Forecasting Seasonal Habitat Connectivity in a Developing Landscape


Abstract
:1. Introduction
2. Data and Methods
2.1. Black Bear Data
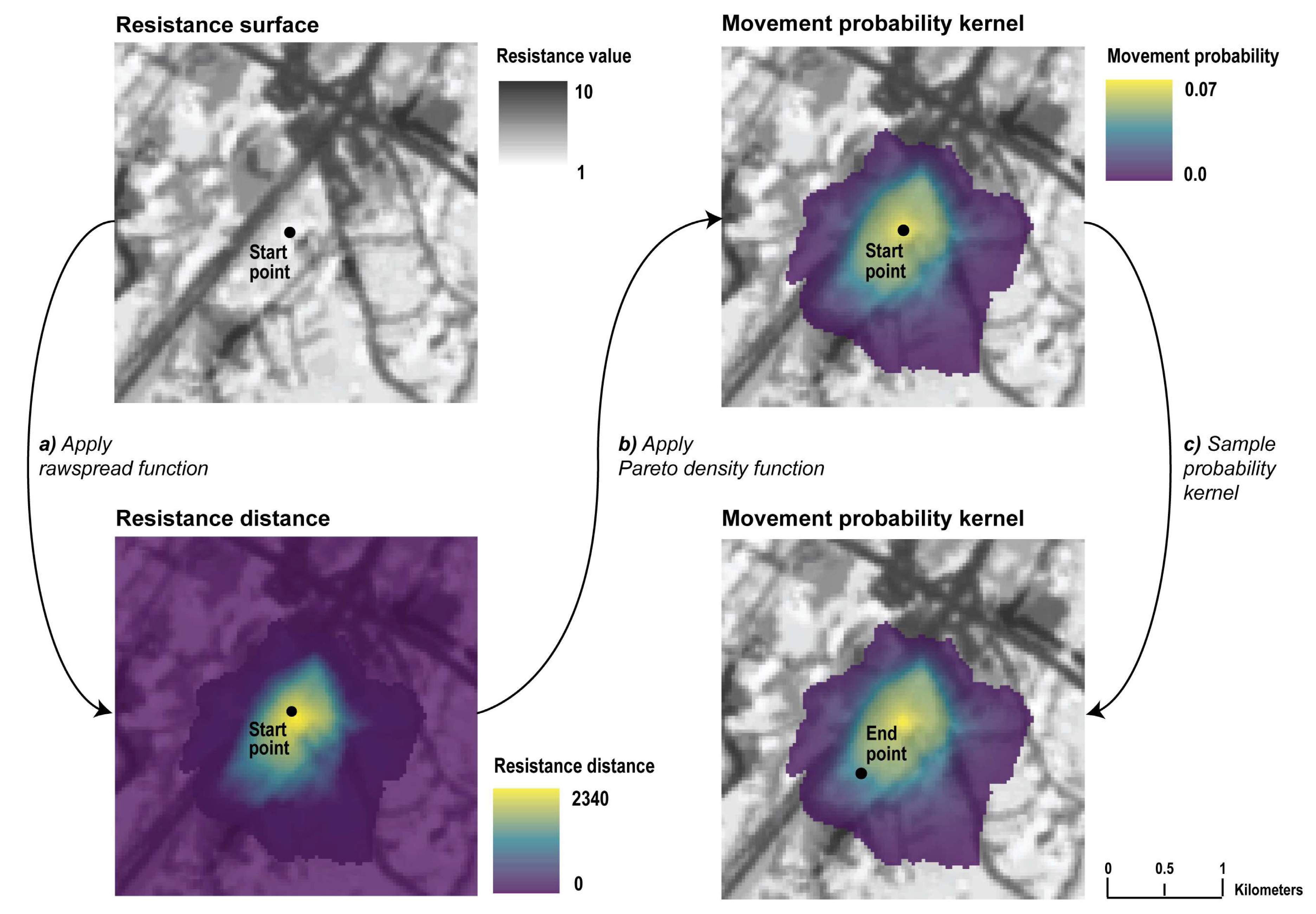
2.2. Resistance Surfaces
2.3. Step Length Distributions
2.4. Individual-based Movement Model
2.5. Model Validation and Calibration
2.6. Future Projections
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Year | Full Date Range of Data | No. Fixes Cleaned and Outside Hibernation Period | No. Steps Spring | No. Steps Summer | No. Steps Fall | Age of Young | |
|---|---|---|---|---|---|---|---|---|
| Start | End | |||||||
| 234 | 2012 | 2012.02.22 | 2013.03.11 | 3521 | 502 | 708 | yearling | |
| 310 | 2012 | 2012.03.28 | 2013.02.11 | 4039 | 462 | 622 | 940 | newborn |
| 258 | 2012 | 2012.03.14 | 2013.02.08 | 3668 | 876 | 216 | 998 | yearling |
| 253 | 2012 | 2012.03.09 | 2013.02.07 | 4186 | 320 | 574 | 1068 | newborn |
| 323 | 2012 | 2012.03.21 | 2013.03.01 | 3770 | 748 | 480 | 608 | yearling |
| 258 | 2013 | 2013.03.17 | 2014.02.27 | 3277 | 338 | 998 | newborn | |
| 298 | 2013 | 2013.04.02 | 2014.03.25 | 6503 | 228 | 860 | 2204 | newborn |
| 388 | 2013 | 2013.02.21 | 2013.09.11 | 4262 | 868 | 944 | yearling | |
| 393 | 2013 | 2013.05.09 | 2014.02.24 | 3624 | 228 | 750 | newborn | |
| 310 | 2013 | 2013.02.15 | 2014.03.07 | 4743 | 586 | 386 | 492 | yearling |
| 395 | 2013 | 2013.05.21 | 2014.01.19 | 3726 | 1234 | 912 | newborn | |
| 391 | 2013 | 2013.05.02 | 2014.01.17 | 2786 | 668 | 606 | none | |
| 356 | 2013 | 2013.05.01 | 2014.03.17 | 4410 | 1196 | 982 | none | |
| 253 | 2013 | 2013.02.23 | 2014.03.06 | 5845 | 944 | 1284 | 1266 | yearling |
| 370 | 2013 | 2013.03.06 | 2014.02.28 | 4475 | 286 | 598 | 1306 | newborn |
| 323 | 2013 | 2013.03.02 | 2013.09.09 | 2307 | 510 | 504 | none | |
| 355 | 2013 | 2013.03.01 | 2014.03.10 | 3452 | 686 | 622 | 992 | newborn |
| 373 | 2013 | 2013.03.05 | 2014.03.14 | 2727 | 286 | 598 | 1306 | none |
| 393 | 2014 | 2014.02.22 | 2015.03.02 | 4239 | 1072 | 748 | 542 | yearling |
| 298 | 2014 | 2014.03.26 | 2014.06.17 | 1065 | 552 | yearling | ||
| 406 | 2014 | 2014.06.19 | 2015.02.12 | 2114 | 482 | Unknown | ||
| 258 | 2014 | 2014.02.28 | 2015.02.17 | 3665 | 440 | 338 | 120 | yearling |
| 391 | 2014 | 2014.03.11 | 2015.02.10 | 3805 | 746 | 596 | none | |
| 356 | 2014 | 2014.02.26 | 2015.01.08 | 5024 | 426 | 1214 | 966 | newborn |
| 373 | 2014 | 2014.03.15 | 2014.11.01 | 3365 | 816 | 466 | none | |
| 269 | 2014 | 2014.02.24 | 2015.03.02 | 3481 | 478 | yearling | ||
| 355 | 2014 | 2014.03.10 | 2015.01.28 | 4693 | 524 | 756 | 1284 | newborn |
| 370 | 2014 | 2014.03.01 | 2015.01.01 | 3271 | 816 | 466 | yearling | |
| 406 | 2015 | 2015.03.11 | 2016.01.05 | 4102 | 346 | 496 | 1434 | newborn |
| 426 | 2015 | 2015.05.30 | 2016.01.04 | 3647 | 550 | 1228 | newborn | |
| 393 | 2015 | 2015.03.10 | 2015.12.01 | 4043 | 804 | 532 | newborn | |
| 391 | 2015 | 2015.03.02 | 2016.01.09 | 5003 | 386 | 522 | 1380 | newborn |
| 395 | 2015 | 2015.03.05 | 2016.01.19 | 5153 | 436 | 724 | 1504 | newborn |
| 403 | 2015 | 2015.02.19 | 2015.11.23 | 2973 | 518 | 392 | none | |
| 356 | 2015 | 2015.03.06 | 2015.12.13 | 3318 | 156 | 526 | 518 | newborn |
| 404 | 2015 | 2015.02.12 | 2015.06.18 | 1526 | 800 | none | ||
| 432 | 2015 | 2015.07.15 | 2015.12.14 | 2594 | 1026 | newborn | ||
| 428 | 2015 | 2015.05.01 | 2016.01.04 | 2590 | 668 | none | ||
| 425 | 2015 | 2015.05.30 | 2016.01.04 | 2110 | 352 | newborn | ||
| 373 | 2015 | 2015.03.16 | 2015.08.25 | 2632 | 540 | 580 | newborn | |
| 269 | 2015 | 2015.03.13 | 2015.11.10 | 3176 | 336 | 1052 | newborn | |
| 355 | 2015 | 2015.01.29 | 2015.12.02 | 802 | 766 | 488 | yearling | |
| 436 | 2016 | 2016.03.22 | 2016.12.06 | 4492 | 440 | 1006 | newborn | |
| 425 | 2016 | 2016.03.09 | 2016.12.13 | 5025 | 586 | 1156 | newborn | |
| 432 | 2016 | 2016.02.16 | 2017.01.23 | 3752 | 758 | 416 | yearling | |
| 424 | 2016 | 2016.03.11 | 2016.12.27 | 4320 | 416 | 808 | 1242 | newborn |
| 356 | 2016 | 2015.12.21 | 2106.12.06 | 3287 | 204 | 350 | 624 | newborn |
| 395 | 2016 | 2016.01.19 | 2016.12.20 | 4515 | 1302 | 878 | 1504 | yearling |
| 406 | 2016 | 2016.01.05 | 2016.12.06 | 4185 | 954 | 524 | 700 | yearling |
| 426 | 2016 | 2016.02.01 | 2016.12.26 | 4883 | 1190 | yearling | ||
| 433 | 2017 | 2017.02.03 | 2017.11.27 | 4211 | 670 | 704 | 604 | none |
| 426 | 2017 | 2017.03.09 | 2017.11.28 | 4814 | 518 | newborn | ||
| 450 | 2017 | 2017.02.23 | 2017.11.14 | 4024 | 386 | none | ||
| 476 | 2017 | 2017.07.06 | 2017.12.21 | 3092 | 1528 | none | ||
| 309 | 2017 | 2017.03.10 | 2017.12.01 | 5052 | 534 | 1400 | newborn | |
| 470 | 2017 | 2017.06.16 | 2018.01.07 | 2410 | 392 | none | ||
| 424 | 2017 | 2017.02.24 | 2017.12.04 | 4415 | 694 | 848 | yearling | |
| 472 | 2017 | 2017.06.20 | 2017.12.18 | 3186 | 822 | 1412 | none | |
| 432 | 2017 | 2017.03.06 | 2017.12.05 | 5270 | 466 | 882 | newborn | |
| 425 | 2017 | 2016.12.12 | 2017.11.14 | 4724 | 990 | 848 | 860 | yearling |
| 436 | 2017 | 2016.11.15 | 2018.01.02 | 4823 | 480 | 740 | 1206 | yearling |
| 465 | 2017 | 2017.05.27 | 2017.11.14 | 3335 | 856 | 754 | yearling | |
| 451 | 2017 | 2017.03.22 | 2017.12.09 | 5842 | 598 | 1930 | newborn | |
| 445 | 2017 | 2017.03.30 | 2017.11.07 | 3743 | 604 | 448 | yearling | |
| 471 | 2017 | 2017.06.17 | 2018.01.02 | 3435 | 708 | newborn | ||
Appendix B

Appendix C

Appendix D
Appendix E
| Mean Step Length | Mean Home Range Size | |
|---|---|---|
| Empirical bears | 193.34 m (Range: 0–1998 m) | 62.51 km2 (Range: 5.47–260.55 km2) |
| Percent of Pareto distribution scale parameter | ||
| 0 | 73.29 m (Range: 0–902 m) | 44.38 km2 (Range: 10.82–110.31 km2) |
| 10 | 82.53 m (Range: 0–2249 m) | 77.97 km2 (Range: 6.85–228.44 km2) |
| 20 | 94.73 m (Range: 0–1757 m) | 104.74 km2 (Range: 29.75–231.53 km2) |
References
- Rudnick, D.; Beier, P.; Cushman, S.; Dieffenbach, F.; Epps, C.W.; Gerber, L.; Hartter, J.; Jenness, J.; Kintsch, J.; Merenlender, A.M.; et al. The Role of Landscape Connectivity in Planning and Implementing Conservation and Restoration Priorities; Issues in Ecology. Report No. 16.; Ecological Society of America: Washington, DC, USA, 2012. [Google Scholar]
- Hilty, J.A.; Keeley, A.T.H.; Lidicker, W.Z., Jr.; Merenlender, A.M. Corridor Ecology, 2nd ed.: Linking Landscapes for Biodiversity Conservation and Climate Adaptation; Island Press: Washington, DC, USA, 2019. [Google Scholar]
- Chen, I.-C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid Range Shifts of Species Associated with High Levels of Climate Warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Keeley, A.T.H.; Beier, P.; Creech, T.; Jones, K.; Jongman, R.H.; Stonecipher, G.; Tabor, G.M. Thirty years of connectivity conservation planning: An assessment of factors influencing plan implementation. Environ. Res. Lett. 2019, 14, 103001. [Google Scholar] [CrossRef]
- Saura, S.; Bertzky, B.; Bastin, L.; Battistella, L.; Mandrici, A.; Dubois, G. Protected area connectivity: Shortfalls in global targets and country-level priorities. Biol. Conserv. 2018, 219, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Koen, E.L.; Bowman, J.; Sadowski, C.; Walpole, A.A. Landscape connectivity for wildlife: Development and validation of multispecies linkage maps. Methods Ecol. Evol. 2014, 5, 626–633. [Google Scholar] [CrossRef]
- Proctor, M.F.; Nielsen, S.E.; Kasworm, W.F.; Servheen, C.; Radandt, T.G.; MacHutchon, A.G.; Boyce, M.S. Grizzly bear connectivity mapping in the Canada-United States trans-border region. J. Wildl. Manag. 2015, 79, 544–558. [Google Scholar] [CrossRef]
- Mueller, T.; Olson, K.A.; Fuller, T.K.; Schaller, G.B.; Murray, M.G.; Leimgruber, P. In search of forage: Predicting dynamic habitats of Mongolian gazelles using satellite-based estimates of vegetation productivity. J. Appl. Ecol. 2008, 45, 649–658. [Google Scholar] [CrossRef]
- Bélisle, M. Measuring Landscape Connectivity: The Challenge of Behavioral Landscape Ecology. Ecology 2005, 86, 1988–1995. [Google Scholar] [CrossRef] [Green Version]
- Osipova, L.; Okello, M.M.; Njumbi, S.J.; Ngene, S.; Western, D.; Hayward, M.; Balkenhol, N. Using step-selection functions to model landscape connectivity for African elephants: Accounting for variability across individuals and seasons. Anim. Conserv. 2018, 22, 35–48. [Google Scholar] [CrossRef]
- Zeller, K.A.; Wattles, D.W.; Conlee, L.; DeStefano, S. Black bears alter movements in response to anthropogenic features with time of day and season. Mov. Ecol. 2019, 7, 19. [Google Scholar] [CrossRef]
- Wolf, M.; Weissing, F.J. Animal personalities: Consequences for ecology and evolution. Trends Ecol. Evol. 2012, 27, 452–461. [Google Scholar] [CrossRef]
- Chetkiewicz, C.-L.B.; Boyce, M.S. Use of resource selection functions to identify conservation corridors. J. Appl. Ecol. 2009, 46, 1036–1047. [Google Scholar] [CrossRef]
- Parks, S.A.; Carroll, C.; Dobrowski, S.Z.; Allred, B.W. Human land uses reduce climate connectivity across North America. Glob. Chang. Biol. 2020, 26, 2944–2955. [Google Scholar] [CrossRef] [PubMed]
- Kool, J.; Moilanen, A.; Treml, E.A. Population connectivity: Recent advances and new perspectives. Landsc. Ecol. 2012, 28, 165–185. [Google Scholar] [CrossRef]
- Martensen, A.C.; Saura, S.; Fortin, M.-J. Spatio-temporal connectivity: Assessing the amount of reachable habitat in dynamic landscapes. Methods Ecol. Evol. 2017, 8, 1253–1264. [Google Scholar] [CrossRef]
- Wimberly, M.C. Species Dynamics in Disturbed Landscapes: When does a Shifting Habitat Mosaic Enhance Connectivity? Landsc. Ecol. 2006, 21, 35–46. [Google Scholar] [CrossRef]
- Hodgson, J.; Moilanen, A.; Thomas, C.D. Metapopulation responses to patch connectivity and quality are masked by successional habitat dynamics. Ecology 2009, 90, 1608–1619. [Google Scholar] [CrossRef] [Green Version]
- Roe, J.H.; Brinton, A.C.; Georges, A. Temporal and spatial variation in landscape connectivity for a freshwater turtle in a temporally dynamic wetland system. Ecol. Appl. 2009, 19, 1288–1299. [Google Scholar] [CrossRef]
- Adriaensen, F.; Chardon, J.; De Blust, G.; Swinnen, E.; Villalba, S.; Gulinck, H.; Matthysen, E. The application of ‘least-cost’ modelling as a functional landscape model. Landsc. Urban Plan. 2003, 64, 233–247. [Google Scholar] [CrossRef]
- Loro, M.; Ortega, E.; Arce, R.M.; Geneletti, D. Ecological connectivity analysis to reduce the barrier effect of roads. An innovative graph-theory approach to define wildlife corridors with multiple paths and without bottlenecks. Landsc. Urban Plan. 2015, 139, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Compton, B.W.; McGarigal, K.; Cushman, S.A.; Gamble, L.R. A Resistant-Kernel Model of Connectivity for Amphibians that Breed in Vernal Pools. Conserv. Biol. 2007, 21, 788–799. [Google Scholar] [CrossRef]
- McRae, B.H.; Dickson, B.; Keitt, T.H.; Shah, V.B. Using Circuit Theory to Model Connectivity in Ecology, Evolution, and Conservation. Ecology 2008, 89, 2712–2724. [Google Scholar] [CrossRef] [PubMed]
- McRae, B.H.; Hall, S.A.; Beier, P.; Theobald, D.M. Where to Restore Ecological Connectivity? Detecting Barriers and Quantifying Restoration Benefits. PLoS ONE 2012, 7, e52604. [Google Scholar] [CrossRef]
- Kool, J.; Paris, C.B.; Barber, P.H.; Cowen, R.K. Connectivity and the development of population genetic structure in Indo-West Pacific coral reef communities. Glob. Ecol. Biogeogr. 2011, 20, 695–706. [Google Scholar] [CrossRef]
- Hauenstein, S.; Fattebert, J.; Grüebler, M.U.; Naef-Daenzer, B.; Pe’er, G.; Hartig, F. Calibrating an individual-based movement model to predict functional connectivity for little owls. Ecol. Appl. 2019, 29, e01873. [Google Scholar] [CrossRef] [PubMed]
- Mui, A.; Caverhill, B.; Johnson, B.; Fortin, M.-J.; He, Y. Using multiple metrics to estimate seasonal landscape connectivity for Blanding’s turtles (Emydoidea blandingii) in a fragmented landscape. Landsc. Ecol. 2016, 32, 531–546. [Google Scholar] [CrossRef]
- Grimm, V.; Railsback, S.F. Individual-based Modeling and Ecology; Princeton University Press: Princeton, NJ, USA, 2005. [Google Scholar]
- McLane, A.J.; Semeniuk, C.; McDermid, G.J.; Marceau, D.J. The role of agent-based models in wildlife ecology and management. Ecol. Model. 2011, 222, 1544–1556. [Google Scholar] [CrossRef]
- Wood, K.A.; Stillman, R.A.; Hilton, G.M. Conservation in a changing world needs predictive models. Anim. Conserv. 2017, 21, 87–88. [Google Scholar] [CrossRef] [Green Version]
- Kramer-Schadt, S.; Revilla, E.; Wiegand, T. Lynx reintroductions in fragmented landscapes of Germany: Projects with a future or misunderstood wildlife conservation? Biol. Conserv. 2005, 125, 169–182. [Google Scholar] [CrossRef]
- DeAngelis, D.L.; Grimm, V. Individual-based models in ecology after four decades. F1000Prime Rep. 2014, 6. [Google Scholar] [CrossRef] [Green Version]
- Bauduin, S.; McIntire, E.; St-Laurent, M.-H.; Cumming, S. Overcoming challenges of sparse telemetry data to estimate caribou movement. Ecol. Model. 2016, 335, 24–34. [Google Scholar] [CrossRef]
- Wiegand, T.; Jeltsch, F.; Hanski, I.; Grimm, V. Using pattern-oriented modeling for revealing hidden information: A key for reconciling ecological theory and application. Oikos 2003, 100, 209–222. [Google Scholar] [CrossRef] [Green Version]
- Pe’Er, G.; Kramer-Schadt, S. Incorporating the perceptual range of animals into connectivity models. Ecol. Model. 2008, 213, 73–85. [Google Scholar] [CrossRef]
- R Core Team. R: A language and Environment for Statistical Computing. Version 3.6.2. 2019. Available online: https://cran.r-project.org/ (accessed on 6 December 2019).
- Lewis, J.S.; Rachlow, J.L.; Garton, E.O.; Vierling, L.A. Effects of habitat on GPS collar performance: Using data screening to reduce location error. J. Appl. Ecol. 2007, 44, 663–671. [Google Scholar] [CrossRef]
- Telonics. Gen 4 GPS Systems Manual. Telonics, Inc., 2012. Available online: www.telonics.com. (accessed on 1 March 2018).
- Estrada, E.G.; Alva, J.A.V. Gpdtest: Bootstrap Goodness-Of-Fit Test for the Generalized Pareto Distribution. Version 0.4. 2012. Available online: https://cran.r-project.org/web/packages/gPdtest/ (accessed on 26 April 2019).
- Plunkett, E.B. Gridprocess: Package for Processing Raster Data. Version 0.1.3. 2019. Available online: https://github.com/ethanplunkett/gridprocess (accessed on 4 June 2019).
- Tillé, Y.; Matei, A. Sampling: Survey Sampling. Version 2.8. 2016. Available online: https://CRAN.R-project.org/package=sampling (accessed on 22 September 2016).
- Grimm, V.; Berger, U.; Bastiansen, F.; Eliassen, S.; Ginot, V.; Giske, J.; Goss-Custard, J.; Grand, T.; Heinz, S.K.; Huse, G.; et al. A standard protocol for describing individual-based and agent-based models. Ecol. Model. 2006, 198, 115–126. [Google Scholar] [CrossRef]
- Grimm, V.; Berger, U.; DeAngelis, D.L.; Polhill, J.G.; Giske, J.; Railsback, S.F.; Polhill, N.M.G.J.G. The ODD protocol: A review and first update. Ecol. Model. 2010, 221, 2760–2768. [Google Scholar] [CrossRef] [Green Version]
- McGarigal, K.; Compton, B.W.; Plunkett, E.B.; DeLuca, W.V.; Grand, J. Designing sustainable landscapes project. University of Massachusetts, Amherst. 2017. Available online: www.umass.edu/landeco/research/dsl/dsl.html (accessed on 10 March 2020).
- McGarigal, K.; Plunkett, E.B.; Willey, L.L.; Compton, B.W.; DeLuca, W.; Grand, J. Modeling non-stationary urban growth: The SPRAWL model and the ecological impacts of development. Landsc. Urban Plan. 2018, 177, 178–190. [Google Scholar] [CrossRef]
- UMass Donahue Institute. Population Projections. UMass Donahue Institute, MassDOT Vintage. 2018. Available online: pep.donahue-institute.org (accessed on 7 April 2020).
- Pe’er, G.; Henle, K.; Dislich, C.; Frank, K. Breaking Functional Connectivity into Components: A Novel Approach Using an Individual-Based Model, and First Outcomes. PLoS ONE 2011, 6, e22355. [Google Scholar]
- Allen, C.H.; Parrott, L.; Kyle, C. An individual-based modelling approach to estimate landscape connectivity for bighorn sheep (Ovis canadensis). PeerJ 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Costello, C.M. Estimates of dispersal and home-range fidelity in American black bears. J. Mammal. 2010, 91, 116–121. [Google Scholar] [CrossRef]
- Rogers, L.L. Effects of Food Supply and Kinship on Social Behavior, Movements, and Population Growth of Black Bears in Northeastern Minnesota. Wildl. Monogr. 1987, 97, 3–72. [Google Scholar]
- Kristensen, T.V.; Puckett, E.E.; Landguth, E.L.; Belant, J.L.; Hast, J.T.; Carpenter, C.; Sajecki, J.L.; Beringer, J.; Means, M.; Cox, J.J.; et al. Spatial genetic structure in American black bears (Ursus americanus): Female philopatry is variable and related to population history. Heredity 2017, 120, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Avgar, T.; Deardon, R.; Fryxell, J.M. An empirically parameterized individual based model of animal movement, perception, and memory. Ecol. Model. 2013, 251, 158–172. [Google Scholar] [CrossRef]
- Marley, J.; Hyde, A.; Salkeld, J.H.; Prima, M.-C.; Parrott, L.; Senger, S.E.; Tyson, R.C. Does human education reduce conflicts between humans and bears? An agent-based modelling approach. Ecol. Model. 2017, 343, 15–24. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeller, K.A.; Wattles, D.W.; Bauder, J.M.; DeStefano, S. Forecasting Seasonal Habitat Connectivity in a Developing Landscape. Land 2020, 9, 233. https://doi.org/10.3390/land9070233
Zeller KA, Wattles DW, Bauder JM, DeStefano S. Forecasting Seasonal Habitat Connectivity in a Developing Landscape. Land. 2020; 9(7):233. https://doi.org/10.3390/land9070233
Chicago/Turabian StyleZeller, Katherine A., David W. Wattles, Javan M. Bauder, and Stephen DeStefano. 2020. "Forecasting Seasonal Habitat Connectivity in a Developing Landscape" Land 9, no. 7: 233. https://doi.org/10.3390/land9070233