
How to Improve the Benefits of Short-Term Fallow on Soil Physicochemical and Microbial Properties: A Case Study from the Yellow River Delta
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Experimental Design
- CK, traditional tillage (deep ploughing), planting sorghum, and applying nitrogen, phosphorus, and potassium fertilizer (N:P:K 3:1:3) and urea (46.2% of total nitrogen content), a total of 330 kg ha−1.
- F-noman, no fertilizer input, no tillage (artificial weeding without tillage), and spontaneous vegetation fallow, later referred to as natural fallow.
- M-noman, no fertilizer input, only minimal tillage (artificial weeding), mixed sowing of corn and alfalfa, referred to as grain and grass fallow.
- F-irrig, the vegetation is naturally growing vegetation, no fertilizer input, no tillage, irrigation, irrigation to about 60% soil water content.
- M-irrig, the vegetation was mixed with corn and alfalfa, and there was no fertilizer input. Only minimal tillage (artificial weeding) was carried out, and the soil water content was about 60%.
- F-ferti, no tillage, natural fallow combined with the application of decomposed cow manure fertilizer (organic carbon content 400 g kg−1, total nitrogen 7.0 g kg−1, total phosphorus content 11.5 g kg−1, and total potassium content 9.8 g kg−1), a total amount of 1500 kg ha−1.
- M-ferti, minimum tillage, grain and grass fallow (vegetation type corn and alfalfa) combined with decomposed cow manure fertilizer, a total amount of 1500 kg ha−1.
2.2. Soil Sampling
2.3. Determination of Soil Physicochemical Properties and Microbial Biomass
2.4. 16SrDNA Sequence Analysis of Soil Microbial Communities
2.5. Statistical Analysis
3. Results
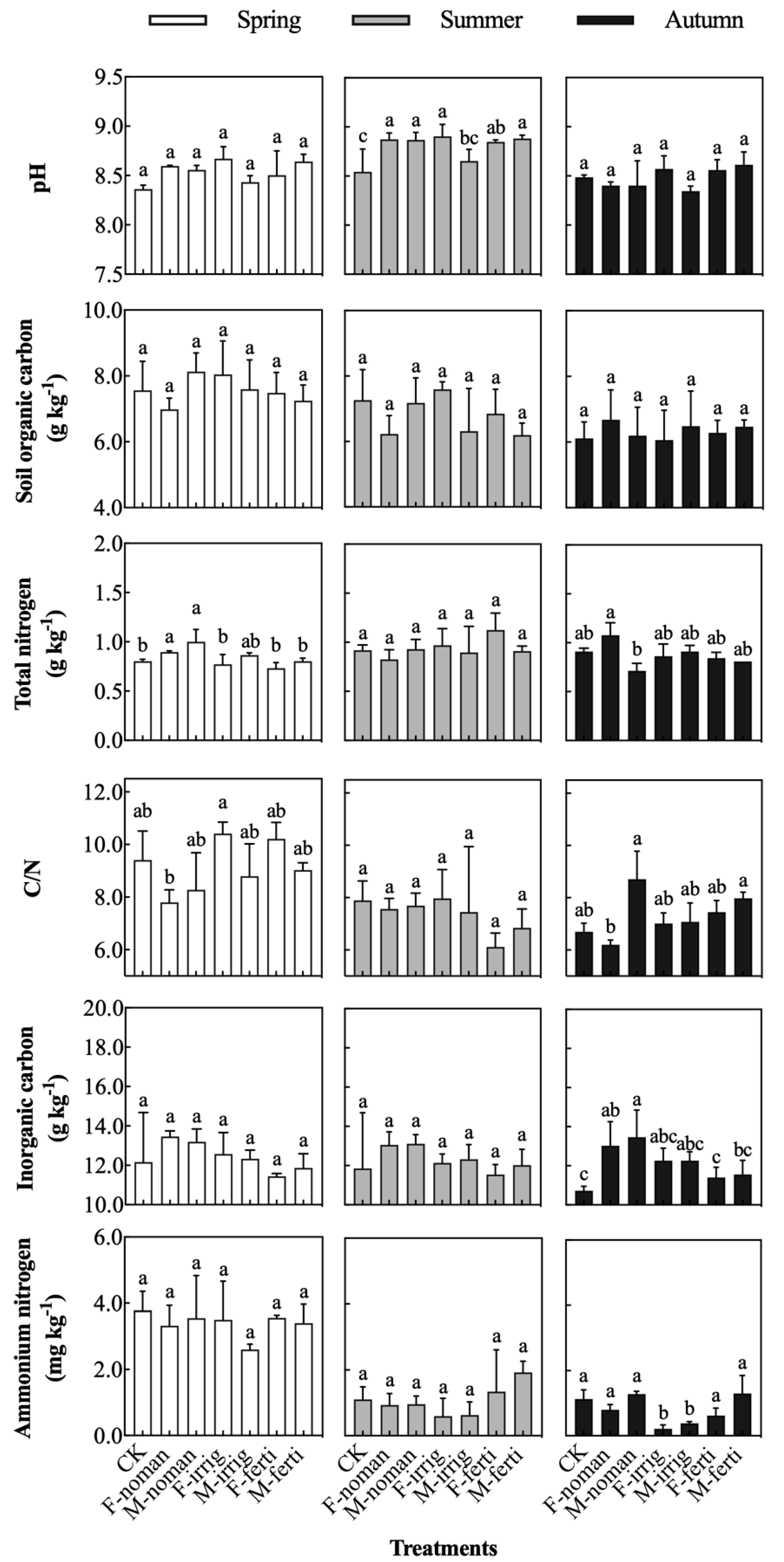
3.1. Effects of Seasons and Management Methods on Soil Physicochemical Properties
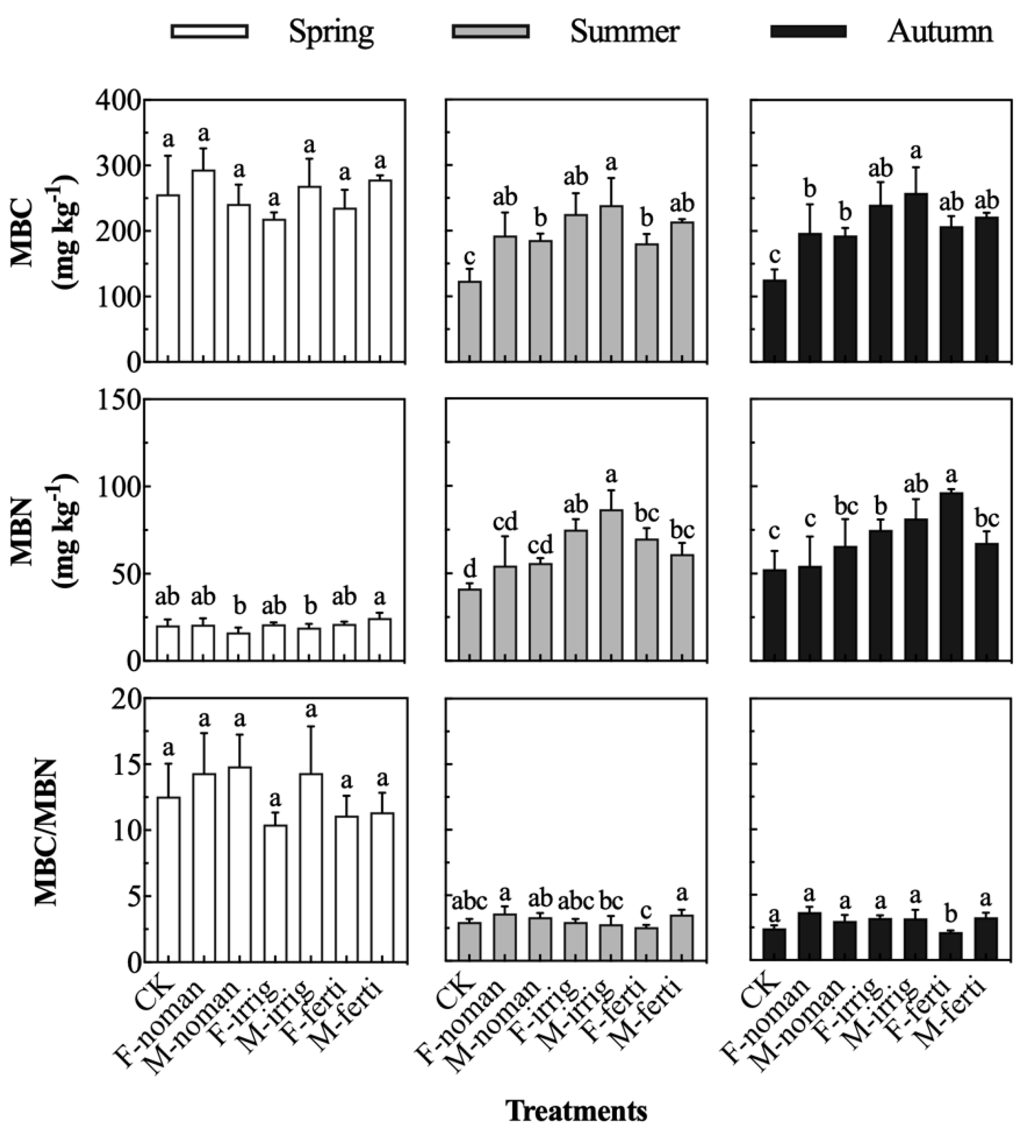
3.2. Effects of Seasons and Management Methods on Soil Microbial Biomass
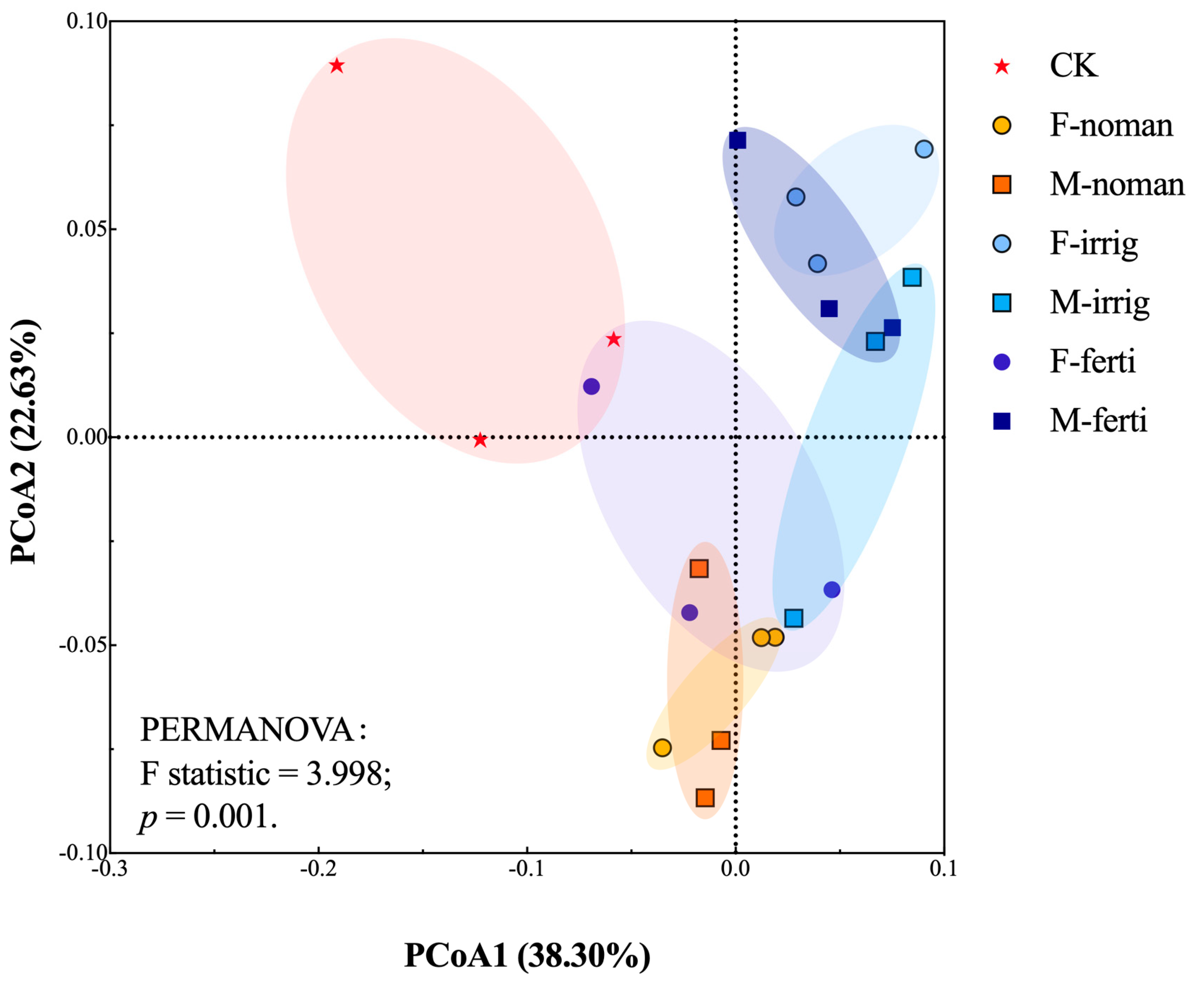
3.3. Effects of Seasons and Different Treatments on Soil Microbial Communities
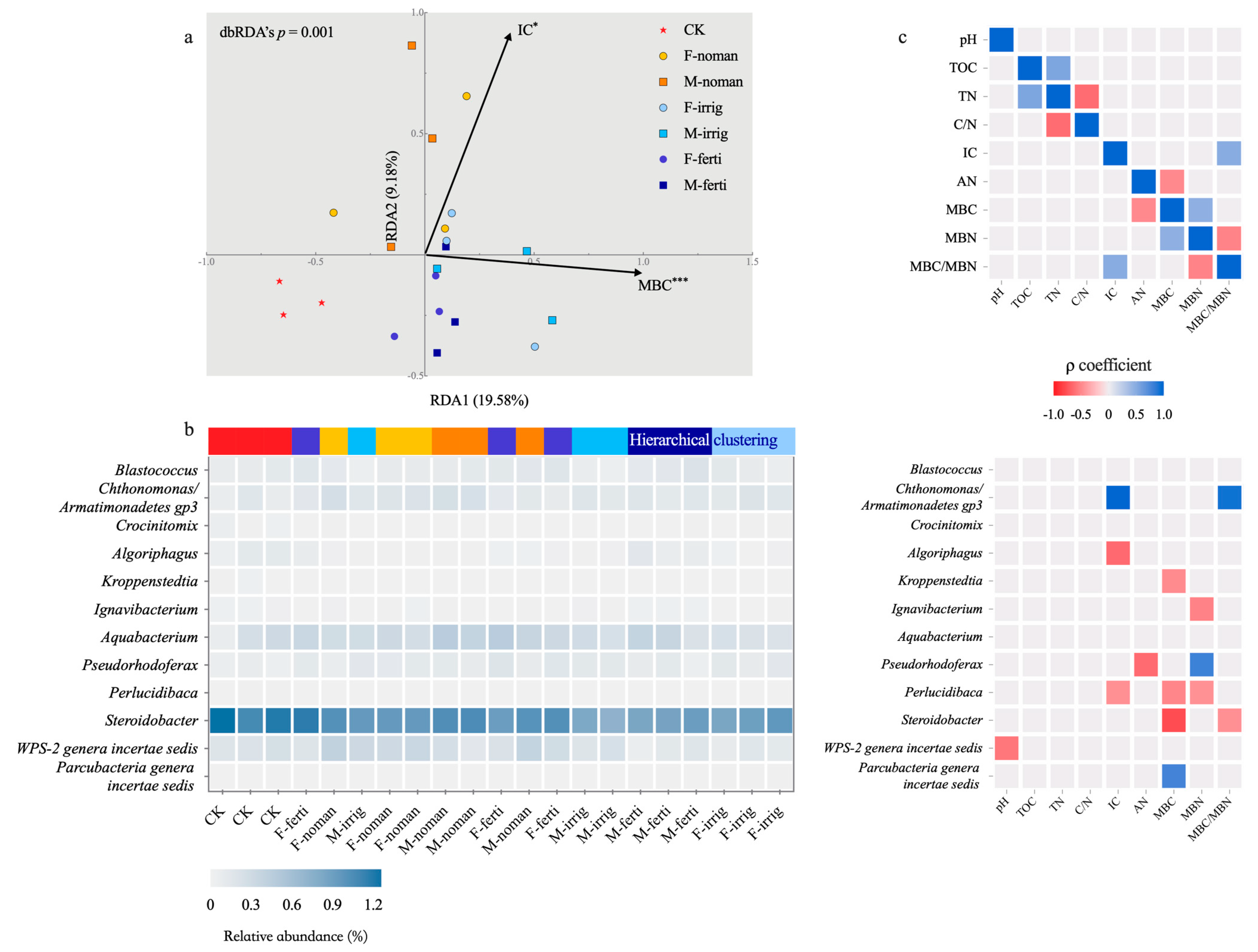
3.4. Relationship between Soil Properties and Microbial Communities in Autumn
4. Discussion
4.1. Soil Carbon and Nitrogen Content Were Affected by Fallowing Management Methods
4.2. Microbial Properties Changes Caused by Fallow Management
4.3. Implications for Fallow Management
- Unmanaged native vegetation fallow (F-noman) improved the inorganic carbon storage, soil nitrification, and disease resistance, and adjusted the soil pH to inhibit denitrification, nitrate reduction, and organic decomposition.
- Unmanaged grain and grass fallow (M-noman) also improved the inorganic carbon content, soil nitrification, and disease resistance, and enhanced the decomposition ability of organic pollutants, inhibiting nitrate reduction and organic decomposition. However, this management method resulted in a reduction in the total nitrogen content and an increase in the carbon–nitrogen ratio to a certain extent.
- Native vegetation fallow under irrigation management (F-irrig) reduced the content of ammonium nitrogen, inhibited nitrate reduction, denitrification, and organic decomposition, increased the content of microbial nitrogen, and promoted the growth of surface vegetation.
- Crop and pasture vegetation fallow under irrigation management (M-irrig) had the same effect as F-irrig, which reduced the content of ammonium nitrogen, inhibited nitrate reduction, denitrification, and organic decomposition, increased the content of microbial nitrogen, and promoted the growth of surface vegetation.
- Native vegetation fallow under the management of farm manure (F-ferti) improved the microbial biomass nitrogen, the ability to resist soil-borne diseases, the decomposition of organic pollutants, and the ability to promote vegetation growth. In terms of nitrate reduction, the effects of different bacteria were cancelled out, so the effect was not considered.
- Crop and pasture vegetation fallow managed using farmyard manure (M-ferti) only improved the soil’s ability to resist soil-borne diseases and inhibited denitrification, but the other effects were not significant.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chen, R.; de Sherbinin, A.; Ye, C.; Shi, G. China’s Soil Pollution: Farms on the Frontline. Science 2014, 344, 691. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Pijanowski, B.C. The effects of China’s cultivated land balance program on potential land productivity at a national scale. Appl. Geogr. 2014, 46, 158–170. [Google Scholar] [CrossRef]
- He, X.; Xu, Y.; Zhang, X. Traditional farming system for soil conservation on slope farmland in southwestern China. Soil Tillage Res. 2007, 94, 193–200. [Google Scholar] [CrossRef]
- Bai, Z.; Caspari, T.; Gonzalez, M.R.; Batjes, N.H.; Mäder, P.; Bünemann, E.K.; de Goede, R.; Brussaard, L.; Xu, M.; Ferreira, C.S.S.; et al. Effects of agricultural management practices on soil quality: A review of long-term experiments for Europe and China. Agric. Ecosyst. Environ. 2018, 265, 1–7. [Google Scholar] [CrossRef]
- Feng, Z.; Yang, Y.; Zhang, Y.; Zhang, P.; Li, Y. Grain-for-green policy and its impacts on grain supply in West China. Land Use Policy 2005, 22, 301–312. [Google Scholar] [CrossRef]
- Wang, Q.; Lu, C.; Li, H.; He, J.; Sarker, K.K.; Rasaily, R.G.; Liang, Z.; Qiao, X.; Li, H.; McHugh, A.D.J. The effects of no-tillage with subsoiling on soil properties and maize yield: 12-Year experiment on alkaline soils of Northeast China. Soil Tillage Res. 2014, 137, 43–49. [Google Scholar] [CrossRef]
- Li, G.; He, T.; Zhang, M.; Wu, C. Spatiotemporal variations in the eco-health condition of China’s long-term stable cultivated land using Google Earth Engine from 2001 to 2019. Appl. Geogr. 2022, 149, 102819. [Google Scholar] [CrossRef]
- Derry, D.D.; Voroney, R.P.; Briceño, J.A. Long-term effects of short-fallow frijol tapado on soil phosphorus pools in Costa Rica. Agric. Ecosyst. Environ. 2005, 110, 91–103. [Google Scholar] [CrossRef]
- Guo, L.-J.; Zhang, Z.-S.; Wang, D.-D.; Li, C.-F.; Cao, C.-G. Effects of short-term conservation management practices on soil organic carbon fractions and microbial community composition under a rice-wheat rotation system. Biol. Fertil. Soils 2015, 51, 65–75. [Google Scholar] [CrossRef]
- Catão, E.C.P.; Lopes, F.A.C.; Rubini, M.R.; Nardoto, G.B.; Prosser, J.I.; Krüger, R.H. Short-term impact of soybean management on ammonia oxidizers in a Brazilian savanna under restoration as revealed by coupling different techniques. Biol. Fertil. Soils 2016, 52, 401–412. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Wu, C. Effects of Short-Term Set-Aside Management Practices on Soil Microorganism and Enzyme Activity in China. Int. J. Environ. Res. Public Health 2017, 14, 913. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; He, T.; Zhang, M.; Wu, C. Could Land Abandonment with Human Intervention Benefit Cropland Restoration? From the Perspective of Soil Microbiota. Land 2021, 10, 1049. [Google Scholar] [CrossRef]
- Li, G.; de Vries, W.T.; Wu, C.; Zheng, H. Improvement of subsoil physicochemical and microbial properties by short-term fallow practices. PeerJ 2019, 7, e7501. [Google Scholar] [CrossRef]
- Li, G.; Zhang, M.; Wu, C. Short-term fallow practices drive soil bacterial community changes: A case study from China. Appl. Soil Ecol. 2021, 165, 103988. [Google Scholar] [CrossRef]
- Zak, D.R.; Holmes, W.E.; White, D.C.; Peacock, A.D.; Tilman, D. PLANT DIVERSITY, SOIL MICROBIAL COMMUNITIES, AND ECOSYSTEM FUNCTION: ARE THERE ANY LINKS? Ecology 2003, 84, 2042–2050. [Google Scholar] [CrossRef] [Green Version]
- Rukun, L. General status of nutrients (N, P, K) in soils of China. Acta Pedol. Sin. 1989, 26, 280–286. [Google Scholar]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Joergensen, R.G. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the kEC value. Soil Biol. Biochem. 1996, 28, 25–31. [Google Scholar] [CrossRef]
- Angenent, L.T.; Kelley, S.T.; Amand, A.S.; Pace, N.R.; Hernandez, M.T. Molecular identification of potential pathogens in water and air of a hospital therapy pool. Proc. Natl. Acad. Sci. USA 2005, 102, 4860. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapteva, Y.S.; Zolova, O.E.; Shlyapnikov, M.G.; Tsfasman, I.M.; Muranova, T.A.; Stepnaya, O.A. Cloning and expression analysis of genes encoding lytic endopeptidases L1 and L5 from Lysobacter sp. strain XL1. Appl. Environ. Microbiol. 2012, 78, 7082–7089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Wu, C.; Gao, W. Effects of short-term fallow managements on soil microbial properties: A case study in China. Appl. Soil Ecol. 2018, 125, 128–137. [Google Scholar] [CrossRef]
- An, H.; Wu, X.; Zhang, Y.; Tang, Z. Effects of land-use change on soil inorganic carbon: A meta-analysis. Geoderma 2019, 353, 273–282. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, X.; Li, X.; Wang, J.; Xu, M.; Li, D. Dynamics of soil organic and inorganic carbon in the cropland of upper Yellow River Delta, China. Sci. Rep. 2016, 6, 36105. [Google Scholar] [CrossRef]
- Homyak, P.M.; Allison, S.D.; Huxman, T.E.; Goulden, M.L.; Treseder, K.K. Effects of Drought Manipulation on Soil Nitrogen Cycling: A Meta-Analysis. J. Geophys. Res. Biogeosci. 2017, 122, 3260–3272. [Google Scholar] [CrossRef] [Green Version]
- Jia, G.-M.; Cao, J.; Wang, G. Influence of land management on soil nutrients and microbial biomass in the central loess plateau, northwest China. Land Degrad. Dev. 2005, 16, 455–462. [Google Scholar] [CrossRef]
- Kumar, B.; Dhar, S.; Paul, S.; Paramesh, V.; Dass, A.; Upadhyay, P.K.; Kumar, A.; Abdelmohsen, S.A.M.; Alkallas, F.H.; El-Abedin, T.K.Z.; et al. Microbial Biomass Carbon, Activity of Soil Enzymes, Nutrient Availability, Root Growth, and Total Biomass Production in Wheat Cultivars under Variable Irrigation and Nutrient Management. Agronomy 2021, 11, 669. [Google Scholar] [CrossRef]
- Zuo, X.; Zhang, J.; Lv, P.; Zhou, X.; Li, Y.; Luo, Y.; Luo, Y.; Lian, J.; Yue, X. Plant functional diversity mediates the effects of vegetation and soil properties on community-level plant nitrogen use in the restoration of semiarid sandy grassland. Ecol. Indic. 2016, 64, 272–280. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, H.; Sun, L.; Qi, G.; Chen, S.; Zhao, X. Microbial community composition is related to soil biological and chemical properties and bacterial wilt outbreak. Sci. Rep. 2017, 7, 343. [Google Scholar] [CrossRef] [Green Version]
- Swenson, T.L.; Karaoz, U.; Swenson, J.M.; Bowen, B.P.; Northen, T.R. Linking soil biology and chemistry in biological soil crust using isolate exometabolomics. Nat. Commun. 2018, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.C.-Y.; Dunfield, P.F.; Morgan, X.C.; Crowe, M.A.; Houghton, K.M.; Vyssotski, M.; Ryan, J.L.J.; Lagutin, K.; McDonald, I.R.; Stott, M.B. Chthonomonas calidirosea gen. nov., sp. nov., an aerobic, pigmented, thermophilic microorganism of a novel bacterial class, Chthonomonadetes classis nov., of the newly described phylum Armatimonadetes originally designated candidate division OP10. Int. J. Syst. Evol. Microbiol. 2011, 61, 2482–2490. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.; Ruan, Y.; Xue, C.; Zhong, S.; Li, R.; Shen, Q. Soils naturally suppressive to banana Fusarium wilt disease harbor unique bacterial communities. Plant Soil 2015, 393, 21–33. [Google Scholar] [CrossRef]
- Nedashkovskaya, O.I.; Vancanneyt, M.; Van Trappen, S.; Vandemeulebroecke, K.; Lysenko, A.M.; Rohde, M.; Falsen, E.; Frolova, G.M.; Mikhailov, V.V.; Swings, J. Description of Algoriphagus aquimarinus sp. nov., Algoriphagus chordae sp. nov. and Algoriphagus winogradskyi sp. nov., from sea water and algae, transfer of Hongiella halophila Yi and Chun 2004 to the genus Algoriphagus as Algoriphagus halophilus comb. nov. and emended descriptions of the genera Algoriphagus Bowman et al. 2003 and Hongiella Yi and Chun 2004. Int. J. Syst. Evol. Microbiol. 2004, 54, 1757–1764. [Google Scholar] [CrossRef]
- Xiong, W.; Mathies, C.; Bradshaw, K.; Carlson, T.; Tang, K.; Wang, Y. Benzene removal by a novel modification of enhanced anaerobic biostimulation. Water Res. 2012, 46, 4721–4731. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Liu, Y.; Shi, D. Effect of lead pollution on bacterial diversity in rhizosphere of clover based on high-throughput sequencing. Res. Environ. Sci. 2018, 31, 102–110. [Google Scholar]
- Aburto-Medina, A.; Adetutu, E.M.; Aleer, S.; Weber, J.; Patil, S.S.; Sheppard, P.J.; Ball, A.S.; Juhasz, A.L. Comparison of indigenous and exogenous microbial populations during slurry phase biodegradation of long-term hydrocarbon-contaminated soil. Biodegradation 2012, 23, 813–822. [Google Scholar] [CrossRef]
- Bruland, N.; Bathe, S.; Willems, A.; Steinbüchel, A. Pseudorhodoferax soli gen. nov., sp. nov. and Pseudorhodoferax caeni sp. nov., two members of the class Betaproteobacteria belonging to the family Comamonadaceae. Int. J. Syst. Evol. Microbiol. 2009, 59, 2702–2707. [Google Scholar] [CrossRef] [Green Version]
- Jordaan, K.; Bezuidenhout, C.C. Bacterial community composition of an urban river in the North West Province, South Africa, in relation to physico-chemical water quality. Environ. Sci. Pollut. Res. 2016, 23, 5868–5880. [Google Scholar] [CrossRef] [PubMed]
- Fahrbach, M.; Kuever, J.; Remesch, M.; Huber, B.E.; Kämpfer, P.; Dott, W.; Hollender, J. Steroidobacter denitrificans gen. nov., sp. nov., a steroidal hormone-degrading gammaproteobacterium. Int. J. Syst. Evol. Microbiol. 2008, 58, 2215–2223. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Physicochemical Properties | Sample Times’ Effects | Treatments’ Effects | The Interactions |
|---|---|---|---|
| pH | 53.011 *** | 4.588 ** | 1.614 |
| SOC (g kg−1) | 24.190 *** | 0.520 | 1.577 |
| Total N (g kg−1) | 3.759 | 0.534 | 5.311 *** |
| C/N | 73.854 *** | 1.424 | 3.983 *** |
| Inorganic C (g kg−1) | 0.994 | 2.129 | 0.485 |
| Ammonium N (mg kg−1) | 145.253 *** | 2.559 | 0.943 |
| Microbial Properties | Sample Times’ Effects | Treatments’ Effects | Interactions |
|---|---|---|---|
| Microbial biomass C | 185.161 *** | 4.302 * | 6.365 *** |
| Microbial biomass N | 346.459 *** | 7.535 *** | 6.554 *** |
| Microbial biomass C/N | 198.435 *** | 2.077 | 1.950 |
| Factors | F-Statistics | p Values |
|---|---|---|
| Sample times effects | 12.782 | 0.001 |
| Treatments effects | 0.731 | 0.575 |
| Interactions | 12.143 | 0.001 |
| Spring | ||
| Treatments effects | 0.824 | 0.553 |
| Summer | ||
| Treatments effects | 1.185 | 0.272 |
| Autumn | ||
| Treatments effects | 1.196 | 0.245 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.; Wang, Y.; Lv, X.; Yue, S.; Liu, H.; Li, G.; Zhao, J. How to Improve the Benefits of Short-Term Fallow on Soil Physicochemical and Microbial Properties: A Case Study from the Yellow River Delta. Land 2023, 12, 1426. https://doi.org/10.3390/land12071426
Lin Y, Wang Y, Lv X, Yue S, Liu H, Li G, Zhao J. How to Improve the Benefits of Short-Term Fallow on Soil Physicochemical and Microbial Properties: A Case Study from the Yellow River Delta. Land. 2023; 12(7):1426. https://doi.org/10.3390/land12071426
Chicago/Turabian StyleLin, Yaoben, Yuanbo Wang, Xingjun Lv, Shuangyan Yue, Hongmei Liu, Guangyu Li, and Jinghui Zhao. 2023. "How to Improve the Benefits of Short-Term Fallow on Soil Physicochemical and Microbial Properties: A Case Study from the Yellow River Delta" Land 12, no. 7: 1426. https://doi.org/10.3390/land12071426