Dynamics of Laterality in Lake Tanganyika Scale-Eaters Driven by Cross-Predation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling of Scale-Eaters to Assess the Ratios of Laterality
2.2. Sampling of Pairs of Hunting Scale-Eaters and Their Victim Fish to Assess the Combination of Laterality at Each Predation Event
3. Results
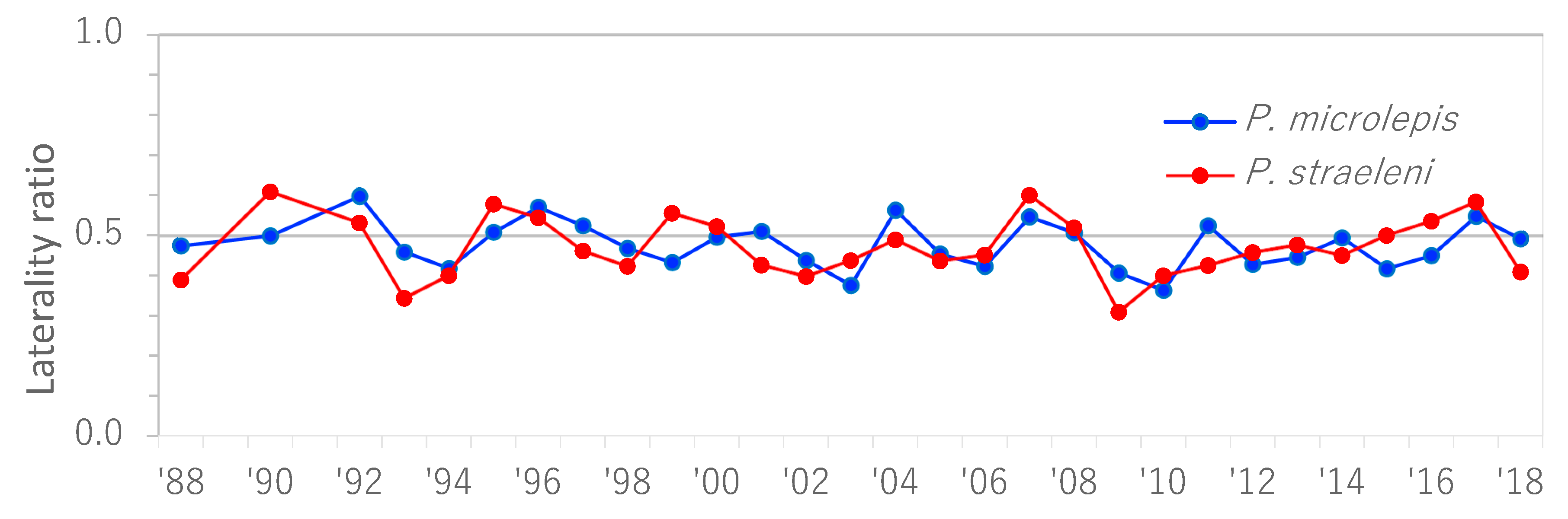
3.1. Temporal Changes in the Laterality of Two Scale-Eaters
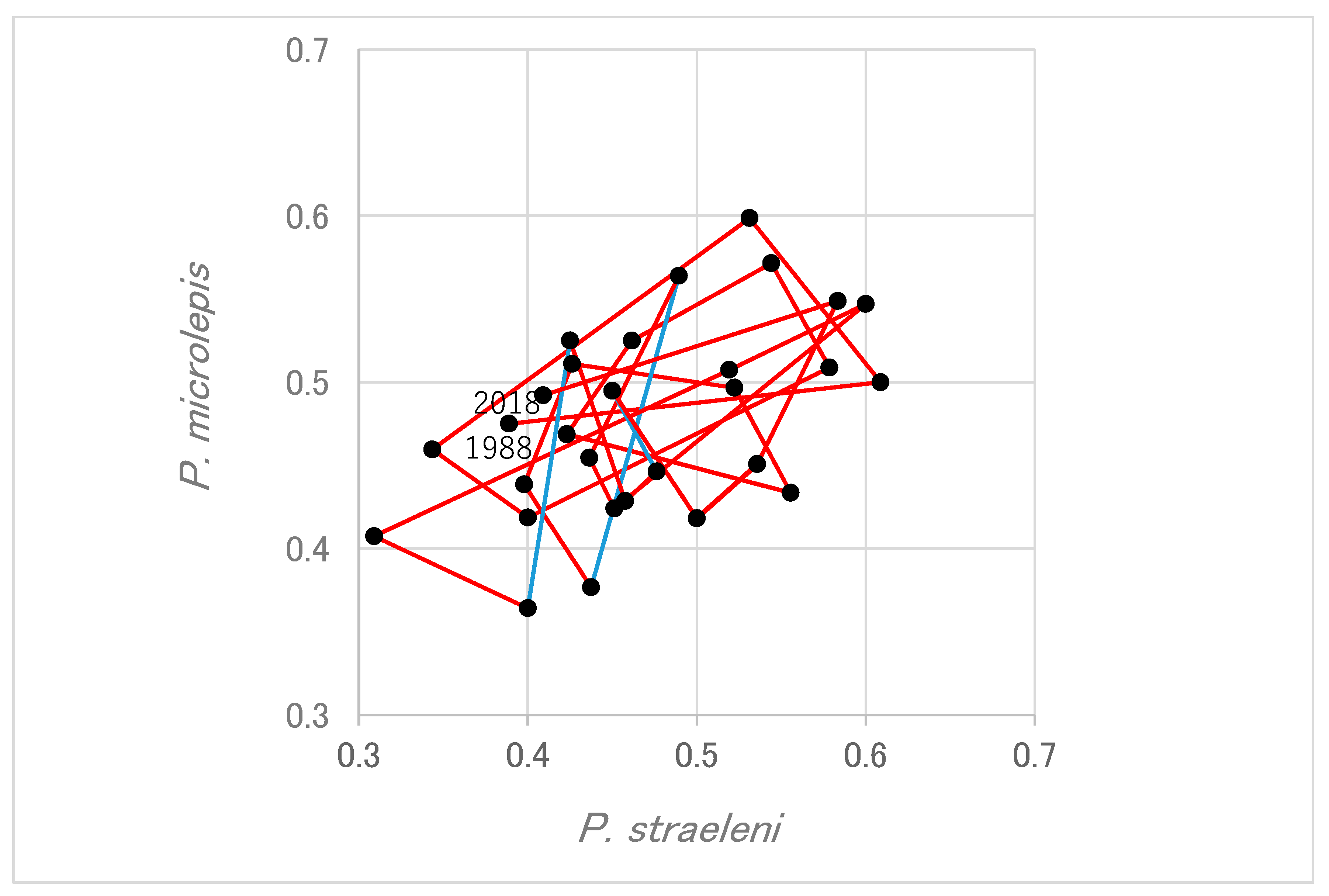
3.2. Correspondence of Fish Laterality between Individual Scale-Eater and Its Prey
4. Discussion
4.1. Relation between the Two Species of Scale-Eater
4.2. Predator and Prey Relationship
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bisazza, A.; Rogers, L.J.; Vallortigara, G. The origins of cerebral asymmetry: A review of evidence of behavioral and brain lateralization in fishes, reptiles and amphibians. Neurosci. Biobehav. Rev. 1998, 22, 411–426. [Google Scholar] [CrossRef]
- Vallortigara, G.; Rogers, L.J.; Bisazza, A. Possible evolutionary origins of cognitive brain lateralization. Brain Res. Rev. 1999, 30, 164–175. [Google Scholar] [CrossRef]
- Rogers, L.J.; Vallortigara, G.; Andrew, R.J. Divided Brains: The Biology and Behaviour of Brain Asymmetries; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Vallortigara, G.; Versace, E. Laterality at the neural, cognitive, and behavioral levels. In APA Handbooks in Psychology. APA Handbook of Comparative Psychology: Basic Concepts, Methods, Neural Substrate, and Behavior; Call, J., Burghardt, G.M., Pepperberg, I.M., Snowdon, C.T., Zentall, T., Eds.; American Psychological Association: Washington, DC, USA, 2017; pp. 557–577. [Google Scholar]
- MacNeilage, P.; Rogers, L.J.; Vallortigara, G. Evolutionary origins of your left and right brain. Sci. Am. 2009, 301, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Vallortigara, G.; Rogers, L.J. Survival with an asymmetrical brain: Advantages and disadvantages of cerebral lateralization. Behav. Brain Sci. 2005, 28, 575–633. [Google Scholar] [CrossRef]
- Vallortigara, G.; Chiandetti, C.; Sovrano, V.A. Brain asymmetry (animal). WIREs Cogn. Sci. 2011, 2, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.J.; Vallortigara, G. When and why did brains break symmetry? Symmetry 2015, 7, 2181–2194. [Google Scholar] [CrossRef]
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry as a measure of development stability: Implications of non-normal distributions and power of statistical tests. Acta Zool. Fennica 1992, 191, 57–72. [Google Scholar]
- Rogers, L.J.; Andrew, R.J. Comparative Vertebrate Lateralization; Cambridge University Press: London, UK, 2002; ISBN 0-521-78161-2. [Google Scholar]
- Ströckens, F.; Gunturkün, O.; Ocklenburg, S. Limb preferences in non-human vertebrates. Laterality Asymmetries Body Brain Cogn. 2013, 18, 536–575. [Google Scholar] [CrossRef]
- Ghirlanda, S.; Vallortigara, G. The evolution of brain lateralization: A game-theoretical analysis of population structure. Proc. R. Soc. Lond. B 2004, 271, 853–857. [Google Scholar] [CrossRef] [PubMed]
- Ghirlanda, S.; Frasnelli, E.; Vallortigara, G. Intraspecific competition and coordination in the evolution of lateralization. Philos. Tranzactions R. Soc. B 2009, 364, 861–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallortigara, G. The Evolutionary psychology of left and right: Costs and benefits of lateralization. Dev. Phychobiol. 2006, 48, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Hori, M. Frequency-dependent natural selection in the handedness of scale-eating cichlid fish. Science 1993, 260, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Liem, K.F.; Stewart, D.J. Evolution of the scale-eating cichlid fishes of Lake Tanganyika: A generic revision with a description of a new species. Bull. Mus. Comp. Zool. 1976, 147, 319–350. [Google Scholar]
- Hori, M. Feeding relationships among cichlid fishes in Lake Tanganyika: Effects of intra- and interspecific variation of feeding behavior on their coexistence. Ecol. Int. Bull. 1991, 19, 89–101. [Google Scholar]
- Takahashi, R.; Moriwaki, T.; Hori, M. Foraging behaviour and functional morphology of two scale-eating cichlids from Lake Tanganyika. J. Fish Biol. 2007, 70, 1458–1469. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Hori, M.; Oda, Y. Lateralized kinematics of predation behavior in a Lake Tanganyika scale-eating cichlid fish. PLoS ONE 2012, 7, e29272. [Google Scholar] [CrossRef] [PubMed]
- Hata, H.; Yasugi, M.; Hori, M. Jaw laterality and related handedness in the hunting behavior of a scale-eating characin, Exodon paradoxus. PLoS ONE 2011, 6, e29349. [Google Scholar] [CrossRef]
- Hori, M.; Ochi, H.; Kohda, M. Inheritance pattern of lateral dimorphism in two cichlids (a scale eater, Perissodus microlepis, and an herbivore, Neolamprologus moorii) in Lake Tanganyika. Zool. Sci. 2007, 24, 489–492. [Google Scholar] [CrossRef] [PubMed]
- Stewart, T.A.; Albertson, R.C. Evolution of a unique predatory feeding apparatus: Functional anatomy, development and a genetic locus for jaw laterality in Lake Tanganyika scale-eating cichlids. BMC Biol. 2010, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Hata, H.; Takahashi, R.; Ashiwa, H.; Awata, S.; Takeyama, T.; Kohda, M.; Hori, M. Inheritance patterns of lateral dimorphism examined by breeding experiments with the Tanganyikan cichlid (Julidochromis transcriptus) and the Japanese medaka (Oryzias latipes). Zool. Sci. 2012, 29, 49–53. [Google Scholar] [CrossRef]
- Hata, H.; Hori, M. Inheritance patterns of morphological laterality in mouth opening of zebrafish, Danio rerio. Laterality 2012, 17, 741–754. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Hori, M. Unstable evolutionarily stable strategy and oscillation: A model on lateral asymmetry in scale-eating cichlids. Am. Nat. 1994, 144, 1001–1020. [Google Scholar] [CrossRef]
- Takahashi, S.; Hori, M. Oscillation maintains polymorphisms—A model of lateral asymmetry in two competing scale-eating cichlids. J. Theor. Biol. 1998, 195, 1–12. [Google Scholar] [CrossRef]
- Hori, M. Structure of littoral fish communities organized by their feeding activities. In Fish Communities in Lake Tanganyika; Kawanabe, H., Hori, M., Nagoshi, M., Eds.; Kyoto University Press: Kyoto, Japan, 1997; pp. 275–298. ISBN 4-87698-042-X. [Google Scholar]
- Takeuchi, Y.; Ochi, H.; Kohda, M.; Sinyinza, D.; Hori, M. A 20-year census of a rocky littoral fish community in Lake Tanganyika. Ecol. Freshw. Fish 2010, 19, 239–248. [Google Scholar] [CrossRef]
- Takahashi, R.; Watanabe, K.; Nishida, M.; Hori, M. Evolution of feeding secialization in Tanganyikan scale-eating cichlids: A molecular phylogenetic approach. BMC Evol. 2007, 7, 195. [Google Scholar]
- Hori, M. Mutualism and commensalism in the fish community of Lake Tanganyika. In Evolution and Coadaptation in Biotic Communities; Kawano, S., Connell, J.H., Hidaka, T., Eds.; Tokyo University Press: Tokyo, Japan, 1987; pp. 219–239. [Google Scholar]
- Hori, M.; Nakajima, M.; Hata, H.; Yasugi, M.; Takahashi, S.; Nakae, M.; Yamaoka, K.; Kohda, M.; Kitamura, J.; Maehata, M.; et al. Laterality is universal among fishes but increasingly cryptic among derived groups. Zool. Sci. 2017, 34, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, M.; Yodo, T.; Katano, O. Righty fish are hooked on the right side of their mouths—Observations from an angling experiment with largemouth bass, Microperus salmoides. Zool. Sci. 2007, 24, 855–859. [Google Scholar] [CrossRef]
- Seki, S.; Kohda, M.; Hori, M. Asymmetry of mouth morph of a freshwater goby, Rhinogobius flumineus. Zool. Sci. 2000, 17, 1321–1325. [Google Scholar] [CrossRef]
- Matsui, S.; Takeuchi, Y.; Hori, M. Relation between morphological antisymmetry and behavioral laterality in a Poeciliid fish. Zool. Sci. 2013, 30, 613–618. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Hori, M. Behavioural laterality in the shrimp-eating cichlid fish, Neolamprologus fasciatus, in Lake Tanganyika. Anim. Behav. 2008, 75, 1359–1366. [Google Scholar] [CrossRef]
- Yasugi, M.; Hori, M. Predominance of cross-predation between lateral morphs in a largemouth bass and a freshwater goby. Zool. Sci. 2011, 28, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Yasugi, M.; Hori, M. Lateralized behavior in the attacks of largemouth bass on Rhinogobius gobies corresponding to their morphological antisymmetry. J. Exp. Biol. 2012, 215, 2390–2398. [Google Scholar] [CrossRef] [PubMed]
- Yasugi, M.; Hori, M. Predominance of parallel- and cross-predation in anglerfish. Mar. Ecol. 2016, 37, 576–587. [Google Scholar] [CrossRef]
- Hori, M.; Yamaoka, K.; Takamura, K. Abundance and micro-distribution of cichlid fishes on a rocky shore of Lake Tanganyika. Afr. Study Monogr. 1983, 3, 25–38. [Google Scholar]
- Hata, H.; Yasugi, M.; Takeuchi, Y.; Hori, M. Distinct lateral dimorphism in the jaw morphology of the scale-eating cichlids, Perissodus microlepis and P. straeleni. Ecol. Evol. 2013, 3, 4641–4647. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, Y.; Hori, M.; Myint, O.; Kohda, M. Lateral bias of agonistic responses to mirror images and morphological asymmetry in the Siamese fighting fish (Betta splendens). Behav. Brain Res. 2010, 208, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Hori, M. Coexistence of competing species by the oscillation polymorphism. J. Theor. Biol. 2005, 235, 591–596. [Google Scholar] [CrossRef]
- Nakajima, M.; Matsuda, H.; Hori, M. A population genetic model for lateral dimorphism frequency in fishes. Am. Nat. 2005, 163, 692–698. [Google Scholar] [CrossRef]
- Nakajima, M.; Matsuda, H.; Hori, M. Persistence and fluctuation of lateral dimorphism in fishes. Am. Nat. 2004, 163, 692–698. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Scale-Eater | Victim Fish | Species | Scale-Eater | Victim Fish | ||
|---|---|---|---|---|---|---|---|
| R | L | R | L | ||||
| P. microlepis | R | 3 | 19 | P. straeleni | R | 4 | 21 |
| L | 25 | 6 | L | 22 | 4 | ||
| Total: 53 | Total: 51 | ||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hori, M.; Kohda, M.; Awata, S.; Takahashi, S. Dynamics of Laterality in Lake Tanganyika Scale-Eaters Driven by Cross-Predation. Symmetry 2019, 11, 119. https://doi.org/10.3390/sym11010119
Hori M, Kohda M, Awata S, Takahashi S. Dynamics of Laterality in Lake Tanganyika Scale-Eaters Driven by Cross-Predation. Symmetry. 2019; 11(1):119. https://doi.org/10.3390/sym11010119
Chicago/Turabian StyleHori, Michio, Masanori Kohda, Satoshi Awata, and Satoshi Takahashi. 2019. "Dynamics of Laterality in Lake Tanganyika Scale-Eaters Driven by Cross-Predation" Symmetry 11, no. 1: 119. https://doi.org/10.3390/sym11010119