Gendered Species Preferences Link Tree Diversity and Carbon Stocks in Cacao Agroforest in Southeast Sulawesi, Indonesia

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Plot Measurements
2.3. C stock Estimation
2.4. Farmer Preferences for Shade Tree Species
2.5. Compilation of C Stocks and biodiversity Data from Previous Research
2.6. Data Analysis
3. Results
3.1. Tree Diversity
3.2. Vegetation Structure
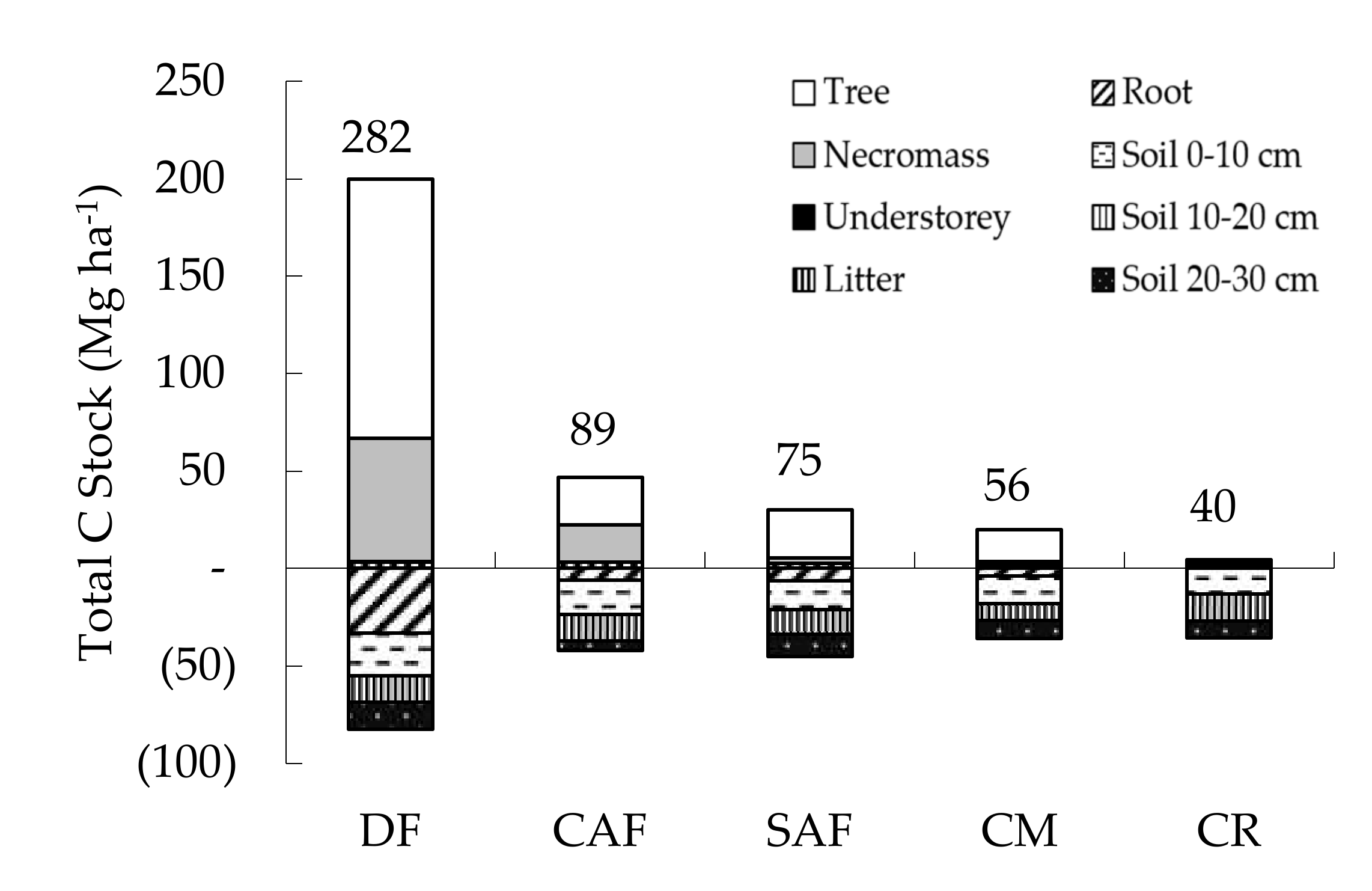
3.3. Tree Biomass, Necromass and C Stocks
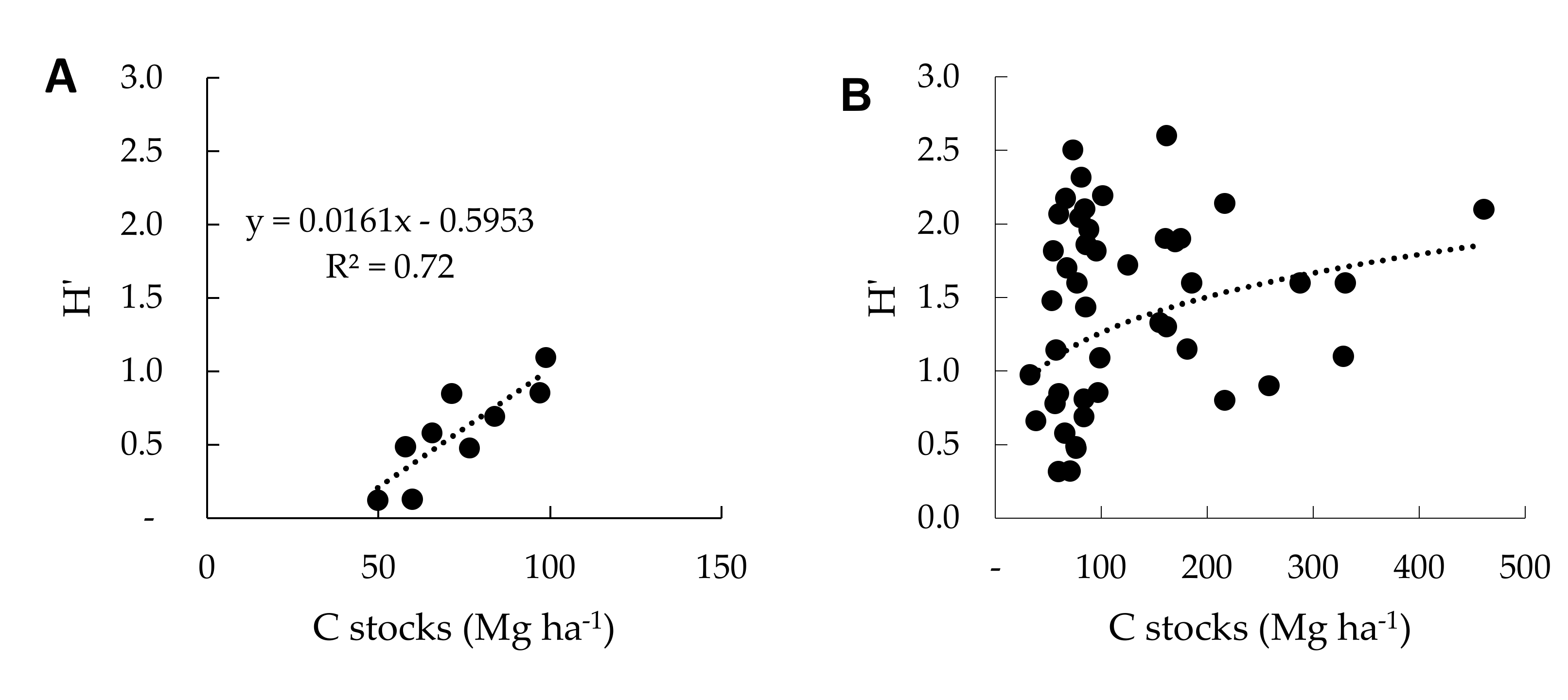
3.4. Relationships between C Stocks and Tree Diversity in Agroforestry Systems
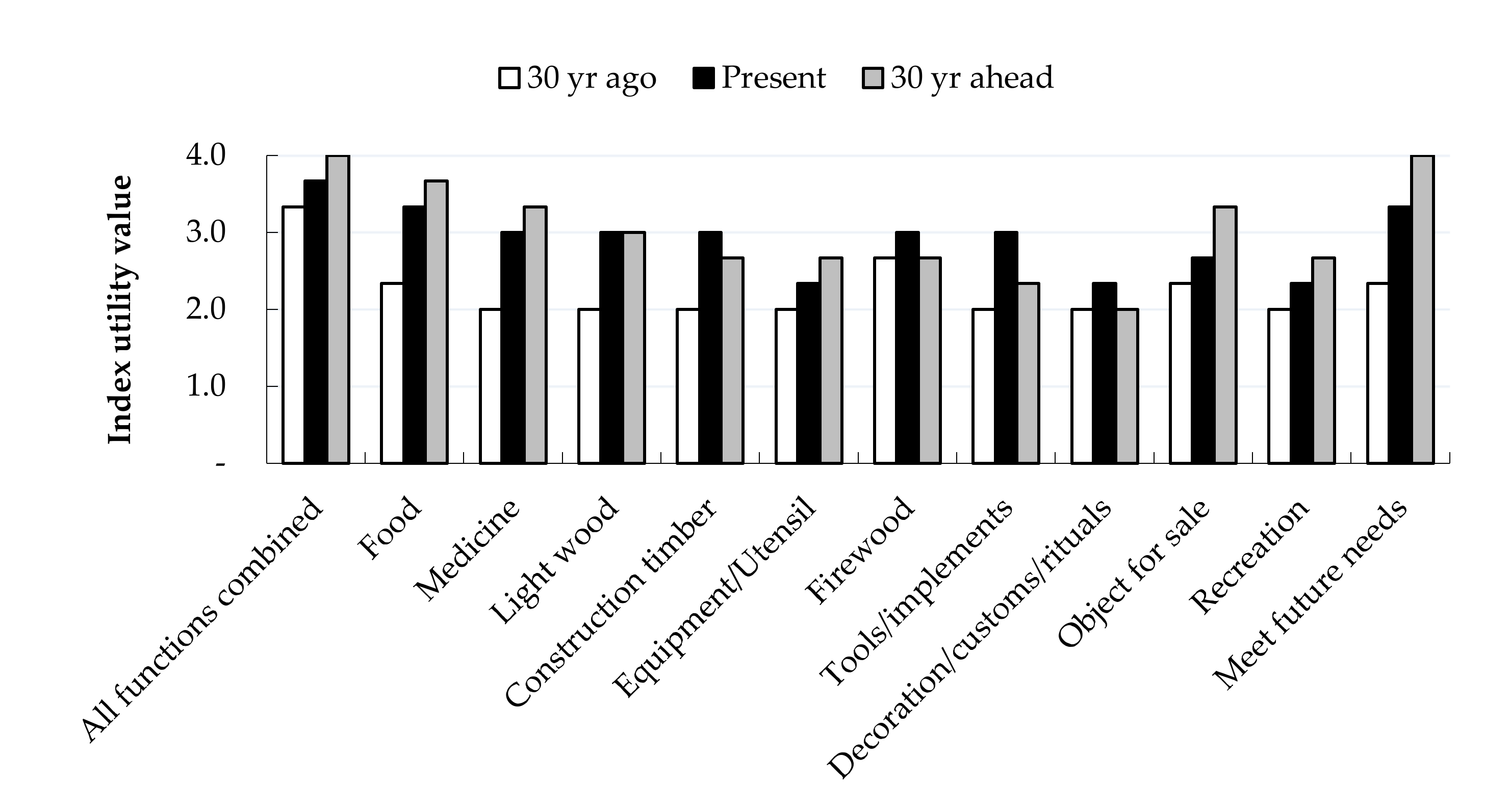
3.5. Farmer Tree Preferences and Utility Value of Cacao-based Agroforestry Systems
4. Discussion
4.1. Land Use Affects Tree Diversity and C Stocks
4.2. Gendered Tree Preferences Increased Diversity, C Stocks and the Utility Value of Agroforestry Systems
4.3. Relationships between C Stocks and Tree Diversity within and Across Land-use Systems
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van Noordwijk, M. Integrated Natural Resource Management as Pathway to Poverty Reduction: Innovating Practices, Institutions and Policies. Agric. Syst. 2019, 172, 60–71. [Google Scholar] [CrossRef]
- Barrios, E.; Valencia, V.; Jonsson, M.; Brauman, A.; Hairiah, K.; Mortimer, P.E.; Okubo, S. Contribution of Trees to the Conservation of Biodiversity and Ecosystem Services in Agricultural Landscapes. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2018, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Cavanaugh, K.C.; Gosnell, J.S.; Davis, S.L.; Ahumada, J.; Boundja, P.; Clark, D.B.; Mugerwa, B.; Jansen, P.A.; O’Brien, T.G.; Rovero, F.; et al. Carbon storage in tropical forests correlates with taxonomic diversity and functional dominance on a global scale. Glob. Ecol. Biogeogr. 2014, 23, 563–573. [Google Scholar] [CrossRef]
- Van de Perre, F.; Willig, M.R.; Presley, S.J.; Andemwana, F.B.; Beeckman, H.; Boeckx, P.; Cooleman, S.; de Haan, M.; De Kesel, A.; Dessein, S.; et al. Reconciling biodiversity and carbon stock conservation in an Afrotropical forest landscape. Sci. Adv. 2018, 4, eaar6603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loreau, M.; Hector, A. Partitioning selection and complementarity in biodiversity experiments. Nature 2001, 412, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Hicks, C.; Woroniecki, S.; Fancourt, M.; Bieri, M.; Garcia Robles, H.; Trumper, K.; Mant, R. The Relationship between Biodiversity, Carbon Storage and the Provision of other Ecosystem Services: Critical Review for the Forestry Component of the International Climate Fund; Department for International Development: Cambridge, UK, 2014; p. 119. [Google Scholar]
- Cardinale, B.J.; Matulich, K.L.; Hooper, D.U.; Byrnes, J.E.; Duffy, E.; Gamfeldt, L.; Balvanera, P.; O’Connor, M.I.; Gonzalez, A. The functional role of producer diversity in ecosystems. Am. J. Bot. 2011, 98, 572–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swift, M.J.; Izac, A.M.N.; van Noordwijk, M. Biodiversity and ecosystem services in agricultural landscapes—Are we asking the right questions? Agric. Ecosyst. Environ. 2004, 104, 113–134. [Google Scholar] [CrossRef]
- Lecina-Diaz, J.; Alvarez, A.; Regos, A.; Drapeau, P.; Paquette, A.; Messier, C.; Retana, J. The positive carbon stocks-biodiversity relationship in forests: Co-occurrence and drivers across five subclimates. Ecol. Appl. 2018, 28, 1481–1493. [Google Scholar] [CrossRef]
- Healy, C.; Gotelli, N.J.; Potvin, C. Partitioning the effects of biodiversity and environmental heterogeneity for productivity and mortality in a tropical tree plantation. J. Ecol. 2008, 96, 903–913. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Talbot, J.; Lewis, S.L.; Phillips, O.L.; Qie, L.; Begne, S.K.; Chave, J.; Cuni-Sanchez, A.; Hubau, W.; Lopez-Gonzalez, G.; et al. Diversity and carbon storage across the tropical forest biome. Sci. Rep. 2017, 7, 39102. [Google Scholar] [CrossRef] [Green Version]
- Poorter, L.; van der Sande, M.T.; Thompson, J.; Arets, E.J.M.M.; Alarcón, A.; Álvarez-Sánchez, J.; Ascarrunz, N.; Balvanera, P.; Barajas-Guzmán, G.; Boit, A.; et al. Diversity enhances carbon storage in tropical forests. Glob. Ecol. Biogeogr. 2015, 24, 1314–1328. [Google Scholar] [CrossRef]
- Conti, G.; Díaz, S. Plant functional diversity and carbon storage—An empirical test in semi-arid forest ecosystems. J. Ecol. 2013, 101, 18–28. [Google Scholar] [CrossRef]
- Mandal, R.A.; Dutta, I.C.; Jha, P.K.; Karmacharya, S. Relationship between carbon stock and plant biodiversity in collaborative forests in Terai, Nepal. ISRN Bot. 2013, 2013, 625767. [Google Scholar] [CrossRef] [Green Version]
- Natalia, D.; Arisoesilaningsih, E.; Hairiah, K. Are high carbon stocks in agroforests and forest associated with high plant species diversity? Agrivita 2017, 39, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Kirby, K.R.; Potvin, C. Variation in carbon storage among tree species: Implications for the management of a small-scale carbon sink project. For. Ecol. Manag. 2007, 246, 208–221. [Google Scholar] [CrossRef]
- Murray, J.P.; Grenyer, R.; Wunder, S.; Raes, N.; Jones, J.P. Spatial patterns of carbon, biodiversity, deforestation threat, and REDD+ projects in Indonesia. Conserv. Biol. 2015, 29, 1434–1445. [Google Scholar] [CrossRef] [Green Version]
- Dewi, S.; van Noordwijk, M.; Zulkarnain, M.T.; Dwiputra, A.; Hyman, G.; Prabhu, R.; Gitz, V.; Nasi, R. Tropical forest-transition landscapes: A portfolio for studying people, tree crops and agro-ecological change in context. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2017, 13, 312–329. [Google Scholar] [CrossRef]
- Sanchez, J.L.M.; Cabrales, L.C.; Académica, D.; Biológicas, D.C.; Juárez, U.; Tabasco, A.D. Is there a relationship between floristic diversity and carbon stocks in tropical vegetation in Mexico? Afr. J. Agric. Res. 2012, 7, 2584–2591. [Google Scholar] [CrossRef]
- Kalaba, F.K.; Quinn, C.H.; Dougill, A.J.; Vinya, R. Floristic composition, species diversity and carbon storage in charcoal and agriculture fallows and management implications in Miombo woodlands of Zambia. For. Ecol. Manag. 2013, 304, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Rutishauser, E.; Hérault, B.; Baraloto, C.; Blanc, L.; Descroix, L.; Sotta, E.D.; Ferreira, J.; Kanashiro, M.; Mazzei, L.; D’Oliveira, M.V.N.; et al. Rapid tree carbon stock recovery in managed Amazonian forests. Curr. Biol. 2015, 25, R787–R788. [Google Scholar] [CrossRef] [Green Version]
- Murdiyarso, D.; van Noordwijk, M.; Wasrin, U.R.; Tomich, T.P.; Gillison, A.N. Environmental benefits and sustainable land-use options in the Jambi transect, Sumatra. J. Veg. Sci. 2002, 13, 429–438. [Google Scholar] [CrossRef]
- Martin, P.A.; Newton, A.C.; Bullock, J.M. Carbon pools recover more quickly than plant biodiversity in tropical secondary forests. Proc. Biol. Sci. 2013, 280, 20132236. [Google Scholar] [CrossRef] [PubMed]
- Van Noordwijk, M.; Rahayu, S.; Gebrekirstos, A.; Kindt, R.; Tata, H.L.; Muchugi, A.; Ordonnez, J.C.; Xu, J. Tree diversity as basis of agroforestry. In Sustainable Development through Trees on Farms: Agroforestry in Its Fifth Decade; Van Noordwijk, M., Ed.; World Agroforestry Centre (ICRAF) Southeast Asia Regional Program: Bogor, Indonesia, 2019; pp. 17–44. [Google Scholar]
- Kehlenbeck, K.; Maass, B.L. Crop diversity and classi cation of homegardens in Central Sulawesi, Indonesia. Differences 2004, 63, 53–62. [Google Scholar]
- Dove, M.R. The Banana Tree at the Gate: A History of Marginal Peoples and Global Markets in Borneo; Yale University Press: New Haven, CT, USA, 2011; Volume 3, p. 332. [Google Scholar]
- Van Noordwijk, M.; Agus, F.; Dewi, S.; Purnomo, H. Reducing emissions from land use in Indonesia: Motivation, policy instruments and expected funding streams. Mitig. Adapt. Strateg. Glob. Chang. 2014, 19, 677–692. [Google Scholar] [CrossRef] [Green Version]
- Schroth, G.; Harvey, C.A. Biodiversity conservation in cocoa production landscapes: An overview. Biodivers. Conserv. 2007, 16, 2237–2244. [Google Scholar] [CrossRef]
- Rahmanulloh, A.; Sofiyuddin, M. Profitability of Land-Use Systems in South Sulawesi and Southeast Sulawesi; World Agroforestry Centre: Bogor, Indonesia, 2012; p. 16. [Google Scholar] [CrossRef]
- Dumont, E.S.; Gnahoua, G.M.; Ohouo, L.; Sinclair, F.L.; Vaast, P. Farmers in Cote d’Ivoire value integrating tree diversity in cocoa for the provision of ecosystem services. Agrofor. Syst. 2014, 88, 1047–1066. [Google Scholar] [CrossRef] [Green Version]
- Smiley, G.L.; Kroschel, J. Temporal change in carbon stocks of cocoa-gliricidia agroforests in Central Sulawesi, Indonesia. Agrofor. Syst. 2008, 73, 219–231. [Google Scholar] [CrossRef]
- Triadiati, T.S.; Guhardja, E.; Sudarsono, Q.I.; Leuschner, C. Litterfall production and leaf-litter decomposition at natural forest and cacao agroforestry in Central Sulawesi, Indonesia. Asian J. Biol. Sci. 2011, 4, 2011. [Google Scholar]
- Deheuvels, O.; Rousseau, G.X.; Quiroga, G.S.; Franco, M.D.; Cerda, R.; Mendoza, S.J.V.; Somarriba, E. Biodiversity is affected by changes in management intensity of cocoa-based agroforests. Agrofor. Syst. 2014, 88, 1081–1099. [Google Scholar] [CrossRef]
- Benton, T. Sustainable Intensification; International Plant Nutrition Institute (IPNI): Indonesia, 2016; pp. 95–109. [Google Scholar] [CrossRef]
- Cerda, R.; Deheuvels, O.; Calvache, D.; Niehaus, L.; Saenz, Y.; Kent, J.; Vilchez, S.; Villota, A.; Martinez, C.; Somarriba, E. Contribution of cocoa agroforestry systems to family income and domestic consumption: Looking toward intensification. Agrofor. Syst. 2014, 88, 957–981. [Google Scholar] [CrossRef]
- Monroe, P.H.M.; Gama-Rodrigues, E.F.; Gama-Rodrigues, A.C.; Marques, J.R.B. Soil carbon stocks and origin under different cacao agroforestry systems in southern Bahia, Brazil. Agric. Ecosyst. Environ. 2016, 221, 99–108. [Google Scholar] [CrossRef]
- Jose, S. Agroforestry for ecosystem services and environmental benefits: An overview. Agrofor. Syst. 2009, 76, 1–10. [Google Scholar] [CrossRef]
- Mulyoutami, E.; Roshetko, J.M.; Martini, E.; Awalina, D. Gender roles and knowledge in plant species selection and domestication: A case study in South and Southeast Sulawesi. Int. For. Rev. 2015, 17, 99–111. [Google Scholar] [CrossRef]
- BPS. Konawe in Fugures; Konawe, B.K., Ed.; Statistics of Konawe Regency: Regency, Indonesia, 2015; p. 286. [Google Scholar]
- Hairiah, K.; Sulistyani, H.; Suprayogo, D.; Purnomosidhi, P.; Widodo, R.H.; van Noordwijk, M. Litter layer residence time in forest and coffee agroforestry systems in Sumberjaya, West Lampung. For. Ecol. Manag. 2006, 224, 45–57. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Hairiah, K.; Dewi, S.; Agus, F.; Velarde, S.; Andree, E.; Rahayu, S.; van Noordwijk, M. Measuring Carbon Stocks; World Agroforestry Centre: Bogor, Indonesia, 2011; p. 95. [Google Scholar]
- Eggleston, S.; Buendia, L.; Miwa, K.; Ngara, T.; Tanabe, K. 2006 IPCC Guidelines for National Greenhouse Gas Inventories; Eggleston, S., Buendia, L., Miwa, K., Ngara, T., Tanabe, K., Eds.; IPCC: Geneva, Switzerland, 2006; p. 6. [Google Scholar]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef]
- Yuliasmara, F.; Wibawa, A.; Prawoto, A.A. Carbon stock in different ages and plantation system of cocoa: Allometric approach. Pelita Perkeb. 2009, 25, 86–100. [Google Scholar] [CrossRef]
- Arifin, J. Estimasi Cadangan Karbon pada Berbagai Sistem Penggunaan Lahan di Kecamatan Ngantang, Malang; Brawijaya University: Malang, Indonesia, 2001. [Google Scholar]
- Mokany, K.; Raison, R.J.; Prokushkin, A.S. Critical analysis of root: Shoot ratios in terrestrial biomes. Glob. Chang. Biol. 2006, 12, 84–96. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. An examination of the degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid tritation method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Markum, M.; Soesilaningsih, E.A.; Suprayogo, D.; Hairiah, K. Plant species diversity in relation to carbon stocks at Jangkok watershed, Lombok Island. AGRIVITA 2013, 35, 45–63. [Google Scholar] [CrossRef]
- Faranisa, R. Keanekaragaman Flora Pada Berbagai Tingkat Kepadatan Tanah di Hutan Pegunungan Taman Nasional Bromo Tengger Semeru (TNBTS). Bachelor’s Thesis, Brawijaya University, Malang, Indonesia, 2017. [Google Scholar]
- Kendom, M. Estimasi Emisi Karbon di DAS Casteel Timur Berdasarkan Perubahan Tutupan Hutan Alami di Wilayah Kabupaten Asmat, Papua. Master’s Thesis, Brawijaya University, Malang, Indonesia, 2013. [Google Scholar]
- Hairiah, K.; Suprayogo, D.; Saputra, D.D. Reklamasi Lahan Pertanian Pascaerupsi Gunung Kelud: Efisiensi Serapan N Dalam System Agroforestry Kopi-Kakao; Brawijaya University: Malang, Indonesia, 2017; p. 53. [Google Scholar]
- Kurniawan, S. Bio-Physical Characteristics, Vegetation, Biomass, Carbon Stock, and Carbon Sequestration in the Bangsri Micro Watershed-East Java. In Final Report CCCD Project; Brawijaya University and Research group of Tropical Agroforestry: Malang, Indonesia, 2018; p. 53. [Google Scholar]
- Prayogo, C. Biodiversity and Ecosystem Management in the Bangsri Micro Watershed-East Java. Final Report CCCD Project; Brawijaya University and Research Group of Tropical Agroforestry: Malang, Indonesia, 2018; p. 65. [Google Scholar]
- Dawoe, E.; Asante, W.; Acheampong, E.; Bosu, P. Shade tree diversity and aboveground carbon stocks in Theobroma cacao agroforestry systems: Implications for REDD + implementation in a West African cacao landscape. Carbon Balance Manag. 2016, 11, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Middendorp, R.S.; Vanacker, V.; Lambin, E.F. Impacts of shaded agroforestry management on carbon sequestration, biodiversity and farmers income in cocoa production landscapes. Landsc. Ecol. 2018, 33, 1953–1974. [Google Scholar] [CrossRef]
- Duguma, B.; Gockowski, J.; Bakala, J. Smallholder cacao (Theobroma cacao Linn.) cultivation in agroforestry systems of West and Central Africa: Challenges and opportunities. Agrofor. Syst. 2001, 51, 177–188. [Google Scholar] [CrossRef]
- Somarriba, E.; Cerda, R.; Orozco, L.; Cifuentes, M.; Dávila, H.; Espin, T.; Mavisoy, H.; Ávila, G.; Alvarado, E.; Poveda, V.; et al. Carbon stocks and cocoa yields in agroforestry systems of Central America. Agric. Ecosyst. Environ. 2013, 173, 46–57. [Google Scholar] [CrossRef]
- Sanderman, J.; Hengl, T.; Fiske, G.J. Soil carbon debt of 12,000 years of human land use. Proc. Natl. Acad. Sci. USA 2017, 114, 9575–9580. [Google Scholar] [CrossRef] [Green Version]
- Norgrove, L.; Hauser, S. Carbon stocks in shaded Theobroma cacao farms and adjacent secondary forests of similar age in Cameroon. Trop. Ecol. 2013, 54, 15–22. [Google Scholar]
- Hairiah, K.; van Noordwijk, M.; Sari, R.R.; Saputra, D.D.; Suprayogo, D.; Kurniawan, S.; Prayogo, C.; Gusli, S. Soil carbon stocks in Indonesian (agro) forest transitions: Compaction conceals lower carbon concentrations in standard accounting. Agric. Ecosyst. Environ. 2020, 294, 106879. [Google Scholar] [CrossRef]
- Wartenberg, A.C.; Blaser, W.J.; Gattinger, A.; Roshetko, J.M.; Van Noordwijk, M.; Six, J. Does shade tree diversity increase soil fertility in cocoa plantations? Agric. Ecosyst. Environ. 2017, 248, 190–199. [Google Scholar] [CrossRef]
- Ureta, J.U.; Evangelista, K.P.A.; Habito, C.M.D.; Lasco, R.D. Exploring gender preferences in farming system and tree species selection: Perspectives of JESAM smallholder farmers in southern Philippines. J. Environ. Sci. Manag. 2016, I, 56–73. [Google Scholar]
- Lasco, R.D.; Espaldon, M.L.O.; Habito, C.M.D. Smallholder farmers’ perceptions of climate change and the roles of trees and agroforestry in climate risk adaptation: Evidence from Bohol, Philippines. Agrofor. Syst. 2015, 90, 521–540. [Google Scholar] [CrossRef]
- Mustapha, R.I.; Jimosh, S.O. Farmer’s preferences for tree species on agroforestry system in Ijebu North Local Government Area, Ogun State, Nigeria. J. Agric. For. Soc. Sci. 2012, 10, 176–187. [Google Scholar]
- Nair, P.K.R.; Garrity, D. Agroforestry—The Future of Global Land Use; Springer International Publishing: New York, NY, USA, 2012. [Google Scholar]
- Liang, J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.D.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; et al. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 354, aaf8957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roshetko, J.M.; Snelder, D.J.; Lasco, R.D.; Van Noordwijk, M. Future challenge: A Paradigm shift in the forestry sector. In Smallholder Tree Growing for Rural Development and Environmental Services; Snelder, D.J., Lasco, R.D., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 453–485. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant species | Equation | Source |
|---|---|---|
| Tropical trees (moist forest) | AGBest = WD*exp(−1.499 + 2.148ln(D) + 0.148ln(D) + 0.207(ln(D))2 − 0.0281(ln(D))3) | Chave et al., 2005 |
| Cacao | AGBest = 0.1208 D1.98 | Yuliasmara, 2008 |
| Banana | AGBest = 0.030 D2.13 | Arifin, 2001 |
| Palms | AGBest = 0.118 D2.53 | Brown, 1997 |
| Land Use Systems | Tree Density, Trees ha−1 | Total BA, m2 ha−1 | Number of Species | H | Dominant/Codominant Species | Benefit |
|---|---|---|---|---|---|---|
| DF | 1275a | 18.42a | 28 | 2.36a | Metrosideros petiolata, Homalium foetidum | Timber |
| CAF | 1317a | 9.14b | 18 | 0.93b | Theobroma cacao, Durio zibethinus, Lansium domesticum | Fruits |
| SAF | 1267a | 7.63b | 4 | 0.58b | Theobroma cacao, Gliricidia sepium | Fruits, Fodder |
| CM | 900b | 8.32b | 2 | 0.24c | Theobroma cacao | Fruits |
| Land Use Systems | Tree, Mg ha−1 | Understorey, Mg ha−1 | Woody necromass, Mg ha−1 | Litter, Mg ha−1 |
|---|---|---|---|---|
| DF | 288.41a | 0.64b | 137.54a | 6.98a |
| CAF | 52.64b | 0.27b | 41.53ab | 7.02a |
| SAF | 54.00b | 0.11b | 5.68b | 5.51a |
| CM | 35.25bc | 0.19b | 3.31b | 4.26a |
| CR | 0c | 0.91a | 0b | 0b |
| No | Species | Main Benefits | Frequency (% of Plots) | Farmer’s Preference (Rank) | |
|---|---|---|---|---|---|
| Male | Female | ||||
| 1 | Theobroma cacao (cacao) | Bean | 100 | 1 | 1 |
| 2 | Pogostemon cablin (patchouli) | Oil | 78 | 2 | 2 |
| 3 | Gliricidia sepium (mother of cacao) | Fodder (leave), climbed tree for pepper | 78 | 11 | 9 |
| 4 | Musa sp (banana) | Fruits, vegetable (flower), cultural services (leaf), toys (trunk) | 67 | - | 4 |
| 5 | Cocos nucifera (coconut) | Fruits, cultural services (leaf), roof (leaf), toys (trunk) | 56 | - | 5 |
| 6 | Capsicum annum (chili pepper) | Vegetable/spice | 56 | - | 7 |
| 7 | Piper nigrum (pepper) | Vegetable/spice | 44 | 10 | 6 |
| 8 | Fagraea fragrans (tembesu) | Timber | 44 | 3 | 10 |
| 9 | Tectona grandis (teak) | Timber | 44 | 4 | 11 |
| 10 | Anthocephalus cadamba (jabon) | Timber | 44 | 5 | - |
| 11 | Durio zibethinus (durian) | Fruits | 44 | 6 | 3 |
| 12 | Lansium domesticum (langsat) | Fruits | 22 | 7 | - |
| 13 | Mangifera indica (mango) | Fruits | 22 | 8 | - |
| 14 | Nephelium lappaceum (rambutan) | Fruits | 22 | 9 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sari, R.R.; Saputra, D.D.; Hairiah, K.; Rozendaal, D.M.A.; Roshetko, J.M.; van Noordwijk, M. Gendered Species Preferences Link Tree Diversity and Carbon Stocks in Cacao Agroforest in Southeast Sulawesi, Indonesia. Land 2020, 9, 108. https://doi.org/10.3390/land9040108
Sari RR, Saputra DD, Hairiah K, Rozendaal DMA, Roshetko JM, van Noordwijk M. Gendered Species Preferences Link Tree Diversity and Carbon Stocks in Cacao Agroforest in Southeast Sulawesi, Indonesia. Land. 2020; 9(4):108. https://doi.org/10.3390/land9040108
Chicago/Turabian StyleSari, Rika Ratna, Danny Dwi Saputra, Kurniatun Hairiah, Danaë M. A. Rozendaal, James M. Roshetko, and Meine van Noordwijk. 2020. "Gendered Species Preferences Link Tree Diversity and Carbon Stocks in Cacao Agroforest in Southeast Sulawesi, Indonesia" Land 9, no. 4: 108. https://doi.org/10.3390/land9040108