
Philosophical Approach to Neural Autoantibodies in Psychiatric Disease—Multi-Systemic Dynamic Continuum from Protective to Harmful Autoimmunity in Neuronal Systems

1
Department of Psychiatry and Psychotherapy, University Medical Center Göttingen, 37075 Göttingen, Germany
2
Translational Psychoneuroscience, University Medical Center Göttingen, 37075 Göttingen, Germany
Antibodies 2023, 12(1), 1; https://doi.org/10.3390/antib12010001
Submission received: 4 November 2022
/
Revised: 7 December 2022
/
Accepted: 20 December 2022
/
Published: 23 December 2022
Abstract
:(1) Background: philosophical views are important to enable a general and multi-systemic view of the potential understanding of autoimmunity in psychiatric disease that is not solely reflected by an immunological viewpoint. (2) Methods: we reviewed current theories of autoimmunity. (3) Results: we propose a novel area view integrating the “self/non-self” and “continuity” model into the expression of varied forms of autoimmunity in psychiatric disease, ranging from protective to harmful autoimmunity consequences framed into micro-systems (nerve cells) and macro-systems (neuronal networks), termed the “multi-systemic dynamic continuum model”. (4) Conclusions: autoimmunity’s dynamic spectrum is delineated here as something that probably functions as a whole entity to maintain, first of all, human homeostasis in behavior affecting cells or neuronal networks differently, and secondly to prevent psychiatric disease.
1. Introduction—Immunological Conception of Central Nervous System Autoimmunity
The term central nervous system (CNS) autoimmunity refers to an immune reaction directed against the body´s own antigens within CNS structures from an immunological point of view. The immune reaction is so diverse and complex that it cannot be simplified as two main mechanisms. Autoantibody-associated psychiatric disease can, however, be categorized as two main immune mechanisms, although other important immune mechanisms also exist. One potential immune mechanism in autoimmune-mediated secondary psychiatric disorders that are associated with autoantibodies to intracellular antigens, is likely to be characterized predominantly by a T cell-dependent mechanism. This suggestion is based on the observation that in encephalitides associated with antibodies and intracellular antigens, CD8+ T cells were frequently found in encephalitic brain tissue [1]. The other major mechanism is characterized by the action of neural autoantibodies, such as antibodies against the N-methyl-D-aspartate receptor [2] detected in an autoimmune encephalitis of psychiatric phenotype [3]. Neural autoantibodies are part of the immune defense, which can play a protective role in destroying cancer (as paraneoplastic antibodies on the one hand), but they can also be destructive when accompanying CNS inflammation and neuronal nerve cell damage [4]. Neuronal autoantibodies may be associated with psychiatric disorders, although no evidence of brain inflammation or brain damage has been found. In such conditions, the significance of elevated levels of neural autoantibodies remains unclear according to what we know now. Specific biomarkers and clinical conditions with specific criteria, as formulated for autoimmune encephalitis [5], autoimmune psychosis [6], or autoimmune-based psychiatric syndrome [7], indicate the possible, or even probable, autoimmune origin of psychiatric symptoms in association with the detection of neural autoantibodies.
This review addresses, on the one hand, whether autoimmunity can be reduced to the distinction between an attack on the self as an autoimmune response and an attack on the non-self as an action that is not part of an autoimmune response. On the other hand, it may also be that autoimmunity, in particular, which has been found in psychiatric disorders probably mediated by autoantibodies, is more complex than the theory of the distinction between self and non-self. We favor the latter, and will make some arguments in our review. In our view, in addition to autoimmunity in psychiatric disorders (which may be evidenced by autoantibodies on the one hand, but may also be due to T cell-driven immune responses on the other), many other factors that may contribute to autoimmunity are important, such as T cells, regulatory T cells (Tregs), and B cells.
2. Methods
We searched PubMed for articles on theories of autoimmunity and autoantibodies related to psychiatric disorders. Our narrative review is based on the selection of these articles.
3. Results
We first introduce relevant theories of autoimmunity and then describe autoantibodies in psychiatric disorders as a possible manifestation of autoimmunity. Later, three different types of autoimmunity are presented, representing possible autoimmunity in autoantibody-associated psychiatric disorders.
3.1. Philosophical Conception of CNS Autoimmunity
A philosophical consideration of CNS autoimmunity comprises two main models entailing explanations, namely the “self-model or non-self-model” as suggested by Burnet [8], and the “continuity hypothesis”, proposed by Pradeu and Carosella [9]. The “self/non-self-model” is currently one of the most common concepts for explaining CNS autoimmunity. This model is based on the assumption that foreign (non-self) elements trigger an immune reaction, whereas the self does not cause an immune reaction [10]. However, there are serious limitations associated with this model, as the immune system needs to include reactions against the self, i.e., if cancer cells develop. Furthermore, other immune cells such as (the much later discovered) regulatory T cells controlling other lymphocyte populations are not considered in this model [9]. Regulatory T cells as “self-cells” can be regarded as cells that respond to other self-cells, such as T-lymphocytes. Thus, immune cells can also be directed against “self” structures, resulting in an autoimmune regulation of immune cell populations. A model differing from the “self/non-self model”, has been developed, namely the “continuity model” [9]. The “continuity model” [9] states that the immune system serves to continuously distinguish antigenic epitopes present in the organism independently of their “self/non-self” nature. The continuity of immune interactions can be interrupted by novel epitopes. The “continuity” hypothesis represents a philosophical shift from a substance-related biological identity to a biological identity based on continuity [9]. The continuity model is suitable for addressing the biological identity of potential autoimmunity, but has limitations concerning the expression forms of CNS autoimmunity in psychiatric disease, addressed by this review below. Another development is the “discontinuity theory,” which states that immune responses depend on the discontinuity of the antigen [11,12]. This theory establishes general principles that are essential to creating a unified immune concept. Selected current concepts of CNS autoimmunity are delineated in Table 1.
3.2. Autoantibody-Mediated Psychiatric Disease
To understand the role of autoimmunity in psychiatric disease, we will elucidate the role neural autoantibodies play in psychiatric disease. Neural autoantibodies are increasingly detected in association with different psychiatric disorders, ranging from dementia to psychotic disorders [6,19,20,21,22]. The meaning of these neural autoantibodies is still enigmatic, although several models have postulated functions in aberrant synaptic transmission [23] and networking [24], as well as inducing specific psychiatric symptoms such as psychosis [25]. An autoimmune basis of psychiatric syndromes is fulfilled if specific criteria are present, such as (1) CNS inflammation measured by CSF pleocytosis, intrathecal IgG synthesis or other signs of brain inflammation in neuroimaging, in conjunction with (2) detecting autoantibodies in the serum and/or blood, (3) specific neuropsychiatric constellations termed as “autoimmune indicators”, such as seizures, motor abnormalities such as catatonia, fever, altered consciousness, severe cognitive dysfunction, or an adverse response to psychopharmacologic drugs [7]. By applying these criteria, clinicians can diagnose an autoimmune origin as being possible or definitive [7]. Autoantibody-mediated psychiatric disease can, on the other hand, originate from an organic autoimmune encephalitis when mainly relying on the Graus criteria [5]. Their criteria were slightly modified in a recent case series in which patients presenting primarily psychiatric symptoms were classified as psychiatric autoimmune encephalitis patients [21]. What is common to both classifications is the required occurrence of neural autoantibodies [5,7]. However, in the Hansen et al. [7] classification, the formulated criteria require signs of brain damage or inflammation via specific additional diagnostics to diagnose an autoimmune-originating psychiatric syndrome. In contrast, the presence in a recent case series of well-characterized autoantibodies such as LGI1, NMDAR or MOG antibodies [21] was regarded as a sufficient criterion to assume a probable psychiatric autoimmune encephalitis. Thus, there is substantial controversy at the moment about which diagnostic evidence must be present to classify a psychiatric disease as being autoimmune-based. We favor a more conservative approach to classifying a psychiatric syndrome as autoimmune-mediated. Several autoantibodies, such as those that are paraneoplastic, are not believed to contribute to the pathogenesis of the disease on their own, but rather in conjunction with T cells. The concept of autoimmunity in psychiatric disease probably encompasses two different mechanisms: (1) an autoantibody-mediated autoimmunity mechanism, mainly detected in patients with antibodies against membrane-surface autoantibodies, and (2), a T cell-mediated mechanism of autoimmunity detected in patients with antibodies against intracellular target antigens. Both mechanisms share an immunity process requiring the ability to have previously made a distinction between the group property “self” and its counterpart “non-self” or “foreign”. Disease states such as autoantibody-associated psychiatric disease incorporate the recognition of the “foreign” either as one’s own tissue or as microorganisms. This process is active and dynamic, so that the borders between the self and of identifying the “foreign” are not constant. The immune system is activated when “foreign” elements are recognized within the body. This recognition of the “foreign” involves a prior process of self-tolerance. The breakdown of self-tolerance induces an immune-system activation, enabling the self and non-self dichotomy, but it also leads to the continuous recognition of antigen epitopes. Such an immune-system activation can be “protective” or “aggressive”. The term protective autoimmunity refers to the autoantibodies and B cells responsible for autoantibody production in psychiatric autoimmune encephalitis or autoimmune-mediated psychiatric syndromes, but also to T cell pathology, as seen in patients with probable autoimmune-mediated psychiatric syndromes associated with intracellular autoantibodies, as described below. In our model presented in Section 3.3, Section 3.4 and Section 3.5, we establish a biological basis for psychiatric disorders that links neuronal systems to autoimmune processes. Autoinflammation in autoimmune states in organic psychiatric disorders implies minimal changes, such as mild encephalitis, and in brain tissue changes such as inflammation or brain injury.
3.3. Protective Autoimmunity
We found a protective autoimmunity with autoantibodies if the immune reaction takes place and eliminates “foreign” or “continuous” antigen epitope elements, such as a paraneoplastic action that causes the elimination of a tumor. Note that neural autoantibodies can be paraneoplastic in nature, such as intracellular antibodies, which means that neural autoantibodies can be the precursor or even consequence of cancer. Neural autoantibodies are often indicative of an unspecified immune response against tumor antigens sharing features with the targeted neural antigens. Thus, neural autoantibodies may be protective, as they are indications of a tumor immunity process taking place. Moreover, these autoantibodies may indicate a potentially harmful future, in that the appearance of these autoantibodies could act as the body’s alarm system before a potential cancer develops, and can thus be interpreted as protective.
No symptoms are felt from the patient and the action leads to optimization of survival. The immune reactions are believed to be part of an active process that is regarded as a complex “normal” physiology [26]. It is known that natural autoantibodies exist that have protective roles and exist in normal [27] and also in disease conditions [28]. Protective autoimmunity is present to recognize epitopes that suggest potential damage for the body. Such a “healthy” autoimmunity protects the body from damage by cancer (Figure 1A). Protective autoimmunity is mainly based on micro-immune reactions on the cell level on the one hand, while on the other hand, protective immune reactions against antigenic structures in the nervous system (whose function is protective) may also affect neuronal networks on the macro level (Figure 1B). Thus, protective autoimmunity could help neuronal systems and neuronal tissue repair and restore themselves to maintain their previous physiological functions.
3.4. Regulatory Autoimmunity
Psychiatric symptoms coinciding with autoimmune-based psychiatric disease are potential correlates of CNS autoinflammation. Although such patients suffer from transient and reversible psychiatric symptoms, no relevant brain damage or brain inflammation is detected when conducting diagnostics such as EEG, CSF analysis, and MRI. The immunity process serves to regulate immunity against the self along with ongoing negotiation of the interaction between the self and “non-self”, as described by Tauber [29], or via continuous recognition of endogenous or exogenous epitopes, as proposed by Pradeu and Carosella [9]. Autoimmunity is part of a balanced physiologic process. It seems to be an action that regulates dynamic self-homeostasis. The regulatory T cells (Tregs) are a paradigmatic example of a regulatory autoimmune reaction to drive or limit cellular immune responses (Figure 1A). Tregs, such as Foxp3-expressing CD4+ Treg cells, are central to preventing the loss of self-tolerance so that autoimmunity states can be induced. In addition, these cells contribute to maintaining immune homeostasis [30]. Treg cells are not directly associated with autoantibodies. Treg cell functions that are not dependent on autoantibodies include the suppression of other T cells that dampen the immune or autoimmune response. Tregs may also limit CNS inflammation by secreting interleukin 10. Treg cells may have neuroprotective properties by placing microglia in a neuroprotective state [31]. In addition, there is a specific subset of Treg cells characterized by CD69+ that is increased in mice and humans and thought to control neuroinflammation [31]. When brain-resistant Treg cells are lost, neuroinflammatory states are enhanced. These specific Tregs are thus important in regulating autoimmunity in the CNS. Another function potentially relevant to Treg cells’ regulatory capability is their ability to downregulate calcium influx, a function impaired in patients with autoimmune CNS diseases [32]. On the other hand, Treg cells may also promote autoantibody production by producing neuritin via follicular Treg cells, which could thereby inhibit autoantibody production by B cells (B cell-driven autoimmunity) [33]. Thus, B cells producing autoantibodies are also an important aspect of regulatory autoimmunity. Moreover, regulatory autoimmunity plays a role in the clearance of cancer cells by cytotoxic CD8+ T cells without the aid of neural autoantibodies. The central role of cytotoxic CD8+ T cells in cancer cell clearance was recently confirmed in an animal study [34]. Taken together, regulatory autoimmunity could also help to reduce brain damage or inflammation in brain tissues already affected by autoimmunity processes.
3.5. Harmful Autoimmunity
Autoimmune processes in the CNS can cause severe damage and neuronal loss. Persistent psychiatric symptoms associated with neural autoantibodies might be a severe and harmful consequence of CNS autoimmunity (Figure 1A). Harmful autoimmunity should also be considered from a psychoneuroimmunological perspective. Anomalies in one cognitive system can affect other cognitive systems. Autoantibodies can enter the brain and induce a severe inflammation that can also trigger transient or persistent focal brain damage, such as hippocampal damage in LGI1 autoantibody-associated disease [4] or unspecific axonal brain damage indicated by elevated neurofilament light chains in the cerebrospinal fluid in LGI1 and NMDAR encephalitis [35]. Harmful autoimmunity, compared to regulatory or protective autoimmunity, is more relevant for diagnostics, as in conditions of harmful autoimmunity brain damage or brain inflammation can be found. The harmful type of autoimmunity ultimately affects both neural networks and nerve cells in a similar fashion and can be regarded as a shift from micro- to macro-neuronal systems (Figure 1B). Harmful autoimmunity thus has the most extensive consequences for neuronal systems compared with protective and regulatory autoimmunity. Harmful autoimmunity can result from both B or T cell-mediated immunopathology and B cell-directed autoantibodies, such as in membrane-surface autoantibody-based psychiatric autoimmune encephalitis. Thus, damaged autoimmunity means injury to the brain, and can also lead to severe inflammatory conditions therein and in associated neuronal systems.
4. Discussion
Our opinion article shows that autoimmunity based on autoantibodies may play different roles, ranging from physiologic homeostasis to regulatory processes in brain functioning and mental health, as well as building antibody-antigen complexes that might precede severe inflammation and brain damage. These varied expressions of autoimmunity serve to maintain brain homeostasis and are thus part of the “multi-systemic dynamic continuum model” (Figure 1B) in multiple neuronal systems ranging from the nerve cell to neuronal networks. According to this model, autoimmunity mainly evolves to promote brain homeostasis in a dynamic fashion to ensure the brain’s survival and mental health. Keeping these reflections in mind, it is obvious that harmful autoimmunity is also dedicated to maintaining brain function via different neuronal systems. Psychiatric autoimmune disease might therefore serve to help the individual re-adjust, as a manifestation of set-point modulation. Autoimmunity should be put in the context of evolutionary, neurocomputational, and cultural aspects to further investigate its effects in humans as protective or harmful. We believe that taking a philosophical approach helps us elucidate still-unanswered questions about the roles autoantibodies play in psychiatry. In a recent large-scale study [36], 49 different autoantibodies were sought in psychiatric disorders in over 7000 patients. In this study, autoantibodies were detected in 346 of 2043 patients with schizophrenia spectrum disorders and in 62 of 267 patients with affective disorders (suggesting a 17–23% frequency of autoantibodies in affective and psychotic disorders). However, autoantibodies in healthy controls were also detected in 400 of 2748 participants, suggesting a frequency of 15%. This large-scale study shows that autoantibodies are more prevalent in psychiatric disorders than in healthy controls, but autoantibodies per se may not be disease-defining because autoantibodies are also observed in healthy controls. Therefore, it is necessary that signs of CNS inflammation diagnosed via neuroimaging, CSF analysis or functional EEG help us to assess the relevance of these autoantibodies by classifying patient syndromes together with diagnostic findings into disease entities such as psychiatric autoimmune encephalitis [5,7,21] or autoimmune psychosis [6]. In another analysis by Daguano Gastaldi [36], immune checkpoint genotypes were identified as a determining factor for brain autoimmunity and the likelihood of disease development. Other factors may also play a role in determining whether autoantibodies are a relevant phenomenon in the expression of autoimmunity, or whether autoantibodies are merely indicative of brain injury following an immune response in a large proportion of patients. The latter is supported by the finding that a neurotrauma may predispose to a higher seroprevalence of NMDAR antibodies [36]. However, the production and occurrence of humoral autoimmunity is multifactorial, and cannot be proven by a single human autoantibody such as the NMDA antibody. Although the main role of autoimmunity is homeostatic regulation, it can also elicit concomitant psychiatric symptoms, which could in turn lead to a worse outcome. The homeostatic nature of autoimmunity is reflected in part by the relapsing-remitting character of autoantibody-associated psychiatric disorders, or it can even alleviate psychiatric disorders, unlike with progressing neurodegenerative diseases.
Funding
Funding is obtained from the Open access fund of the Georg August University of Göttingen.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The author declares no conflict of interest.
References
- Bien, C.G.; Vincent, A.; Barnett, M.H.; Becker, A.J.; Blümcke, I.; Graus, F.; Jellinger, K.A.; Reuss, D.E.; Ribalta, T.; Schlegel, J.; et al. Immunopathology of autoantibody-associated encephalitides: Clues for pathogenesis. Brain 2012, 135, 1622–1638. [Google Scholar] [CrossRef] [Green Version]
- Malviya, M.; Barman, S.; Golombeck, K.S.; Planagumà, J.; Mannara, F.; Strutz-Seebohm, N.; Wrzos, C.; Demir, F.; Baksmeier, C.; Steckel, J.; et al. NMDAR encephalitis: Passive transfer from man to mouse by a recombinant antibody. Ann. Clin. Transl. Neurol. 2017, 4, 768–783. [Google Scholar] [CrossRef] [Green Version]
- Blackman, G.; Lim, M.F.; Pollak, T.; Al-Diwani, A.; Symmonds, M.; Mazumder, A.; Carter, B.; Irani, S.; David, A. The clinical relevance of serum versus CSF NMDAR autoantibodies associated exclusively with psychiatric features: A systematic review and meta-analysis of individual patient data. J. Neurol. 2022, 269, 5302–5311. [Google Scholar] [CrossRef]
- Ramirez-Franco, J.; Debreux, K.; Extremet, J.; Maulet, Y.; Belghazi, M.; Villard, C.; Sangiardi, M.; Youssouf, F.; El Far, L.; Lévêque, C.; et al. Patient-derived antibodies reveal the subcellular distribution and heterogeneous interactome of LGI1. Brain 2022, 145, 3843–3858. [Google Scholar] [CrossRef]
- Graus, F.; Titulaer, M.J.; Balu, R.; Benseler, S.; Bien, C.G.; Cellucci, T.; Cortese, I.; Dale, R.C.; Gelfand, J.M.; Geschwind, M.; et al. A clinical approach to diagnosis of autoimmune encephalitis. Lancet Neurol. 2016, 15, 391–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollak, T.A.; Lennox, B.R.; Müller, S.; Benros, M.E.; Prüss, H.; van Elst, L.T.; Klein, H.; Steiner, J.; Frodl, T.; Bogerts, B.; et al. Autoimmune psychosis: An international consensus on an approach to the diagnosis and management of psychosis of suspected autoimmune origin. Lancet Psychiatry 2020, 7, 93–108. [Google Scholar] [CrossRef]
- Hansen, N.; Lipp, M.; Vogelgsang, J.; Vukovich, R.; Zindler, T.; Luedecke, D.; Gingele, S.; Malchow, B.; Frieling, H.; Kühn, S.; et al. Autoantibody-associated psychiatric symptoms and syndromes in adults: A narrative review and proposed diagnostic approach. Brain Behav. Immun. Health 2020, 9, 100154. [Google Scholar] [CrossRef]
- Burnet, F.M.; Fenner, F. The Production of Antibodies, 2nd ed.; Macmillian: Melbourne, Australia, 1949. [Google Scholar]
- Pradeu, T.; Carosella, E.D. The Self Model and the Conception of Biological Identity in Immunology. Biol. Philos. 2006, 21, 235–252. [Google Scholar] [CrossRef]
- Burnet, F.M. Immunological recognition of self. Science 1961, 133, 307–311. [Google Scholar] [CrossRef]
- Pradeu, T.; Jaeger, S.; Vivier, E. The speed of change: Towards a discontinuity theory of immunity? Nat. Rev. Immunol. 2013, 13, 764–769. [Google Scholar] [CrossRef]
- Pradeu, T.; Vivier, E. The discontinuity theory of immunity. Sci. Immunol. 2016, 1, aag0479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, M.; Ziv, Y. Immunity to self and self-maintenance: A unified theory of brain pathologies. Trends Immunol. 2008, 29, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.; Raposo, C. Protective Autoimmunity: A Unifying Model for the Immune Network Involved in CNS Repair. Neuroscientist 2014, 20, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Winters, N.C. Autoimmunity and its expression in the analytic situation: A contemporary reflection on our inherent self-destructiveness. Int. J. Psychoanal. 2022, 103, 558–580. [Google Scholar] [CrossRef]
- Hart, B.A.; Luchicchi, A.; Schenk, G.J.; Stys, P.K.; Geurts, J.J.G. Mechanistic underpinning of an inside–out concept for autoimmunity in multiple sclerosis. Ann. Clin. Transl. Neurol. 2021, 8, 1709–1719. [Google Scholar] [CrossRef]
- Kamimura, D.; Yamada, M.; Harada, M.; Sabharwal, L.; Meng, J.; Bando, H.; Ogura, H.; Atsumi, T.; Arima, Y.; Murakami, M. The gateway theory: Bridging neural and immune interactions in the CNS. Front. Neurosci. 2013, 7, 204. [Google Scholar] [CrossRef] [Green Version]
- Nataf, S. Autoimmunity as a Driving Force of Cognitive Evolution. Front. Neurosci. 2017, 11, 582. [Google Scholar] [CrossRef] [Green Version]
- Hansen, N. Current Nosology of Neural Autoantibody-Associated Dementia. Front. Aging Neurosci. 2021, 13, 711195. [Google Scholar] [CrossRef]
- Banks, S.A.; Sechi, E.; Flanagan, E.P. Autoimmune encephalopathies presenting as dementia of subacute onset and rapid progression. Ther. Adv. Neurol. Disord. 2021, 14, 1756286421998906. [Google Scholar] [CrossRef]
- Endres, D.; Lüngen, E.; Hasan, A.; Kluge, M.; Fröhlich, S.; Lewerenz, J.; Bschor, T.; Haußleiter, I.S.; Juckel, G.; Bergh, F.T.; et al. Clinical manifestations and immunomodulatory treatment experiences in psychiatric patients with suspected autoimmune encephalitis: A case series of 91 patients from Germany. Mol. Psychiatry 2022, 27, 1479–1489. [Google Scholar] [CrossRef]
- Hansen, N.; Juhl, A.L.; Grenzer, I.M.; Hirschel, S.; Teegen, B.; Fitzner, D.; Bartels, C.; Timäus, C.; Wiltfang, J.; Malchow, B. Cerebrospinal Fluid Total Tau Protein Correlates With Longitudinal, Progressing Cognitive Dysfunction in Anti-Neural Autoantibody-Associated Dementia and Alzheimer’s Dementia: A Case–Control Study. Front. Immunol. 2022, 13, 837376. [Google Scholar] [CrossRef] [PubMed]
- Menke, A.F.; Ismail, F.S.; Dornmair, K.; Cerina, M.; Meuth, S.G.; Melzer, N. GABAA Receptor Autoantibodies Decrease GABAergic Synaptic Transmission in the Hippocampal CA3 Network. Int. J. Mol. Sci. 2022, 23, 3707. [Google Scholar] [CrossRef] [PubMed]
- Andrzejak, E.; Rabinovitch, E.; Kreye, J.; Prüss, H.; Rosenmund, C.; Ziv, N.E.; Garner, C.C.; Ackermann, F. Patient-Derived Anti-NMDAR Antibody Disinhibits Cortical Neuronal Networks through Dysfunction of Inhibitory Neuron Output. J. Neurosci. 2022, 42, 3253–3270. [Google Scholar] [CrossRef]
- Kayser, M.S.; Dalmau, J. Anti-NMDA receptor encephalitis, autoimmunity, and psychosis. Schizophr. Res. 2016, 176, 36–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coutinho, A. The La Dourarin phenomenon: A shift in the paradigm of developmental self-tolerance. Int. J. Dev. Biol. 2005, 49, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, A. The Network Theory: 21 Years Later. Scand. J. Immunol. 1995, 42, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Notkins, A.L. New Predictors of Disease. Sci. Am. 2007, 296, 72–79. [Google Scholar] [CrossRef]
- Tauber, A.I. Reconceiving autoimmunity: An overview. J. Theor. Biol. 2015, 375, 52–60. [Google Scholar] [CrossRef]
- Savage, P.A.; Klawon, D.E.; Miller, C.H. Regulatory T Cell Development. Annu. Rev. Immunol. 2020, 38, 421–453. [Google Scholar] [CrossRef] [Green Version]
- Liston, A.; Dooley, J.; Yshii, L. Brain-resident regulatory T cells and their role in health and disease. Immunol. Lett. 2022, 248, 26–30. [Google Scholar] [CrossRef]
- Schwarz, A.; Schumacher, M.; Pfaff, D.; Schumacher, K.; Jarius, S.; Balint, B.; Wiendl, H.; Haas, J.; Wildemann, B. Fine-Tuning of Regulatory T Cell Function: The Role of Calcium Signals and Naive Regulatory T Cells for Regulatory T Cell Deficiency in Multiple Sclerosis. J. Immunol. 2013, 190, 4965–4970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Figueroa, P.; Roco, J.A.; Papa, I.; Villacís, L.N.; Stanley, M.; Linterman, M.A.; Dent, A.; Canete, P.F.; Vinuesa, C.G. Follicular regulatory T cells produce neuritin to regulate B cells. Cell 2021, 184, 1775–1789. [Google Scholar] [CrossRef] [PubMed]
- Zöphel, D.; Angenendt, A.; Kaschek, L.; Ravichandran, K.; Hof, C.; Janku, S.; Hoth, M.; Lis, A. Faster cytotoxicity with age: Increased perforin and granzyme levels in cytotoxic CD8+ T cells boost cancer cell elimination. Aging Cell 2022, 21, e13668. [Google Scholar] [CrossRef]
- Nissen, M.S.; Ryding, M.; Nilsson, A.C.; Madsen, J.S.; Olsen, D.A.; Halekoh, U.; Lydolph, M.; Illes, Z.; Blaabjerg, M. CSF-Neurofilament Light Chain Levels in NMDAR and LGI1 Encephalitis: A National Cohort Study. Front. Immunol. 2021, 12, 719432. [Google Scholar] [CrossRef] [PubMed]
- Gastaldi, V.D.; Wilke, J.B.; Weidinger, C.A.; Walter, C.; Barnkothe, N.; Teegen, B.; Luessi, F.; Stöcker, W.; Lühder, F.; Begemann, M.; et al. Factors predisposing to humoral autoimmunity against brain-antigens in health and disease: Analysis of 49 autoantibodies in over 7000 subjects. Brain Behav. Immun. 2022, 108, 135–147. [Google Scholar] [CrossRef] [PubMed]
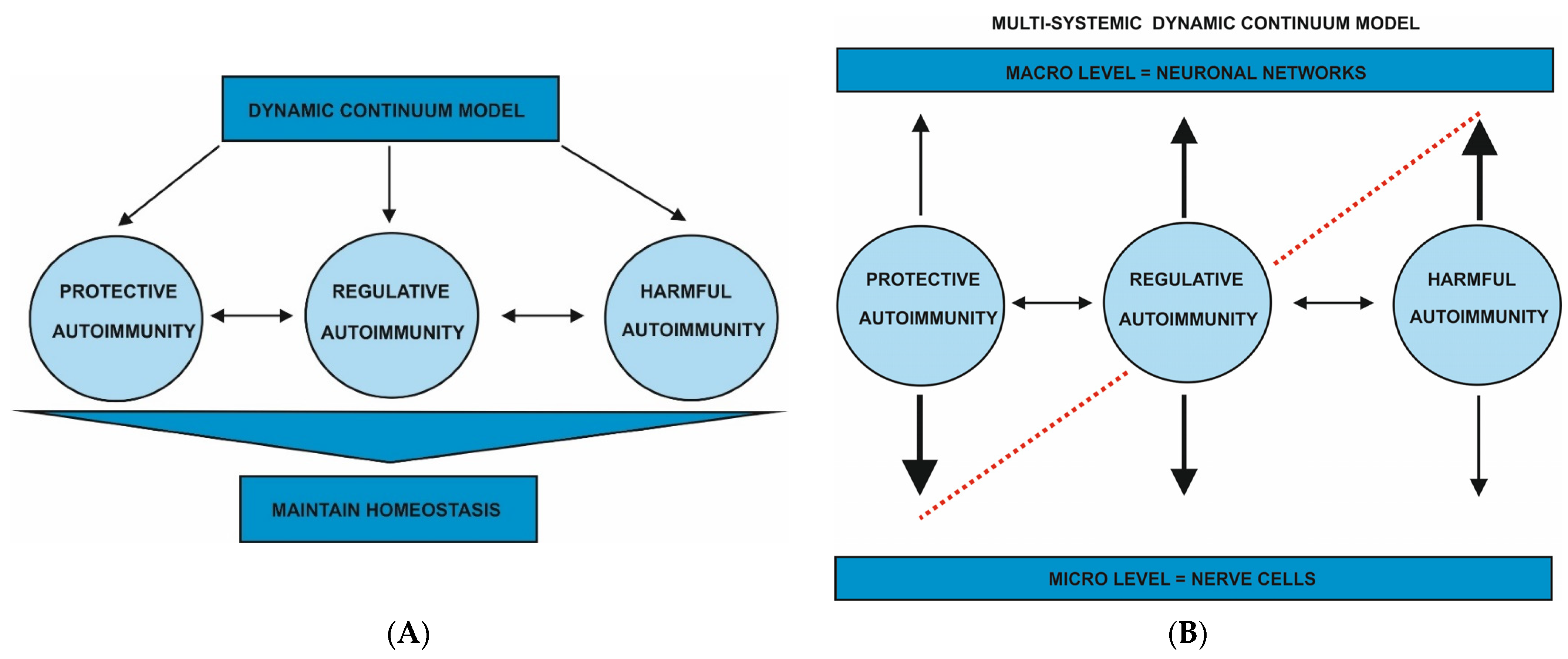
Figure 1.
(A) Dynamic continuum model of autoimmunity. The dynamic continuum model of autoimmunity is depicted in Figure 1, and ranges from protective and regulatory to harmful forms of autoimmunity, which all serve to maintain brain homeostasis and thus mental health. (B) Dynamic continuum model of autoimmunity categorized into neuronal systems. The neuronal systems that are affected by the postulated three types of autoimmunity are the nerve cell on a micro level and neuronal networks on a macro level. Each type of autoimmunity differently affects these neuronal systems, so that a shift more affecting a micro level in protective autoimmunity can have an effect on the macro level with more consequences of harmful autoimmunity for neuronal networks can be observed (multi-systemic dynamic continuum model). The dashed line represents potential imbalance if one form of autoimmunity increases. For example, when harmful autoimmunity increases, protective autoimmunity decreases. When regulatory autoimmunity predominates, harmful and protective autoimmunity are equal. The increase in one form of autoimmunity is additionally indicated by the arrow’s size: a larger arrow indicates this form of autoimmunity’s predominance.
Figure 1.
(A) Dynamic continuum model of autoimmunity. The dynamic continuum model of autoimmunity is depicted in Figure 1, and ranges from protective and regulatory to harmful forms of autoimmunity, which all serve to maintain brain homeostasis and thus mental health. (B) Dynamic continuum model of autoimmunity categorized into neuronal systems. The neuronal systems that are affected by the postulated three types of autoimmunity are the nerve cell on a micro level and neuronal networks on a macro level. Each type of autoimmunity differently affects these neuronal systems, so that a shift more affecting a micro level in protective autoimmunity can have an effect on the macro level with more consequences of harmful autoimmunity for neuronal networks can be observed (multi-systemic dynamic continuum model). The dashed line represents potential imbalance if one form of autoimmunity increases. For example, when harmful autoimmunity increases, protective autoimmunity decreases. When regulatory autoimmunity predominates, harmful and protective autoimmunity are equal. The increase in one form of autoimmunity is additionally indicated by the arrow’s size: a larger arrow indicates this form of autoimmunity’s predominance.


{kind=link}
Table 1.
Current concepts of CNS autoimmunity—their advantages and disadvantages.
| Theory | Advantage | Disadvantage | Reference |
|---|---|---|---|
| Self/non-self model |
|
| [8] |
| Continuity model |
|
| [9] |
| Unified theory of brain pathologies |
|
| [13] |
| Unifying model for immune network for CNS repair functions |
|
| [14] |
| Inherent self-destructiveness |
|
| [15] |
| Inside-out concept of autoimmunity |
|
| [16] |
| Gateway theory |
|
| [17] |
| Force of cognitive evolution |
|
| [18] |
| Multi-systemic dynamic continuum model |
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hansen, N. Philosophical Approach to Neural Autoantibodies in Psychiatric Disease—Multi-Systemic Dynamic Continuum from Protective to Harmful Autoimmunity in Neuronal Systems. Antibodies 2023, 12, 1. https://doi.org/10.3390/antib12010001
AMA Style
Hansen N. Philosophical Approach to Neural Autoantibodies in Psychiatric Disease—Multi-Systemic Dynamic Continuum from Protective to Harmful Autoimmunity in Neuronal Systems. Antibodies. 2023; 12(1):1. https://doi.org/10.3390/antib12010001
Chicago/Turabian StyleHansen, Niels. 2023. "Philosophical Approach to Neural Autoantibodies in Psychiatric Disease—Multi-Systemic Dynamic Continuum from Protective to Harmful Autoimmunity in Neuronal Systems" Antibodies 12, no. 1: 1. https://doi.org/10.3390/antib12010001
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.