair the production of calls [167] similar as in humans. Additionally, as in the Insula cortex (see above), large spindle shaped Von Economo neurons are present in the anterior Cingulate cortex in humans, apes, and other mammals [152,168,169]. Together with the presence of Von Economo neurons in the anterior Insula, it has been proposed that Von Economo neurons may be implicated in primate communication [150]. In contrast to the Insula cortex, no Von Economo neurons have been found in the monkey’s Cingulate cortex yet. According to a recent comparative study including macaques, baboons, chimpanzees, and humans, the Cingulate Sulcus shows a highly conserved morphological antero-posterior organisation of vertical sulci or their precursor ‘dimples’ [14].
While, surprisingly, no structural asymmetries data on the Cingulate Sulcus are available so far in humans, significant population-level leftward asymmetries were found in the anterior portion of the Cingulate Sulcus in chimpanzee, whereas significant rightward biases were found in its posterior portion [170]. In contrast to humans, no population-level lateralisation for the presence of the Paracingulate Sulcus was found in chimpanzees [14]. Nevertheless, interindividual variation of the presence or absence of a Paracingulate Sulcus and variability of the intralimbic sulcus was associated with the production of attention-getting sounds and right handedness for gestural communication in chimpanzees [170]. It is, therefore, not to be excluded that the Cingulate Sulcus could be linked to precursors of human language such as communicative gesture and vocalisations.
7. Superior Temporal Sulcus (STS)
The Superior Temporal sulcus [STS] is a long fold separating the superior from the middle temporal lobe. It is important for a variety of social cognition tasks important for communication [171,172] containing both specialised regions for a particular task and regions that respond to several tasks simultaneously [171]. In fact, specialised areas of the STS have been shown to be implicated in the perception of voices in the right hemisphere [172], faces [173], biological motion [174], audiovisual integration [175], and in the theory of the mind [176,177]. Regarding speech perception in particular, three temporal voice areas are dispatched symmetrically in both hemispheres along an anterio-posterior gradient [178]. Distinct areas of the STS have also been implicated in language processing in hearing and deaf participants, especially in the left hemisphere [2,47,179]. In children, responses to voices in the STS and STG are strongly right-lateralised, an asymmetry which decreased with age [114,180]. This finding suggested that newborns rely for speech processing more on prosodic information, known to be processes in the right hemisphere, than for phonological information, processed in the left hemisphere (see also [181] for a right hemispheric STG temporal primacy in children in contrast to adults). Interestingly, together with Broca’s area, the posterior STS is proposed to constitute the neural network supporting syntactic processes [79], which, until the age of 10, does not process syntax and semantics independently [182].
Regarding structural asymmetries, the left STS has been shown to be longer than the right, but matures later, as seen in preterm newborn infants [55,56,58,183,184]. In addition, the right STS was found to be deeper than the left STS in a portion ventral to Heschl’s gyrus, called the STAP (Superior Temporal Asymmetrical Pit, [53,178,183]). This robust asymmetry is irrespective of age, handedness, and language lateralisation, suggesting a strong genetic influence [53,180,183]. Functional correspondence was found between its deepest point and location of the voice sensitive peak [178]. Several sulcus interruptions ‘plis de passage’ are more present in the left STS, probably resulting from stronger white matter fibres passing underneath the STS [185]. The aforementioned results highlight the STS’ implication in communication through the direct link of language perception and the indirect link with diverse social cognition tasks, which are crucial for complex language processing. This makes the STS and its asymmetric structure a promising key area in comparative studies in order to search for language prerequisites shared between primate relatives.
The STS was intensively studied in macaque monkeys and shown to be one main hub in the social interaction processing network, where it was mapped onto a fine-grain pattern of object, body, and face selectivity [186]. Indeed, its implication in a variety of social cognition tasks was demonstrated. This includes gaze following and joint attention [187] as well as facial movements in the upper STS [188]. In addition, the mid-STS in macaques was recently hypothesised to be equivalent of the human TPJ area, important for the theory of mind, because it was active for predicting social situations [189] and because macaques living in larger groups demonstrate an increased grey matter of this area [190]. Especially regarding communication, similar to humans, voice and face patches were also found in the monkey’s STS [178,191,192,193,194]. In fact, recently it was demonstrated that audiovisual integration occurred in a distinct region in the anterior fundus [195]. The Superior Temporal Asymmetrical Pit ‘STAP’ was not robustly shown to be present in chimpanzees, suggesting a human specific landmark of perisylvian organisation, which was related to human specific social cognition and communication [183]. According to results presented at conferences, structural rightward STS depth asymmetries were also found present in adult and juvenile baboons, within a portion that may overlap with the STAP in humans. Interestingly, preliminary results in baboons suggest that the strength of this rightward STS asymmetry varies according to social cognition proxies such as social group size and gestural communication’s right-handedness [196,197].
8. Inferior Parietal Lobe
The Inferior Parietal Lobe or Geschwind’s territory comprising the angular and the supramarginal gyrus was demonstrated as essential in language processing, connecting indirectly to primary language areas (such as the Planum Temporale or Broca’s area). For example, it was shown to be involved in episodic memory retrieval of words [198] or verbal working memory [199], but also in tool-use [200,201]. The Inferior Parietal Lobe is one of the latest to myelinate in development [202] and was related to the emergence of language in evolution [203] and in development [204]. The Inferior Parietal Lobe includes the Planum Parietale, which lies in the supramarginal gyrus, in the ascending portion of the Sylvian fissure, next to the Planum Temporale [62]. The Planum Parietale is implicated in dyslexia and communication disorders [69] and processing voice spectral information [205]. Adaptation during primate evolution of this area and its connectivity may provide the capacity of enhanced visual analysis of moving images that is important for tool handling and control [206,207,208]. However, due to its anatomical position, this region is a zone of convergence and integration of sensory and motor information via the fronto-parietal network [209]. In fact, the connection between Broca’s area and the Inferior Parietal Lobe is right-lateralised, in contrast to the left lateralisation of the Arcuate Fasciculus (see above). In addition, the Inferior Parietal Lobe inhabits mirror neurons that fire not only during motor execution, but also when observing actions performed by others, and might therefore lead to action understanding and language evolution [210,211].
Due to the aforementioned data, the Inferior Parietal Lobe is a key-region for investigating the lateralised links between actions, tool-use, and language evolution. A structural asymmetry of this area was found in the Planum Parietale. It showed a rightward asymmetry in both right-handed males and left-handed females, which was not correlated with the Planum Temporale asymmetry, indicating functional independence [62]. Another structural asymmetry was found for the parietal operculum, which constitutes the gyrus directly above the Planum Temporale as part of the supramarginal gyrus [212]. It shows a leftward asymmetry, especially for right-handers [213]. Regarding connectivity, tool-making skills elicited plastic remodeling of fronto-parietal white matter projections from the right Inferior Parietal Lobe into the right Broca’s area [201].
Because of the potential overlapping of brain circuits for tool-use and language, nonhuman primate brain studies have focussed on the Inferior Parietal Lobe. A potential interaction might lie in the semantic knowledge important for both language and tools to acquire the skill necessary to perform these actions [200,214,215]. In this view, modifications of circuits that subserve gestures and imitations may have paved the evolutionary way for language and tool-use in humans [215], with the Inferior Parietal Lobe in its heart. Recently, Cheng et al. [216] found that leftward rostral- and rightward caudal inferior parietal structural asymmetries connecting to several areas related to tool-use in humans [215] were similar in chimpanzees and humans but not present in macaques. This finding could be related to human-like leftward asymmetries of the parietal operculum, which were also reported in chimpanzees and related to tool-use but not non-tool-use motor actions [76]. To learn to use tools, humans need a certain level of body awareness to match variations in kinematic details with the desired outcome during practice. Similar awareness is required for other animals on the Mirror Self-Recognition test [217], which also some chimpanzee subjects pass. In fact, chimpanzee subjects that passed the test (to recognize themself in a mirror) also possessed a more right lateralised fronto-parietal connection, exactly as in humans during tool-making learning [82,201,218,219]. This rightward asymmetry of connectivity could be related to a human-like rightward asymmetry initially found for the Planum Parietale in apes [220], a finding which was replicated in chimpanzees, particularly in females in relation to handedness [221]. Future studies in nonhuman primates should continue investigating potential links in lateralisation of the Inferior Parietal Lobe and behaviour. It would help clarifying whether left or right lateralised brain circuits for tool-use paved the way for language brain circuits.
9. The Central Sulcus (CS)
The Central Sulcus (CS) is a major landmark in the brain, dividing the parietal from the frontal lobe and is one of the primary sulci developed in the brain [57]. It also divides the primary motor cortex from the primary somatosensory cortex, where topographic sensory and motor representations of human body parts are organised [222]. Within this topographic organisation, a morphological landmark of hand and finger representations has been documented across the dorsal-ventral plane of the CS, known as KNOB or motor hand area, which has an omega-like shape [223].
Regardless of the hand, the direction of handedness was found associated with contralateral asymmetries of the motor hand area (humans: [92,224,225,226]. In fact, the portion of the CS that delimits the motor hand area was found deeper in the hemisphere contralateral to the preferred hand of the subjects [92]. This feature seems, however, not to be related with language lateralisation. In fact, it has recently been demonstrated that the neural substrates of typical handedness measures and language brain organisation might be not related, but rather independent from each other [27,29,227].
Within an evolutionary framework, hemispheric specialisation and handedness have been historically considered unique to human language evolution [228,229]. However, many primates, such as baboons or chimpanzees, also present right-handed dominance for manipulative actions (wild subjects: [230]), and even stronger right-handed dominance for communicative gestures (captive subjects: [30]). Just as in humans, the nonhuman primates’ direction of handedness for object manipulation was found associated with contralateral asymmetries of the motor hand area within the Central Sulcus (Chimpanzees: [231,232]; Baboons: [233]; Capuchin monkeys: [234]; Squirrel monkeys: [235]). It is notable that the contralateral hand area effect in the CS was found exclusively for manipulative actions [233] but not for communicative gesture handedness. As mentioned in the previous section, handedness for gestural communication was exclusively found related to Broca’s homologue [106], suggesting a potential independent neural substrate of handedness and language organisation in evolution. Preliminary results in juvenile baboons reported a similar neuroanatomical manifestation of early handedness, highlighting that structural asymmetries in the Central Sulcus appear early in development with the emergence of handedness behaviour [236]. Interestingly, the motor hand area of the Central Sulcus was also shown to be related to more complex hierarchical organisation of behaviours as it was related to tool-use handedness in chimpanzees [97].
10. Conclusions
Within an evolutionary framework across primates, we compared data on inter-hemispheric structural asymmetries in most of the key brain structures that are known to be associated with hemispheric specialisation for language processing in humans.
The results of this comparative neuroanatomical approach conducted between humans and mostly chimpanzees, and to a lesser extent baboons, are quite straightforward and challenge the historical view that hemispheric specialisation is a human specific feature of language evolution [228,229]. It becomes clear that we share the structural lateralised patterns for most language-related regions with other primate species, and even new-born monkeys, especially for perisylvian regions, including the Planum Temporale and Broca’s area [24,69,71,73,74,95]. Although less documented in comparison to those two historical lateralised perisylvian regions, there is some evidence that other important regions of interest within the large human language network might share the same hemispheric structural lateralisation across primate species. According to rare recent comparative results, mostly conducted in chimpanzees and to a lesser extend in baboons, those shared features includes the leftward lateralised white matter connectivity tract between Broca’s area and the Planum Temporale-the Arcuate Fasciculus [134], leftward lateralised parietal operculum, rightward lateralised Planum Parietale and fronto-parietal projection [76,220,221], the rightward posterior section of the STS [196,197], as well as the presence of paracingulate sulcus in the left hemisphere [170]. These collective findings suggest an important shared biological ancestral encoding between Old World monkeys, great apes, and humans, that were initially considered as neuroanatomical landmarks of brain lateralisation for language. Their presence in non-linguistic primate species clearly states that these landmarks are not human- or language- specific.
Nevertheless, it remains unclear for which shared ‘domain-general’ cognitive function between human and nonhuman primate species, that is also at the heart of language processing in modern humans, these structural hemispheric asymmetry patterns are related to and have evolved. While reviewing the nonhuman primate literature that reported clear links between anatomical asymmetries for language area homologues and hand preferences for tool-use or for gestural communication [71,75,106], some functional roads become plausible. For instance, it is thus not excluded that both ‘syntactic’ hierarchical sequential processing (such as the one involved in tool making and use in great apes), and the intentional communication properties (shared with the communicative gestural system in primates) might thus constitute a functional candidate to have paved the way for such brain asymmetric organisation within the evolutionary framework of human language. We demonstrate, therefore, that nonhuman primates such as chimpanzees, and even more phylogenetic distant species such as baboons among the Old World monkeys, offer compelling comparative models for the evolution of human brains and behaviours. In particular, a sulci and region-of-interest approach seems fruitful for comparing brain structures. In the same vein, handedness measurements for different behaviours seem fruitful for linking the lateralised brain anatomy to lateralised behaviours. Interesting future work could now be in transferring this handedness knowledge to humans, to clarify, for example, whether communicative gesture handedness might be a marker for language lateralisation.
To interpret brain asymmetry and its evolution, we highlighted the importance of investigating the lateralisation of behaviours such as gestural communication, tool use, and bimanual coordination within a comparative approach [38]. Indeed, a blending of the two is necessary in order to gain a holistic view on how language lateralisation has evolved, which ultimately also provides an initial platform for its emergence.
Regarding specifically language evolution, we hypothesise that asymmetries for language areas may not have initially evolved for language, if language is seen as a cognitive module. Rather, each asymmetry could have evolved independently for independent cognitive functions to adapt to unknown environmental pressures. This could explain the unclear relationship between structural and functional asymmetries related to language areas. The structural asymmetry would here be a more ancient fossil of other cognitive specialisations on which the functional language asymmetry got piggybacked. The functional and structural specialisation of these structures may next have been important nests for developing intentional communicative behaviour in nonhuman primates, which later evolved to language processing in our species. In other words, ‘perisylvian language asymmetries’ are not specific to language but could rather be exaptations of pre-existing specialisations for other cognitive functions, which together make up what we call ‘language’. Therefore, ‘domain-general’ language-related brain architecture associated with intentional communicative or syntactic behaviours might rather be shared between humans, apes, and at least baboons of the Old World monkey family. Such an asymmetric brain organisation might have, thus, emerged from their common ancestor around 25 million years ago and later increased during hominin evolution.
Author Contributions
Conceptualization, Y.B. and A.M.; writing—original draft preparation, Y.B.; writing—review and editing, Y.B. and A.M.; supervision, A.M.; project administration, A.M.; funding acquisition, A.M. All authors have read and agreed to the published version of the manuscript.
Funding
The project has received funding from the European Research Council under the European Union’s Horizon 2020 research and innovation program grant agreement No. 716931-GESTIMAGE-ERC-2016-STG (P.I. Adrien Meguerditchian), as well as from grants ANR-16-CONV-0002 (ILCB), ANR-17-EURE-0029 (NeuroSchool) and the Excellence Initiative of Aix-Marseille University A*MIDEX AMX-19-IET-004 (NeuroMarseille).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Berwick, R.C.; Chomsky, N. Why Only Us: Language and Evolution; MIT Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Fedorenko, E. The role of domain-general cognitive control in language comprehension. Front. Psychol. 2014, 5, 335. [Google Scholar] [CrossRef] [PubMed]
- Fitch, W.T. The Evolution of Language; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Liebal, K.; Waller, B.M.; Slocombe, K.E.; Burrows, A.M. Primate Communication: A Multimodal Approach; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Fagot, J.; Boë, L.-J.; Berthomier, F.; Claidière, N.; Malassis, R.; Meguerditchian, A.; Rey, A.; Montant, M. The baboon: A model for the study of language evolution. J. Hum. Evol. 2019, 126, 39–50. [Google Scholar] [CrossRef]
- Geschwind, N. The Organization of Language and the Brain. Science 1970, 170, 940–944. [Google Scholar] [CrossRef]
- Toga, A.W.; Thompson, P.M. Mapping brain asymmetry. Nat. Rev. Neurosci. 2003, 4, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Biduła, S.P.; Kroliczak, G. Structural asymmetry of the insula is linked to the lateralization of gesture and language. Eur. J. Neurosci. 2015, 41, 1438–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, Y.; Loh, K.K.; Coulon, O.; Meguerditchian, A. The Arcuate Fasciculus and language origins: Disentangling existing conceptions that influence evolutionary accounts. Neurosci. Biobehav. Rev. 2022, 134, 104490. [Google Scholar] [CrossRef] [PubMed]
- Catani, M.; Mesulam, M. The arcuate fasciculus and the disconnection theme in language and aphasia: History and current state. Cortex 2008, 44, 953–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffau, H. The error of Broca: From the traditional localizationist concept to a connectomal anatomy of human brain. J. Chem. Neuroanat. 2018, 89, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Catani, M.; Dawson, M. Language Processing, Development and Evolution. Conn’s Transl. Neurosci. 2017, 679–692. [Google Scholar] [CrossRef]
- Keller, S.S.; Roberts, N.; García-Fiñana, M.; Mohammadi, S.; Ringelstein, E.B.; Knecht, S.; Deppe, M. Can the Language-dominant Hemisphere Be Predicted by Brain Anatomy? J. Cogn. Neurosci. 2010, 23, 2013–2029. [Google Scholar] [CrossRef] [PubMed]
- Amiez, C.; Sallet, J.; Hopkins, W.D.; Meguerditchian, A.; Hadj-Bouziane, F.; Ben Hamed, S.; Wilson, C.R.E.; Procyk, E.; Petrides, M. Sulcal organization in the medial frontal cortex provides insights into primate brain evolution. Nat. Commun. 2019, 10, 3437. [Google Scholar] [CrossRef] [Green Version]
- Gerrits, R.; Verhelst, H.; Dhollander, T.; Xiang, L.; Vingerhoets, G. Structural perisylvian asymmetry in naturally occurring atypical language dominance. Anat. Embryol. 2021, 227, 573–586. [Google Scholar] [CrossRef] [PubMed]
- Tzourio-Mazoyer, N.; Crivello, F.; Mazoyer, B. Is the planum temporale surface area a marker of hemispheric or regional language lateralization? Anat. Embryol. 2018, 223, 1217–1228. [Google Scholar] [CrossRef]
- Dehaene-Lambertz, G.; Dehaene, S.; Hertz-Pannier, L. Functional Neuroimaging of Speech Perception in Infants. Science 2002, 298, 2013–2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catani, M.; Allin, M.P.G.; Husain, M.; Pugliese, L.; Mesulam, M.M.; Murray, R.M.; Jones, D.K. Symmetries in human brain language pathways correlate with verbal recall. Proc. Natl. Acad. Sci. USA 2007, 104, 17163–17168. [Google Scholar] [CrossRef] [Green Version]
- Darwin, C. The Descent of Man and Selection in Relation to Sex; Princeton University Press: Princeton, NJ, USA, 1981; Volume 1, [Originally Work Published 1871]. [Google Scholar]
- Hopkins, W.D.; Misiura, M.; Pope, S.M.; Latash, E.M. Behavioral and brain asymmetries in primates: A preliminary evaluation of two evolutionary hypotheses. Ann. N. Y. Acad. Sci. 2015, 1359, 65–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirier, C.; Ben Hamed, S.; Garcia-Saldivar, P.; Kwok, S.C.; Meguerditchian, A.; Merchant, H.; Rogers, J.; Wells, S.; Fox, A.S. Beyond MRI: On the scientific value of combining non-human primate neuroimaging with metadata. NeuroImage 2021, 228, 117679. [Google Scholar] [CrossRef] [PubMed]
- Milham, M.; Petkov, C.I.; Margulies, D.S.; Schroeder, C.E.; Basso, M.A.; Belin, P.; Fair, D.A.; Fox, A.; Kastner, S.; Mars, R.; et al. Accelerating the Evolution of Nonhuman Primate Neuroimaging. Neuron 2020, 105, 600–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, W.D.; Taglialatela, J.P.; Meguerditchian, A.; Nir, T.; Schenker, N.M.; Sherwood, C.C. Gray matter asymmetries in chimpanzees as revealed by voxel-based morphometry. NeuroImage 2008, 42, 491–497. [Google Scholar] [CrossRef] [Green Version]
- Marie, D.; Roth, M.; Lacoste, R.; Nazarian, B.; Bertello, A.; Anton, J.-L.; Hopkins, W.D.; Margiotoudi, K.; Love, S.A.; Meguerditchian, A. Left Brain Asymmetry of the Planum Temporale in a Nonhominid Primate: Redefining the Origin of Brain Specialization for Language. Cereb. Cortex 2018, 28, 1808–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knecht, S.; Dräger, B.; Deppe, M.; Bobe, L.; Lohmann, H.; Flöel, A.; Ringelstein, E.-B.; Henningsen, H. Handedness and hemispheric language dominance in healthy humans. Brain 2000, 123, 2512–2518. [Google Scholar] [CrossRef] [Green Version]
- Amunts, K.; Jäncke, L.; Mohlberg, H.; Steinmetz, H.; Zilles, K. Interhemispheric asymmetry of the human motor cortex related to handedness and gender. Neuropsychologia 2000, 38, 304–312. [Google Scholar] [CrossRef]
- Groen, M.A.; Whitehouse, A.J.O.; Badcock, N.A.; Bishop, D.V.M. Associations between Handedness and Cerebral Lateralisation for Language: A Comparison of Three Measures in Children. PLoS ONE 2013, 8, e64876. [Google Scholar] [CrossRef] [Green Version]
- Mazoyer, B.; Zago, L.; Jobard, G.; Crivello, F.; Joliot, M.; Perchey, G.; Mellet, E.; Petit, L.; Tzourio-Mazoyer, N. Gaussian Mixture Modeling of Hemispheric Lateralization for Language in a Large Sample of Healthy Individuals Balanced for Handedness. PLoS ONE 2014, 9, e101165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ocklenburg, S.; Beste, C.; Arning, L.; Peterburs, J.; Güntürkün, O. The ontogenesis of language lateralization and its relation to handedness. Neurosci. Biobehav. Rev. 2014, 43, 191–198. [Google Scholar] [CrossRef]
- Meguerditchian, A.; Vauclair, J.; Hopkins, W.D. On the origins of human handedness and language: A comparative review of hand preferences for bimanual coordinated actions and gestural communication in nonhuman primates. Dev. Psychobiol. 2013, 55, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Hewes, G.W.; Andrew, R.J.; Carini, L.; Choe, H.; Gardner, R.A.; Kortlandt, A.; Krantz, G.S.; McBride, G.; Nottebohm, F.; Pfeiffer, J.; et al. Primate Communication and the Gestural Origin of Language [and Comments and Reply]. Curr. Anthr. 1973, 14, 5–24. [Google Scholar] [CrossRef]
- Tomasello, M. Origins of Human Communication; The MIT Press: Cambridge, UK, 2008. [Google Scholar]
- Meguerditchian, A.; Vauclair, J. Communicative Signaling, Lateralization and Brain Substrate in Nonhuman Primates: Toward a Gestural or a Multimodal Origin of Language? Hum. Mente J. Philos. Stud. 2014, 7, 135–160. [Google Scholar]
- Molesti, S.; Meguerditchian, A.; Bourjade, M. Gestural communication in olive baboons [Papio anubis]: Repertoire and intentionality. Anim. Cogn. 2019, 23, 19–40. [Google Scholar] [CrossRef]
- Greenfield, P.M. Language, tools and brain: The ontogeny and phylogeny of hierarchically organized sequential behavior. Behav. Brain Sci. 1991, 14, 531–551. [Google Scholar] [CrossRef] [Green Version]
- Corballis, M.C. From mouth to hand: Gesture, speech and the evolution of right- handedness. Brain Sci. 2004, 26, 199–260. [Google Scholar] [CrossRef] [PubMed]
- Stout, D.; Toth, N.; Schick, K.; Chaminade, T. Neural correlates of Early Stone Age toolmaking: Technology, language and cognition in human evolution. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 1939–1949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forrester, G.S.; Quaresmini, C.; Leavens, D.; Mareschal, D.; Thomas, M.S. Human handedness: An inherited evolutionary trait. Behav. Brain Res. 2013, 237, 200–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higuchi, S.; Chaminade, T.; Imamizu, H.; Kawato, M. Shared neural correlates for language and tool use in Broca’s area. NeuroReport 2009, 20, 1376–1381. [Google Scholar] [CrossRef]
- Thibault, S.; Py, R.; Gervasi, A.M.; Salemme, R.; Koun, E.; Lövden, M.; Boulenger, V.; Roy, A.C.; Brozzoli, C. Tool use and language share syntactic processes and neural patterns in the basal ganglia. Science 2021, 374, eabe0874. [Google Scholar] [CrossRef]
- Galaburda, A.M.; LeMay, M.; Kemper, T.L.; Geschwind, N. Right-Left Asymmetries in the Brain. Science 1978, 199, 852–856. [Google Scholar] [CrossRef]
- Mesulam, M.M. From sensation to cognition. Brain 1998, 121, 1013–1052. [Google Scholar] [CrossRef]
- Wernicke, C. Der Aphasische Symptomencomplex: Eine Psychologische Studie auf Anatomischer Basis; Cohn: Breslau, Poland, 1874. [Google Scholar]
- Dronkers, N.F.; Wilkins, D.P.; Van Valin, R.D.; Redfern, B.B.; Jaeger, J.J. Lesion analysis of the brain areas involved in language comprehension. Cognition 2004, 92, 145–177. [Google Scholar] [CrossRef] [Green Version]
- Borovsky, A.; Saygin, A.P.; Bates, E.; Dronkers, N. Lesion correlates of conversational speech production deficits. Neuropsychologia 2007, 45, 2525–2533. [Google Scholar] [CrossRef] [Green Version]
- Shapleske, J.; Rossell, S.; Woodruff, P.; David, A. The planum temporale: A systematic, quantitative review of its structural, functional and clinical significance. Brain Res. Rev. 1999, 29, 26–49. [Google Scholar] [CrossRef]
- Vigneau, M.; Beaucousin, V.; Hervé, P.; Duffau, H.; Crivello, F.; Houdé, O.; Mazoyer, B.; Tzourio-Mazoyer, N. Meta-analyzing left hemisphere language areas: Phonology, semantics, and sentence processing. NeuroImage 2006, 30, 1414–1432. [Google Scholar] [CrossRef] [PubMed]
- Dehaene-Lambertz, G.; Montavont, A.; Jobert, A.; Allirol, L.; Dubois, J.; Hertz-Pannier, L.; Dehaene, S. Language or music, mother or Mozart? Structural and environmental influences on infants’ language networks. Brain Lang. 2010, 114, 53–65. [Google Scholar] [CrossRef]
- Mahmoudzadeh, M.; Dehaene-Lambertz, G.; Fournier, M.; Kongolo, G.; Goudjil, S.; Dubois, J.; Grebe, R.; Wallois, F. Syllabic discrimination in premature human infants prior to complete formation of cortical layers. Proc. Natl. Acad. Sci. USA 2013, 110, 4846–4851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geschwind, N.; Levitsky, W. Human Brain: Left-Right Asymmetries in Temporal Speech Region. Science 1968, 161, 186–187. [Google Scholar] [CrossRef]
- Wada, J.A.; Clarke, R.; Hamm, A. Cerebral Hemispheric Asymmetry in Humans. Arch. Neurol. 1975, 32, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Witelson, S.F.; Pallie, W. Left hemisphere Spezialisation for Language in the Newborn. Brain 1973, 96, 641–646. [Google Scholar] [CrossRef]
- Glasel, H.; Leroy, F.; Dubois, J.; Hertz-Pannier, L.; Mangin, J.; Dehaene-Lambertz, G. A robust cerebral asymmetry in the infant brain: The rightward superior temporal sulcus. NeuroImage 2011, 58, 716–723. [Google Scholar] [CrossRef]
- Hill, J.; Inder, T.; Neil, J.; Dierker, D.; Harwell, J.; Van Essen, D. Similar patterns of cortical expansion during human development and evolution. Proc. Natl. Acad. Sci. USA 2010, 107, 13135–13140. [Google Scholar] [CrossRef] [Green Version]
- Dubois, J.; Benders, M.; Cachia, A.; Lazeyras, F.; Leuchter, R.H.-V.; Sizonenko, S.V.; Borradori-Tolsa, C.; Mangin, J.F.; Hüppi, P.S. Mapping the Early Cortical Folding Process in the Preterm Newborn Brain. Cereb. Cortex 2008, 18, 1444–1454. [Google Scholar] [CrossRef] [Green Version]
- Dubois, J.; Benders, M.; Lazeyras, F.; Borradori-Tolsa, C.; Leuchter, R.H.-V.; Mangin, J.; Hüppi, P. Structural asymmetries of perisylvian regions in the preterm newborn. NeuroImage 2010, 52, 32–42. [Google Scholar] [CrossRef]
- Chi, J.G.; Dooling, E.C.; Gilles, F.H. Gyral development of the human brain. Ann. Neurol. 1977, 1, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Chi, J.G.; Dooling, E.C.; Gilles, F.H. Left-Right Asymmetries of the Temporal Speech Areas of the Human Fetus. Arch. Neurol. 1977, 34, 346–348. [Google Scholar] [CrossRef] [PubMed]
- Josse, G.; Hervé, P.-Y.; Crivello, F.; Mazoyer, B.; Tzourio-Mazoyer, N. Hemispheric specialization for language: Brain volume matters. Brain Res. 2006, 1068, 184–193. [Google Scholar] [CrossRef]
- Gauger, L.M.; Lombardino, L.J.; Leonard, C.M. Brain Morphology in Children with Specific Language Impairment. J. Speech Lang. Hear. Res. 1997, 40, 1272–1284. [Google Scholar] [CrossRef] [PubMed]
- Altarelli, I.; Leroy, F.; Monzalvo, K.; Fluss, J.; Billard, C.; Dehaene-Lambertz, G.; Galaburda, A.M.; Ramus, F. Planum temporale asymmetry in developmental dyslexia: Revisiting an old question. Hum. Brain Mapp. 2014, 35, 5717–5735. [Google Scholar] [CrossRef]
- Jäncke, L.; Schlaug, G.; Huang, Y.; Steinmetz, H. Asymmetry of the planum parietale. NeuroReport 1994, 5, 1161–1163. [Google Scholar] [CrossRef]
- Dorsaint-Pierre, R.; Penhune, V.B.; Watkins, K.; Neelin, P.; Lerch, J.P.; Bouffard, M.; Zatorre, R.J. Asymmetries of the planum temporale and Heschl’s gyrus: Relationship to language lateralization. Brain 2006, 129, 1164–1176. [Google Scholar] [CrossRef] [Green Version]
- Eckert, M.A.; Leonard, C.M.; Possing, E.T.; Binder, J.R. Uncoupled leftward asymmetries for planum morphology and functional language processing. Brain Lang. 2006, 98, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Greve, D.N.; Van der Haegen, L.; Cai, Q.; Stufflebeam, S.; Sabuncu, M.R.; Fischl, B.; Brysbaert, M. A Surface-based Analysis of Language Lateralization and Cortical Asymmetry. J. Cogn. Neurosci. 2013, 25, 1477–1492. [Google Scholar] [CrossRef]
- Kolinsky, R.; Morais, J.; Cohen, L.; Dehaene-Lambertz, G.; Dehaene, S. The impact of literacy on the language brain areas. Rev. Neuropsychol. 2014, 6, 173–181. [Google Scholar] [CrossRef]
- Tzourio-Mazoyer, N.; Mazoyer, B. Variations of planum temporale asymmetries with Heschl’s Gyri duplications and association with cognitive abilities: MRI investigation of 428 healthy volunteers. Anat. Embryol. 2017, 222, 2711–2726. [Google Scholar] [CrossRef] [PubMed]
- Ocklenburg, S.; Friedrich, P.; Fraenz, C.; Schlüter, C.; Beste, C.; Güntürkün, O.; Genç, E. Neurite architecture of the planum temporale predicts neurophysiological processing of auditory speech. Sci. Adv. 2018, 4, eaar6830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gannon, P.J.; Holloway, R.L.; Broadfield, D.C.; Braun, A.R. Asymmetry of Chimpanzee Planum Temporale: Humanlike Pattern of Wernicke’s Brain Language Area Homolog. Science 1998, 279, 220–222. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.D.; Marino, L.; Rilling, J.K.; MacGregor, L.A. Planum temporale asymmetries in great apes as revealed by magnetic resonance imaging [MRI]. NeuroReport 1998, 9, 2913–2918. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.D.; Nir, T.M. Planum temporale surface area and grey matter asymmetries in chimpanzees [Pan troglodytes]: The effect of handedness and comparison with findings in humans. Behav. Brain Res. 2010, 208, 436–443. [Google Scholar] [CrossRef] [Green Version]
- Cantalupo, C.; Pilcher, D.L.; Hopkins, W.D. Are planum temporale and sylvian fissure asymmetries directly related? A MRI study in great apes. Neuropsychologia 2003, 41, 1975–1981. [Google Scholar] [CrossRef]
- Becker, Y.; Sein, J.; Velly, L.; Giacomino, L.; Renaud, L.; Lacoste, R.; Anton, J.-L.; Nazarian, B.; Berne, C.; Meguerditchian, A. Early Left-Planum Temporale Asymmetry in newborn monkeys [Papio anubis]: A longitudinal structural MRI study at two stages of development. NeuroImage 2021, 227, e117575. [Google Scholar] [CrossRef]
- Becker, Y.; Phelipon, R.; Sein, J.; Velly, L.; Renaud, L.; Meguerditchian, A. Planum temporale grey matter volume asymmetries in newborn monkeys [Papio anubis]. Brain Struct. Funct. 2022, 227, 463–468. [Google Scholar] [CrossRef]
- Meguerditchian, A.; Gardner, M.J.; Schapiro, S.J.; Hopkins, W.D. The sound of one-hand clapping: Handedness and perisylvian neural correlates of a communicative gesture in chimpanzees. Proc. R. Soc. B Boil. Sci. 2012, 279, 1959–1966. [Google Scholar] [CrossRef] [Green Version]
- Gilissen, E.P.; Hopkins, W.D. Asymmetries of the Parietal Operculum in Chimpanzees [Pan troglodytes] in Relation to Handedness for Tool Use. Cereb. Cortex 2013, 23, 411–422. [Google Scholar] [CrossRef] [Green Version]
- Broca, P. Remarques sur le siège de la Faculté du Langage Articulé, Suivies d’une Observation d’aphémie [perte de la parole]; Bulletin et Memoires de la Societe Anatomique de Paris: Paris, France, 1861; Volume 6, pp. 330–357. [Google Scholar]
- Hickok, G.; Poeppel, D. The cortical organization of speech processing. Nat. Rev. Neurosci. 2007, 8, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Friederici, A.D. Language in Our Brain: The Origins of a Uniquely Human Capacity; The MIT Press: Cambridge, MA, USA, 2017. [Google Scholar]
- Gentilucci, M.; Volta, R.D. Spoken Language and arm Gestures are Controlled by the same Motor Control System. Q. J. Exp. Psychol. 2008, 61, 944–957. [Google Scholar] [CrossRef] [PubMed]
- Koechlin, E.; Jubault, T. Broca’s Area and the Hierarchical Organization of Human Behavior. Neuron 2006, 50, 963–974. [Google Scholar] [CrossRef] [Green Version]
- Stout, D.; Hecht, E.E. Evolutionary neuroscience of cumulative culture. Proc. Natl. Acad. Sci. USA 2017, 114, 7861–7868. [Google Scholar] [CrossRef] [Green Version]
- Emmorey, K.; Grabowski, T.; McCullough, S.; Damasio, H.; Ponto, L.; Hichwa, R.; Bellugi, U. Motor-iconicity of sign language does not alter the neural systems underlying tool and action naming. Brain Lang. 2004, 89, 27–37. [Google Scholar] [CrossRef]
- Campbell, R.; Macsweeney, M.; Waters, D. Sign Language and the Brain: A Review. J. Deaf Stud. Deaf Educ. 2007, 13, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Imada, T.; Zhang, Y.; Cheour, M.; Taulu, S.; Ahonen, A.; Kuhl, P.K. Infant speech perception activates Broca’s area: A developmental magnetoencephalography study. NeuroReport 2006, 17, 957–962. [Google Scholar] [CrossRef]
- Dehaene-Lambertz, G.; Hertz-Pannier, L.; Dubois, J.; Mériaux, S.; Roche, A.; Sigman, M.; Dehaene, S. Functional organization of perisylvian activation during presentation of sentences in preverbal infants. Proc. Natl. Acad. Sci. USA 2006, 103, 14240–14245. [Google Scholar] [CrossRef] [Green Version]
- Keller, S.S.; Crow, T.; Foundas, A.; Amunts, K.; Roberts, N. Broca’s area: Nomenclature, anatomy, typology and asymmetry. Brain Lang. 2009, 109, 29–48. [Google Scholar] [CrossRef]
- Sprung-Much, T.; Eichert, N.; Nolan, E.; Petrides, M. Broca’s area and the search for anatomical asymmetry: Commentary and perspectives. Anat. Embryol. 2022, 227, 441–449. [Google Scholar] [CrossRef]
- Keller, S.S.; Highley, J.R.; Garcia-Finana, M.; Sluming, V.; Rezaie, R.; Roberts, N. Sulcal variability, stereological measurement and asymmetry of Broca’s area on MR images. J. Anat. 2007, 211, 534–555. [Google Scholar] [CrossRef] [PubMed]
- Scheibel, A.B. A dendritic correlate of human speech. In Cerebral Dominance: The Biological Foundations; Harvard University Press: Cambridge, MA, USA, 1984; pp. 43–52. [Google Scholar]
- Amunts, K.; Schleicher, A.; Mohlberg, H.; Uylings, H.B.; Zilles, K. Broca’s region revisited: Cytoarchitecture and intersubject variability. J. Comp. Neurol. 1999, 412, 319–341. [Google Scholar] [CrossRef]
- Amunts, K.; Schleicher, A.; Ditterich, A.; Zilles, K. Broca’s region: Cytoarchitectonic asymmetry and developmental changes. J. Comp. Neurol. 2003, 465, 72–89. [Google Scholar] [CrossRef]
- Kurth, F.; Cherbuin, N.; Luders, E. Speaking of aging: Changes in gray matter asymmetry in Broca’s area in later adulthood. Cortex 2020, 129, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Simonds, R.J.; Scheibel, A.B. The postnatal development of the motor speech area: A preliminary study. Brain Lang. 1989, 37, 42–58. [Google Scholar] [CrossRef]
- Cantalupo, C.; Hopkins, W.D. Asymmetric Broca’s area in great apes. Nature 2001, 414, 505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, S.S.; Deppe, M.; Herbin, M.; Gilissen, E. Variability and asymmetry of the sulcal contours defining Broca’s area homologue in the chimpanzee brain. J. Comp. Neurol. 2012, 520, 1165–1180. [Google Scholar] [CrossRef]
- Hopkins, W.D. Motor and Communicative Correlates of the Inferior Frontal Gyrus [Broca’s Area] in Chimpanzees. In Origins of Human Language: Continuities and Splits with Nonhuman Primates; Boë, L.-J., Fagot, J., Perrier, P., Schwartz, J.-L., Eds.; Peter Lang: Oxford, UK, 2017; pp. 153–186. [Google Scholar]
- Schenker-Ahmed, N.; Hopkins, W.D.; Spocter, M.; Garrison, A.R.; Stimpson, C.D.; Erwin, J.M.; Hof, P.R.; Sherwood, C.C. Broca’s Area Homologue in Chimpanzees [Pan troglodytes]: Probabilistic Mapping, Asymmetry, and Comparison to Humans. Cereb. Cortex 2010, 20, 730–742. [Google Scholar] [CrossRef] [Green Version]
- Graïc, J.-M.; Peruffo, A.; Corain, L.; Centelleghe, C.; Granato, A.; Zanellato, E.; Cozzi, B. Asymmetry in the Cytoarchitecture of the Area 44 Homolog of the Brain of the Chimpanzee Pan troglodytes. Front. Neuroanat. 2020, 14. [Google Scholar] [CrossRef]
- Petrides, M. Lateral prefrontal cortex: Architectonic and functional organization. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 781–795. [Google Scholar] [CrossRef] [Green Version]
- Petrides, M.; Cadoret, G.; Mackey, S. Orofacial somatomotor responses in the macaque monkey homologue of Broca’s area. Nature 2005, 435, 1235–1238. [Google Scholar] [CrossRef] [PubMed]
- Belmalih, A.; Borra, E.; Contini, M.; Gerbella, M.; Rozzi, S.; Luppino, G. Multimodal architectonic subdivision of the rostral part [area F5] of the macaque ventral premotor cortex. J. Comp. Neurol. 2009, 512, 183–217. [Google Scholar] [CrossRef] [PubMed]
- Hage, S.R.; Nieder, A. Single neurons in monkey prefrontal cortex encode volitional initiation of vocalizations. Nat. Commun. 2013, 4, 2409. [Google Scholar] [CrossRef] [PubMed]
- Taglialatela, J.P.; Russell, J.L.; Schaeffer, J.A.; Hopkins, W.D. Communicative Signaling Activates ‘Broca’s’ Homolog in Chimpanzees. Curr. Biol. 2008, 18, 343–348. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, W.D.; Meguerditchian, A.; Coulon, O.; Misiura, M.; Pope, S.; Mareno, M.C.; Schapiro, S.J. Motor skill for tool-use is associated with asymmetries in Broca’s area and the motor hand area of the precentral gyrus in chimpanzees [Pan troglodytes]. Behav. Brain Res. 2017, 318, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Becker, Y.; Claidière, N.; Margiotoudi, K.; Marie, D.; Roth, M.; Nazarian, B.; Anton, J.-L.; Coulon, O.; Meguerditchian, A. Broca’s cerebral asymmetry reflects gestural communication’s lateralisation in monkeys [Papio anubis]. eLife 2022, 11, e70521. [Google Scholar] [CrossRef]
- Catani, M.; Schotten, M.T. Atlas of Human Brain Connections; OUP Oxford: Oxford, UK, 2012. [Google Scholar]
- Metellus, P.; Boussen, S.; Guye, M.; Trebuchon, A. Successful Insular Glioma Removal in a Deaf Signer Patient during an Awake Craniotomy Procedure. World Neurosurg. 2017, 98, 883-e1. [Google Scholar] [CrossRef]
- Dubois, J.; Poupon, C.; Thirion, B.; Simonnet, H.; Kulikova, S.; Leroy, F.; Hertz-Pannier, L.; Dehaene-Lambertz, G. Exploring the Early Organization and Maturation of Linguistic Pathways in the Human Infant Brain. Cereb. Cortex 2016, 26, 2283–2298. [Google Scholar] [CrossRef]
- Brauer, J.; Anwander, A.; Perani, D.; Friederici, A.D. Dorsal and ventral pathways in language development. Brain Lang. 2013, 127, 289–295. [Google Scholar] [CrossRef] [Green Version]
- Brauer, J.; Anwander, A.; Friederici, A.D. Neuroanatomical Prerequisites for Language Functions in the Maturing Brain. Cereb. Cortex 2011, 21, 459–466. [Google Scholar] [CrossRef]
- Dubois, J.; Hertz-Pannier, L.; Dehaene-Lambertz, G.; Cointepas, Y.; Le Bihan, D. Assessment of the early organization and maturation of infants’ cerebral white matter fiber bundles: A feasibility study using quantitative diffusion tensor imaging and tractography. NeuroImage 2006, 30, 1121–1132. [Google Scholar] [CrossRef] [PubMed]
- Dubois, J.; Hertz-Pannier, L.; Cachia, A.; Mangin, J.F.; Le Bihan, D.; Dehaene-Lambertz, G. Structural Asymmetries in the Infant Language and Sensori-Motor Networks. Cereb. Cortex 2009, 19, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Perani, D.; Saccuman, M.C.; Scifo, P.; Anwander, A.; Spada, D.; Baldoli, C.; Poloniato, A.; Lohmann, G.; Friederici, A.D. Neural language networks at birth. Proc. Natl. Acad. Sci. USA 2011, 108, 16056–16061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friederici, A.D. Language Development and the Ontogeny of the Dorsal Pathway. Front. Evol. Neurosci. 2012, 4, 3. [Google Scholar] [CrossRef] [Green Version]
- Büchel, C.; Raedler, T.; Sommer, M.; Sach, M.; Weiller, C.; Koch, M. White Matter Asymmetry in the Human Brain: A Diffusion Tensor MRI Study. Cereb. Cortex 2004, 14, 945–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nucifora, P.G.P.; Verma, R.; Melhem, E.R.; Gur, R.E.; Gur, R.C. Leftward asymmetry in relative fiber density of the arcuate fasciculus. NeuroReport 2005, 16, 791–794. [Google Scholar] [CrossRef] [Green Version]
- Powell, H.R.; Parker, G.J.; Alexander, D.C.; Symms, M.R.; Boulby, P.A.; Wheeler-Kingshott, C.A.; Barker, G.J.; Noppeney, U.; Koepp, M.J.; Duncan, J.S. Hemispheric asymmetries in language-related pathways: A combined functional MRI and tractography study. NeuroImage 2006, 32, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Takaya, S.; Kuperberg, G.R.; Liu, H.; Greve, D.N.; Makris, N.; Stufflebeam, S.M. Asymmetric projections of the arcuate fasciculus to the temporal cortex underlie lateralized language function in the human brain. Front. Neuroanat. 2015, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Thiebaut de Schotten, M.; Ffytche, D.H.; Bizzi, A.; Dell’Acqua, F.; Allin, M.; Walshe, M.; Murray, R.; Williams, S.C.; Murphy, D.G.M.; Catani, M. Atlasing location, asymmetry and inter-subject variability of white matter tracts in the human brain with MR diffusion tractography. NeuroImage 2011, 54, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Balériaux, D.; Kavec, M.; Metens, T.; Absil, J.; Denolin, V.; Pardou, A.; Avni, F.; Van Bogaert, P.; Aeby, A. Structural asymmetries in motor and language networks in a population of healthy preterm neonates at term equivalent age: A diffusion tensor imaging and probabilistic tractography study. NeuroImage 2010, 51, 783–788. [Google Scholar] [CrossRef]
- Song, J.W.; Mitchell, P.D.; Kolasinski, J.; Ellen Grant, P.; Galaburda, A.M.; Takahashi, E. Asymmetry of White Matter Pathways in Developing Human Brains. Cereb. Cortex 2015, 25, 2883–2893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebel, C.; Beaulieu, C. Lateralization of the arcuate fasciculus from childhood to adulthood and its relation to cognitive abilities in children. Hum. Brain Mapp. 2009, 30, 3563–3573. [Google Scholar] [CrossRef] [PubMed]
- López-Barroso, D.; Catani, M.; Ripollés, P.; Dell’Acqua, F.; Rodríguez-Fornells, A.; de Diego-Balaguer, R. Word learning is mediated by the left arcuate fasciculus. Proc. Natl. Acad. Sci. USA 2013, 110, 13168–13173. [Google Scholar] [CrossRef] [Green Version]
- Salvan, P.; Tournier, J.D.; Batalle, D.; Falconer, S.; Chew, A.; Kennea, N.; Aljabar, P.; Dehaene-Lambertz, G.; Arichi, T.; Edwards, A.D.; et al. Language ability in preterm children is associated with arcuate fasciculi microstructure at term. Hum. Brain Mapp. 2017, 38, 3836–3847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Propper, R.E.; O’Donnell, L.J.; Whalen, S.; Tie, Y.; Norton, I.H.; Suarez, R.O.; Zollei, L.; Radmanesh, A.; Golby, A.J. A combined fMRI and DTI examination of functional language lateralization and arcuate fasciculus structure: Effects of degree versus direction of hand preference. Brain Cogn. 2010, 73, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Fan, Y.; Zou, Q.; Wang, J.; Gao, J.-H.; Niu, Z. Temporal Reliability and Lateralization of the Resting-State Language Network. PLoS ONE 2014, 9, e85880. [Google Scholar] [CrossRef] [PubMed]
- Silva, G.; Citterio, A. Hemispheric asymmetries in dorsal language pathway white-matter tracts: A magnetic resonance imaging tractography and functional magnetic resonance imaging study. Neuroradiol. J. 2017, 30, 470–476. [Google Scholar] [CrossRef]
- Verhelst, H.; Dhollander, T.; Gerrits, R.; Vingerhoets, G. Fibre-specific laterality of white matter in left and right language dominant people. NeuroImage 2021, 230, 117812. [Google Scholar] [CrossRef]
- Schmahmann, J.D.; Pandya, D.N. Fiber Pathways of the Brain; Oxford University Press: Oxford, UK, 2006. [Google Scholar]
- Petrides, M.; Pandya, D.N. Distinct Parietal and Temporal Pathways to the Homologues of Broca’s Area in the Monkey. PLoS Biol. 2009, 7, e1000170. [Google Scholar] [CrossRef] [Green Version]
- Petrides, M. Neuroanatomy of Language Regions of the Human Brain, 1st ed.; Elsevier/AP, Academic Press is an imprint of Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Rilling, J.K.; Glasser, M.F.; Preuss, T.M.; Ma, X.; Zhao, T.; Hu, X.; Behrens, T.E.J. The evolution of the arcuate fasciculus revealed with comparative DTI. Nat. Neurosci. 2008, 11, 426–428. [Google Scholar] [CrossRef]
- Rilling, J.K.; Glasser, M.F.; Jbabdi, S.; Andersson, J.; Preuss, T.M. Continuity, Divergence, and the Evolution of Brain Language Pathways. Front. Evol. Neurosci. 2012, 3, 11. [Google Scholar] [CrossRef] [Green Version]
- Frey, S.; Mackey, S.; Petrides, M. Cortico-cortical connections of areas 44 and 45B in the macaque monkey. Brain Lang. 2014, 131, 36–55. [Google Scholar] [CrossRef]
- Eichert, N.; Verhagen, L.; Folloni, D.; Jbabdi, S.; Khrapitchev, A.A.; Sibson, N.R.; Mantini, D.; Sallet, J.; Mars, R. What is special about the human arcuate fasciculus? Lateralization, projections, and expansion. Cortex 2019, 118, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Barrett, R.L.C.; Dawson, M.; Dyrby, T.B.; Krug, K.; Ptito, M.; D’Arceuil, H.; Croxson, P.L.; Johnson, P.J.; Howells, H.; Forkel, S.; et al. Differences in Frontal Network Anatomy Across Primate Species. J. Neurosci. 2020, 40, 2094–2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balezeau, F.; Wilson, B.; Gallardo, G.; Dick, F.; Hopkins, W.; Anwander, A.; Friederici, A.D.; Griffiths, T.D.; Petkov, C.I. Primate auditory prototype in the evolution of the arcuate fasciculus. Nat. Neurosci. 2020, 23, 611–614. [Google Scholar] [CrossRef] [PubMed]
- Rocchi, F.; Oya, H.; Balezeau, F.; Billig, A.; Kocsis, Z.; Jenison, R.L.; Nourski, K.; Kovach, C.; Steinschneider, M.; Kikuchi, Y.; et al. Common fronto-temporal effective connectivity in humans and monkeys. Neuron 2021, 109, 852–868.e8. [Google Scholar] [CrossRef]
- Dronkers, N.F. A new brain region for coordinating speech articulation. Nature 1996, 384, 159–161. [Google Scholar] [CrossRef]
- Oh, A.; Duerden, E.G.; Pang, E.W. The role of the insula in speech and language processing. Brain Lang. 2014, 135, 96–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bamiou, D.-E.; Musiek, F.E.; Luxon, L.M. The insula [Island of Reil] and its role in auditory processing: Literature review. Brain Res. Rev. 2003, 42, 143–154. [Google Scholar] [CrossRef]
- Ackermann, H.; Riecker, A. The contribution of the insula to motor aspects of speech production: A review and a hypothesis. Brain Lang. 2004, 89, 320–328. [Google Scholar] [CrossRef]
- Friederici, A.D. The neural basis for human syntax: Broca’s area and beyond. Curr. Opin. Behav. Sci. 2018, 21, 88–92. [Google Scholar] [CrossRef]
- Zaccarella, E.; Friederici, A.D. Merge in the Human Brain: A Sub-Region Based Functional Investigation in the Left Pars Opercularis. Front. Psychol. 2015, 6, 1818. [Google Scholar] [CrossRef] [Green Version]
- Lœvenbruck, H.; Baciu, M.; Segebarth, C.; Abry, C. The left inferior frontal gyrus under focus: An fMRI study of the production of deixis via syntactic extraction and prosodic focus. J. Neurolinguistics 2005, 18, 237–258. [Google Scholar] [CrossRef] [Green Version]
- Lœvenbruck, H.; Vilain, C.; Dohen, M. From gestural pointing to vocal pointing in the brain. Rev. Fr. Linguist. Appl. 2008, 13, 23–33. [Google Scholar] [CrossRef]
- Chiarello, C.; Vazquez, D.; Felton, A.; Leonard, C.M. Structural asymmetry of anterior insula: Behavioral correlates and individual differences. Brain Lang. 2013, 126, 109–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Economo, C.V. Eine neue art spezialzellen des lobus cinguli und lobus insulae. Z. Die Gesamte Neurol. Und Psychiatr. 1926, 100, 706–712. [Google Scholar] [CrossRef]
- Evrard, H.C. Von Economo and fork neurons in the monkey insula, implications for evolution of cognition. Curr. Opin. Behav. Sci. 2018, 21, 182–190. [Google Scholar] [CrossRef]
- Bauernfeind, A.L.; de Sousa, A.A.; Avasthi, T.; Dobson, S.D.; Raghanti, M.A.; Lewandowski, A.H.; Zilles, K.; Semendeferi, K.; Allman, J.M.; Craig, A.D.; et al. A volumetric comparison of the insular cortex and its subregions in primates. J. Hum. Evol. 2013, 64, 263–279. [Google Scholar] [CrossRef] [Green Version]
- Nimchinsky, E.A.; Gilissen, E.; Allman, J.M.; Perl, D.P.; Erwin, J.M.; Hof, P.R. A neuronal morphologic type unique to humans and great apes. Proc. Natl. Acad. Sci. USA 1999, 96, 5268–5273. [Google Scholar] [CrossRef] [Green Version]
- Toga, A.W.; Thompson, P.M. Temporal Dynamics of Brain Anatomy. Annu. Rev. Biomed. Eng. 2003, 5, 119–145. [Google Scholar] [CrossRef] [PubMed]
- Amodio, D.M.; Frith, C.D. Meeting of minds: The medial frontal cortex and social cognition. Nat. Rev. Neurosci. 2006, 7, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Loh, K.K.; Petrides, M.; Hopkins, W.D.; Procyk, E.; Amiez, C. Cognitive control of vocalizations in the primate ventrolateral-dorsomedial frontal [VLF-DMF] brain network. Neurosci. Biobehav. Rev. 2017, 82, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Loh, K.K.; Procyk, E.; Neveu, R.; Lamberton, F.; Hopkins, W.D.; Petrides, M.; Amiez, C. Cognitive control of orofacial motor and vocal responses in the ventrolateral and dorsomedial human frontal cortex. Proc. Natl. Acad. Sci. USA 2020, 117, 4994–5005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talairach, J.; Bancaud, J.; Geier, S.; Bordas-Ferrer, M.; Bonis, A.; Szikla, G.; Rusu, M. The cingulate gyrus and human behaviour. Electroencephalogr. Clin. Neurophysiol. 1973, 34, 45–52. [Google Scholar] [CrossRef]
- Ackermann, H.; Ziegler, W. Akinetischer Mutismus-eine Literaturübersicht. Fortschr. Neurol. Psychiatr. 1995, 63, 59–67. [Google Scholar] [CrossRef]
- Benga, O. Intentional communication and the anterior cingulate cortex. Interact. Stud. 2005, 6, 201–221. [Google Scholar] [CrossRef]
- Mundy, P. A review of joint attention and social-cognitive brain systems in typical development and autism spectrum disorder. Eur. J. Neurosci. 2018, 47, 497–514. [Google Scholar] [CrossRef]
- Amiez, C.; Petrides, M. Neuroimaging Evidence of the Anatomo-Functional Organization of the Human Cingulate Motor Areas. Cereb. Cortex 2014, 24, 563–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ide, A.; Dolezal, C.; Fernández, M.; Labbé, E.; Mandujano, R.; Montes, S.; Segura, P.; Verschae, G.; Yarmuch, P.; Aboitiz, F. Hemispheric differences in variability of fissural patterns in parasylvian and cingulate regions of human brains. J. Comp. Neurol. 1999, 410, 235–242. [Google Scholar] [CrossRef]
- Amiez, C.; Wilson, C.R.E.; Procyk, E. Variations of cingulate sulcal organization and link with cognitive performance. Sci. Rep. 2018, 8, 13988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paus, T.; Tomaiuolo, F.; Otaky, N.; Macdonald, D.; Petrides, M.; Atlas, J.; Morris, R.; Evans, A.C. Human Cingulate and Paracingulate Sulci: Pattern, Variability, Asymmetry, and Probabilistic Map. Cereb. Cortex 1996, 6, 207–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jürgens, U.; Pratt, R. The cingular vocalization pathway in the squirrel monkey. Exp. Brain Res. 1979, 34, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Jürgens, U.; Ploog, D. Cerebral representation of vocalization in the squirrel monkey. Exp. Brain Res. 1970, 10, 532–554. [Google Scholar] [CrossRef] [PubMed]
- Aitken, P.G. Cortical control of conditioned and spontaneous vocal behavior in rhesus monkeys. Brain Lang. 1981, 13, 171–184. [Google Scholar] [CrossRef]
- Allman, J.M.; Tetreault, N.A.; Hakeem, A.Y.; Manaye, K.F.; Semendeferi, K.; Erwin, J.M.; Park, S.; Goubert, V.; Hof, P.R. The von Economo neurons in frontoinsular and anterior cingulate cortex in great apes and humans. Anat. Embryol. 2010, 214, 495–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakeem, A.Y.; Sherwood, C.C.; Bonar, C.J.; Butti, C.; Hof, P.R.; Allman, J.M. Von Economo Neurons in the Elephant Brain. Anat. Rec. 2009, 292, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.D.; Procyk, E.; Petrides, M.; Schapiro, S.J.; Mareno, M.C.; Amiez, C. Sulcal Morphology in Cingulate Cortex is Associated with Voluntary Oro-Facial Motor Control and Gestural Communication in Chimpanzees [Pan troglodytes]. Cereb. Cortex 2021, 31, 2845–2854. [Google Scholar] [CrossRef]
- Deen, B.; Koldewyn, K.; Kanwisher, N.; Saxe, R. Functional Organization of Social Perception and Cognition in the Superior Temporal Sulcus. Cereb. Cortex 2015, 25, 4596–4609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belin, P.; Zatorre, R.J.; Lafaille, P.; Ahad, P.A.; Pike, B. Voice-selective areas in human auditory cortex. Nature 2000, 403, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Pitcher, D.; Dilks, D.D.; Saxe, R.R.; Triantafyllou, C.; Kanwisher, N. Differential selectivity for dynamic versus static information in face-selective cortical regions. NeuroImage 2011, 56, 2356–2363. [Google Scholar] [CrossRef]
- Pelphrey, K.A.; Morris, J.P.; Michelich, C.R.; Allison, T.; McCarthy, G. Functional Anatomy of Biological Motion Perception in Posterior Temporal Cortex: An fMRI Study of Eye, Mouth and Hand Movements. Cereb. Cortex 2005, 15, 1866–1876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, K.I.; Moss, H.E.; Stamatakis, E.A.; Tyler, L.K. Binding crossmodal object features in perirhinal cortex. Proc. Natl. Acad. Sci. USA 2006, 103, 8239–8244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciaramidaro, A.; Adenzato, M.; Enrici, I.; Erk, S.; Pia, L.; Bara, B.; Walter, H. The intentional network: How the brain reads varieties of intentions. Neuropsychologia 2007, 45, 3105–3113. [Google Scholar] [CrossRef]
- Wyk, B.C.V.; Hudac, C.M.; Carter, E.J.; Sobel, D.M.; Pelphrey, K.A. Action Understanding in the Superior Temporal Sulcus Region. Psychol. Sci. 2009, 20, 771–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodin, C.; Takerkart, S.; Belin, P.; Coulon, O. Anatomo-functional correspondence in the superior temporal sulcus. Anat. Embryol. 2018, 223, 221–232. [Google Scholar] [CrossRef]
- Moreno, A.; Limousin, F.; Dehaene, S.; Pallier, C. Brain correlates of constituent structure in sign language comprehension. NeuroImage 2018, 167, 151–161. [Google Scholar] [CrossRef]
- Bonte, M.; Frost, M.A.; Rutten, S.; Ley, A.; Formisano, E.; Goebel, R. Development from childhood to adulthood increases morphological and functional inter-individual variability in the right superior temporal cortex. NeuroImage 2013, 83, 739–750. [Google Scholar] [CrossRef]
- Brauer, J.; Neumann, J.; Friederici, A.D. Temporal dynamics of perisylvian activation during language processing in children and adults. NeuroImage 2008, 41, 1484–1492. [Google Scholar] [CrossRef] [Green Version]
- Skeide, M.A.; Brauer, J.; Friederici, A.D. Syntax gradually segregates from semantics in the developing brain. NeuroImage 2014, 100, 106–111. [Google Scholar] [CrossRef]
- Leroy, F.; Cai, Q.; Bogart, S.L.; Dubois, J.; Coulon, O.; Monzalvo, K.; Fischer, C.; Glasel, H.; Van der Haegen, L.; Bénézit, A.; et al. New human-specific brain landmark: The depth asymmetry of superior temporal sulcus. Proc. Natl. Acad. Sci. USA 2015, 112, 1208–1213. [Google Scholar] [CrossRef] [Green Version]
- Leroy, F.; Glasel, H.; Dubois, J.; Hertz-Pannier, L.; Thirion, B.; Mangin, J.-F.; Dehaene-Lambertz, G. Early Maturation of the Linguistic Dorsal Pathway in Human Infants. J. Neurosci. 2011, 31, 1500–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Guen, Y.; Leroy, F.; Auzias, G.; Riviere, D.; Grigis, A.; Mangin, J.-F.; Coulon, O.; Dehaene-Lambertz, G.; Frouin, V. The chaotic morphology of the left superior temporal sulcus is genetically constrained. NeuroImage 2018, 174, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Sliwa, J.; Freiwald, W.A. A dedicated network for social interaction processing in the primate brain. Science 2017, 356, 745–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marciniak, K.; Atabaki, A.; Dicke, P.W.; Thier, P. Disparate substrates for head gaze following and face perception in the monkey superior temporal sulcus. eLife 2014, 3, e03222. [Google Scholar] [CrossRef] [PubMed]
- Fisher, C.; Freiwald, W.A. Contrasting Specializations for Facial Motion within the Macaque Face-Processing System. Curr. Biol. 2015, 25, 261–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roumazeilles, L.; Eichert, N.; Bryant, K.L.; Folloni, D.; Sallet, J.; Vijayakumar, S.; Foxley, S.; Tendler, B.C.; Jbabdi, S.; Reveley, C.; et al. Longitudinal connections and the organization of the temporal cortex in macaques, great apes, and humans. PLoS Biol. 2020, 18, e3000810. [Google Scholar] [CrossRef] [PubMed]
- Sallet, J.; Mars, R.B.; Noonan, M.P.; Andersson, J.L.; O’Reilly, J.X.; Jbabdi, S.; Croxson, P.L.; Jenkinson, M.; Miller, K.L.; Rushworth, M.F.S. Social Network Size Affects Neural Circuits in Macaques. Science 2011, 334, 697–700. [Google Scholar] [CrossRef] [Green Version]
- Ghazanfar, A.A.; Chandrasekaran, C.; Logothetis, N.K. Interactions between the Superior Temporal Sulcus and Auditory Cortex Mediate Dynamic Face/Voice Integration in Rhesus Monkeys. J. Neurosci. 2008, 28, 4457–4469. [Google Scholar] [CrossRef]
- Petkov, C.I.; Kayser, C.; Steudel, T.; Whittingstall, K.; Augath, M.; Logothetis, N.K. A voice region in the monkey brain. Nat. Neurosci. 2008, 11, 367–374. [Google Scholar] [CrossRef]
- Belin, P.; Bodin, C.; Aglieri, V. A “voice patch” system in the primate brain for processing vocal information? Hear. Res. 2018, 366, 65–74. [Google Scholar] [CrossRef]
- Bodin, C.; Trapeau, R.; Nazarian, B.; Sein, J.; Degiovanni, X.; Baurberg, J.; Rapha, E.; Renaud, L.; Giordano, B.L.; Belin, P. Functionally homologous representation of vocalizations in the auditory cortex of humans and macaques. Curr. Biol. 2021, 31, 4839–4844.e4. [Google Scholar] [CrossRef] [PubMed]
- Khandhadia, A.P.; Murphy, A.P.; Romanski, L.M.; Bizley, J.K.; Leopold, D.A. Audiovisual integration in macaque face patch neurons. Curr. Biol. 2021, 31, 1826–1835.e3. [Google Scholar] [CrossRef] [PubMed]
- Meguerditchian, A. “Human-specific” brain lateralization landmarks found in monkeys and their socio-cognitive correlates in both adults and infants [Papio anubis]. In Proceedings of the Communication at NeuroFrance, Strasbourg, France, 19–21 May 2021. [Google Scholar]
- Meguerditchian, A.; Marie, D.; Love, S.A.; Margiotoudi, K.; Bertello, A.; Lacoste, R.; Roth, M.; Nazarian, B.; Anton, J.-L.; Coulon, O. Human-Like Brain Specialization in Baboons: An in vivo Anatomical Mri Study of Language Areas Homologs in 96 Subjects. In The Evolution of Language, Proceedings of the 11th International Conference [EVOLANG11], New Oleans, LA, USA, 20–24 March 2016; Roberts, S.G., Cuskley, C., McCrohon, L., Barceló-Coblijn, L., Feher, O., Verhoef, T., Eds.; EvoLang Scientific Committee: New Orleans, Tulane, 2016. [Google Scholar] [CrossRef]
- Vilberg, K.L.; Rugg, M.D. Memory retrieval and the parietal cortex: A review of evidence from a dual-process perspective. Neuropsychologia 2008, 46, 1787–1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquemot, C.; Scott, S.K. What is the relationship between phonological short-term memory and speech processing? Trends Cogn. Sci. 2006, 10, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Stout, D.; Chaminade, T. Stone tools, language and the brain in human evolution. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Hecht, E.E.; Gutman, D.A.; Khreisheh, N.; Taylor, S.V.; Kilner, J.; Faisal, A.A.; Bradley, B.A.; Chaminade, T.; Stout, D. Acquisition of Paleolithic toolmaking abilities involves structural remodeling to inferior frontoparietal regions. Anat. Embryol. 2015, 220, 2315–2331. [Google Scholar] [CrossRef]
- Flechsig, P. Developmental [myelogenetic] localisation of the cerebral cortex in the human subject. Lancet 1901, 2, 1027–1029. [Google Scholar] [CrossRef] [Green Version]
- Geschwind, N. Disconnexion syndromes in animals and man. Brain 1965, 88, 237. [Google Scholar] [CrossRef] [Green Version]
- Catani, M.; Bambini, V. A model for Social Communication and Language Evolution and Development [SCALED]. Curr. Opin. Neurobiol. 2014, 28, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Lattner, S.; Meyer, M.E.; Friederici, A.D. Voice perception: Sex, pitch, and the right hemisphere. Hum. Brain Mapp. 2005, 24, 11–20. [Google Scholar] [CrossRef]
- Vanduffel, W.; Fize, D.; Peuskens, H.; Denys, K.; Sunaert, S.; Todd, J.T.; Orban, G.A. Extracting 3D from Motion: Differences in Human and Monkey Intraparietal Cortex. Science 2002, 298, 413–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grefkes, C.; Fink, G.R. REVIEW: The functional organization of the intraparietal sulcus in humans and monkeys. J. Anat. 2005, 207, 3–17. [Google Scholar] [CrossRef]
- Orban, G.A.; Claeys, K.; Nelissen, K.; Smans, R.; Sunaert, S.; Todd, J.T.; Wardak, C.; Durand, J.-B.; Vanduffel, W. Mapping the parietal cortex of human and non-human primates. Neuropsychologia 2006, 44, 2647–2667. [Google Scholar] [CrossRef]
- Budisavljevic, S.; Castiello, U.; Begliomini, C. Handedness and White Matter Networks. Neuroscientist 2021, 27, 88–103. [Google Scholar] [CrossRef] [PubMed]
- Arbib, M.A. From monkey-like action recognition to human language: An evolutionary framework for neurolinguistics. Behav. Brain Sci. 2005, 28, 105–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzolatti, G.; Sinigaglia, C. The functional role of the parieto-frontal mirror circuit: Interpretations and misinterpretations. Nat. Rev. Neurosci. 2010, 11, 264–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eickhoff, S.B.; Schleicher, A.; Zilles, K.; Amunts, K. The Human Parietal Operculum. I. Cytoarchitectonic Mapping of Subdivisions. Cereb. Cortex 2006, 16, 254–267. [Google Scholar] [CrossRef]
- Habib, M.; Robichon, F.; Levrier, O.; Khalil, R.; Salamon, G. Diverging Asymmetries of Temporo-parietal Cortical Areas: A Reappraisal of Geschwind/Galaburda Theory. Brain Lang. 1995, 48, 238–258. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Frey, S.H.; Newman-Norlund, R.; Grafton, S.T. A Distributed Left Hemisphere Network Active during Planning of Everyday Tool Use Skills. Cereb. Cortex 2005, 15, 681–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, J.W. Cortical Networks Related to Human Use of Tools. Neurosci 2006, 12, 211–231. [Google Scholar] [CrossRef]
- Cheng, L.; Zhang, Y.; Li, G.; Wang, J.; Hopkins, W.D.; Sherwood, C.C.; Gong, G.; Fan, L.; Jiang, T. Divergent Connectional Asymmetries of the Inferior Parietal Lobule Shape Hemispheric Specialization in Humans, Chimpanzees, and Macaque Monkeys. Neuroscience 2021. [Google Scholar] [CrossRef]
- Anderson, J.R.; Gallup, G.G. Mirror self-recognition: A review and critique of attempts to promote and engineer self-recognition in primates. Primates 2015, 56, 317–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecht, E.; Mahovetz, L.M.; Preuss, T.M.; Hopkins, W.D. A neuroanatomical predictor of mirror self-recognition in chimpanzees. Soc. Cogn. Affect. Neurosci. 2017, 12, 37–48. [Google Scholar] [CrossRef] [Green Version]
- Stout, D.; Hecht, E.; Khreisheh, N.; Bradley, B.; Chaminade, T. Cognitive Demands of Lower Paleolithic Toolmaking. PLoS ONE 2015, 10, e0121804. [Google Scholar] [CrossRef] [Green Version]
- Gannon, P.J.; Kheck, N.M.; Braun, A.R.; Holloway, R.L. Planum parietale of chimpanzees and orangutans: A comparative resonance of human-like planum temporale asymmetry. Anat. Rec. Part A 2005, 287, 1128–1141. [Google Scholar] [CrossRef] [PubMed]
- Taglialatela, J.P.; Dadda, M.; Hopkins, W.D. Sex differences in asymmetry of the planum parietale in chimpanzees [Pan troglodytes]. Behav. Brain Res. 2007, 184, 185–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penfield, W.; Boldrey, E. somatic motor and sensory representation in the cerebral cortex of man as studied by electrical stimulation. Brain 1937, 60, 389–443. [Google Scholar] [CrossRef]
- Yousry, T. Localization of the motor hand area to a knob on the precentral gyrus. A New Landmark. Brain 1997, 120, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Amunts, K.; Schlaugb, G.; Schleichera, A.; Steinmetzb, H.; Dabringhausa, A.; Roland, P.E.; Zilles, K. Asymmetry in the Human Motor Cortex and Handedness. NeuroImage 1996, 4, 216–222. [Google Scholar] [CrossRef]
- Cykowski, M.D.; Coulon, O.; Kochunov, P.V.; Amunts, K.; Lancaster, J.L.; Laird, A.R.; Glahn, D.C.; Fox, P.T. The Central Sulcus: An Observer-Independent Characterization of Sulcal Landmarks and Depth Asymmetry. Cereb. Cortex 2008, 18, 1999–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.Y.; Klöppel, S.; Rivière, D.; Perrot, M.; Frackowiak, R.; Siebner, H.; Mangin, J.-F. The effect of handedness on the shape of the central sulcus. NeuroImage 2012, 60, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Häberling, I.S.; Corballis, P.M.; Corballis, M.C. Language, gesture, and handedness: Evidence for independent lateralized networks. Cortex 2016, 82, 72–85. [Google Scholar] [CrossRef]
- Crow, T. Directional asymmetry is the key to the origin of modern Homo sapiens [the Broca-Annett axiom]: A reply to Rogers’ review of The Speciation of Modern Homo Sapiens. Laterality Asymmetries Body Brain Cogn. 2004, 9, 233–242. [Google Scholar] [CrossRef]
- Warren, J.M. Handedness and laterality in humans and other animals. Physiol. Psychol. 1980, 8, 351–359. [Google Scholar] [CrossRef] [Green Version]
- Hobaiter, C.; Byrne, R.W. Laterality in the gestural communication of wild chimpanzees. Ann. N. Y. Acad. Sci. 2013, 1288, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, W.D.; Cantalupo, C. Handedness in Chimpanzees [Pan troglodytes] Is Associated with Asymmetries of the Primary Motor Cortex but Not With Homologous Language Areas. Behav. Neurosci. 2004, 118, 1176–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dadda, M.; Cantalupo, C.; Hopkins, W.D. Further evidence of an association between handedness and neuroanatomical asymmetries in the primary motor cortex of chimpanzees [Pan troglodytes]. Neuropsychologia 2006, 44, 2582–2586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margiotoudi, K.; Marie, D.; Claidière, N.; Coulon, O.; Roth, M.; Nazarian, B.; Lacoste, R.; Hopkins, W.D.; Molesti, S.; Fresnais, P.; et al. Handedness in monkeys reflects hemispheric specialization within the central sulcus. An in vivo MRI study in right- and left-handed olive baboons. Cortex 2019, 118, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Phillips, K.A.; Sherwood, C.C. Primary motor cortex asymmetry is correlated with handedness in capuchin monkeys [cebus apella]. Behav. Neurosci. 2005, 119, 1701–1704. [Google Scholar] [CrossRef]
- Nudo, R.; Jenkins, W.; Merzenich, M.; Prejean, T.; Grenda, R. Neurophysiological correlates of hand preference in primary motor cortex of adult squirrel monkeys. J. Neurosci. 1992, 12, 2918–2947. [Google Scholar] [CrossRef] [Green Version]
- Bouziane, S.; Loh, K.K.; Becker, Y.; Brunschvig, S.; Picchiottino, A.; Sein, J.; Coulon, O.; Velly, L.; Renaud, L.; Meguerditchian, A. Early structural asymmetry in the central sulcus is associated with handedness in infant baboons. In Proceedings of the OHBM 2021: 27th Annual Meeting of the Organization for Human Brain Mapping, Monday, Virtual, 21–25 June 2021. [Google Scholar]
Figure 1.
Illustration of the reviewed key structures in a monkey brain. In brown: Broca’s area, in yellow: the Insula, in red: the Arcuate Fasciculus, in blue: the Central Sulcus, in green: the Planum Temporale, in light red: the Planum Parietale, in grey: the Superior Temporal Sulcus, in purple: the Cingulate Sulcus. Arcuate: Arcuate Fasciculus, CS: Central Sulcus, PT: Planum Temporale, PP: Planum Parietale, STS: Superior Temporal Sulcus.
Figure 1.
Illustration of the reviewed key structures in a monkey brain. In brown: Broca’s area, in yellow: the Insula, in red: the Arcuate Fasciculus, in blue: the Central Sulcus, in green: the Planum Temporale, in light red: the Planum Parietale, in grey: the Superior Temporal Sulcus, in purple: the Cingulate Sulcus. Arcuate: Arcuate Fasciculus, CS: Central Sulcus, PT: Planum Temporale, PP: Planum Parietale, STS: Superior Temporal Sulcus.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Becker, Y.; Meguerditchian, A. Structural Brain Asymmetries for Language: A Comparative Approach across Primates. Symmetry 2022, 14, 876. https://doi.org/10.3390/sym14050876
AMA Style
Becker Y, Meguerditchian A. Structural Brain Asymmetries for Language: A Comparative Approach across Primates. Symmetry. 2022; 14(5):876. https://doi.org/10.3390/sym14050876
Chicago/Turabian StyleBecker, Yannick, and Adrien Meguerditchian. 2022. "Structural Brain Asymmetries for Language: A Comparative Approach across Primates" Symmetry 14, no. 5: 876. https://doi.org/10.3390/sym14050876
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.