

Using and Creating Microclimates for Cork Oak Adaptation to Climate Change
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
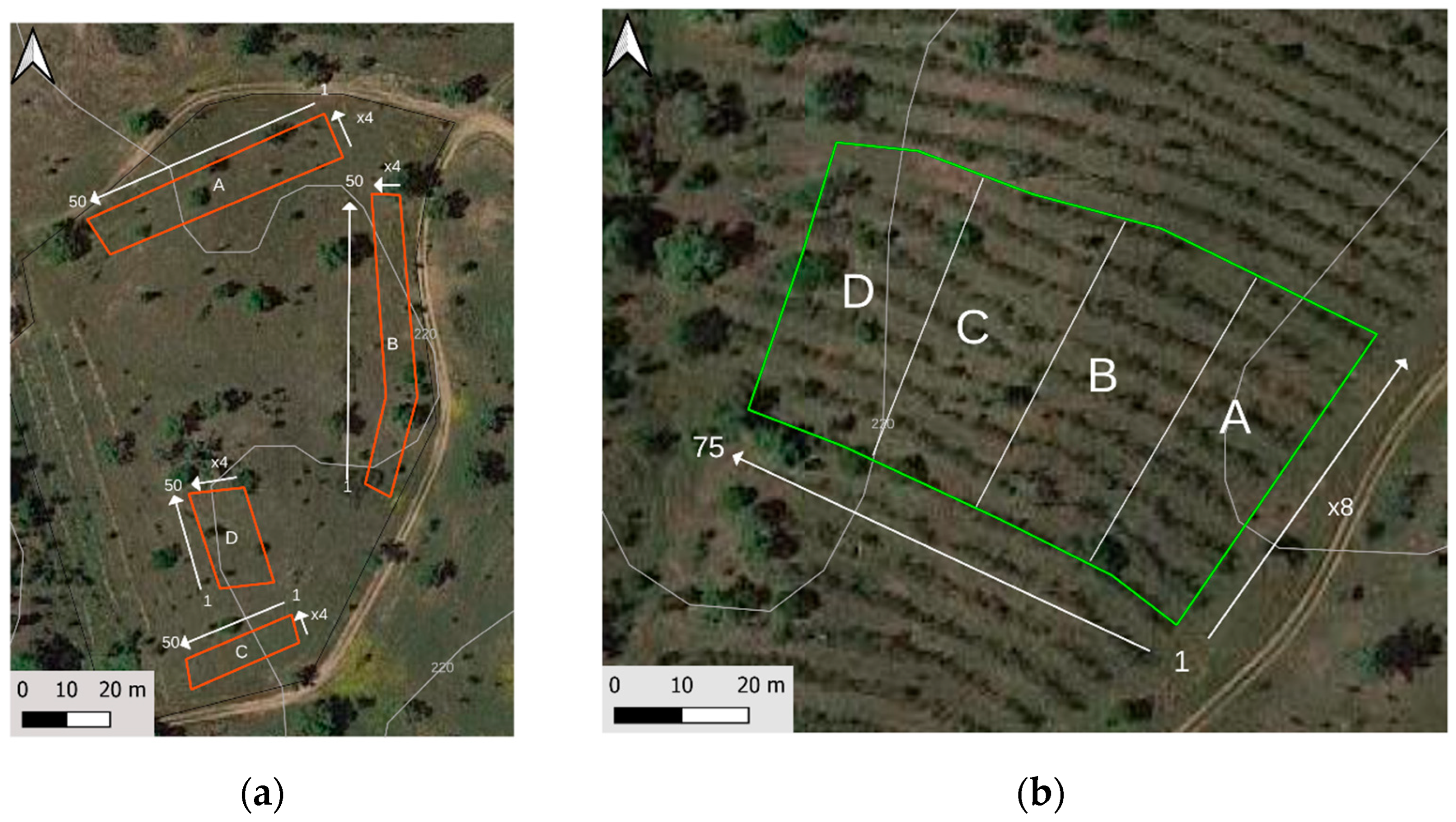
2.2. Experimental Design
2.3. Microclimatic Characterization—Complementary Data
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodrigo Comino, J. Desertification and Degradation Risks vs. Poverty: A Key Topic in Mediterranean Europe. Cuad. Investig. Geográfica 2022, 48, 23–40. [Google Scholar] [CrossRef]
- del Pozo, A.; Brunel-Saldias, N.; Engler, A.; Ortega-Farias, S.; Acevedo-Opazo, C.; Lobos, G.A.; Jara-Rojas, R.; Molina-Montenegro, M.A. Climate Change Impacts and Adaptation Strategies of Agriculture in Mediterranean-Climate Regions (MCRs). Sustainability 2019, 11, 2769. [Google Scholar] [CrossRef] [Green Version]
- Ruti, P.M.; Somot, S.; Giorgi, F.; Dubois, C.; Flaounas, E.; Obermann, A.; Dell’Aquila, A.; Pisacane, G.; Harzallah, A.; Lombardi, E.; et al. Med-CORDEX Initiative for Mediterranean Climate Studies. Bull. Am. Meteorol. Soc. 2015, 97, 1187–1208. [Google Scholar] [CrossRef] [Green Version]
- Barredo, J.I.; Caudullo, G.; Dosio, A. Mediterranean Habitat Loss under Future Climate Conditions: Assessing Impacts on the Natura 2000 Protected Area Network. Appl. Geogr. 2016, 75, 83–92. [Google Scholar] [CrossRef]
- Halbac-Cotoara-Zamfir, R.; Smiraglia, D.; Quaranta, G.; Salvia, R.; Salvati, L.; Giménez-Morera, A. Land Degradation and Mitigation Policies in the Mediterranean Region: A Brief Commentary. Sustainability 2020, 12, 8313. [Google Scholar] [CrossRef]
- Wiebe, K.; Lotze-Campen, H.; Sands, R.; Tabeau, A.; van der Mensbrugghe, D.; Biewald, A.; Bodirsky, B.; Islam, S.; Kavallari, A.; Mason-D’Croz, D.; et al. Climate Change Impacts on Agriculture in 2050 under a Range of Plausible Socioeconomic and Emissions Scenarios. Environ. Res. Lett. 2015, 10, 085010. [Google Scholar] [CrossRef]
- Yang, C.; Fraga, H.; van Ieperen, W.; Trindade, H.; Santos, J.A. Effects of Climate Change and Adaptation Options on Winter Wheat Yield under Rainfed Mediterranean Conditions in Southern Portugal. Clim. Chang. 2019, 154, 159–178. [Google Scholar] [CrossRef] [Green Version]
- Pinto-Correia, T.; Ribeiro, N.; Sá-Sousa, P. Introducing the Montado, the Cork and Holm Oak Agroforestry System of Southern Portugal. Agrofor. Syst. 2011, 82, 99. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, T.M.M.; Coelho, C.O.A.; Ferreira, A.J.D.; Charlton, C.A. Land Degradation Processes in Portugal: Farmers’ Perceptions of the Application of European Agroforestry Programmes. Land Degrad. Dev. 2002, 13, 177–188. [Google Scholar] [CrossRef]
- Fragoso, R.; Marques, C.; Lucas, M.R.; Martins, M.B.; Jorge, R. The Economic Effects of Common Agricultural Policy on Mediterranean Montado/Dehesa Ecosystem. J. Policy Model. 2011, 33, 311–327. [Google Scholar] [CrossRef]
- Correia, T.P. Threatened Landscape in Alentejo, Portugal: The ‘Montado’ and Other ‘Agro-Silvo-Pastoral’ Systems. Landsc. Urban Plan. 1993, 24, 43–48. [Google Scholar] [CrossRef]
- Muñoz-Rojas, J.; Pinto-Correia, T.; Thorsoe, M.H.; Noe, E. The Portuguese Montado: A Complex System under Tension between Different Land Use Management Paradigms. In Silvicultures-Management and Conservation; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Duque-Lazo, J.; Navarro-Cerrillo, R.M.; Ruíz-Gómez, F.J. Assessment of the Future Stability of Cork Oak (Quercus suber L.) Afforestation under Climate Change Scenarios in Southwest Spain. For. Ecol. Manag. 2018, 409, 444–456. [Google Scholar] [CrossRef]
- Paulo, J.A.; Palma, J.H.N.; Gomes, A.A.; Faias, S.P.; Tomé, J.; Tomé, M. Predicting Site Index from Climate and Soil Variables for Cork Oak (Quercus suber L.) Stands in Portugal. New For. 2015, 46, 293–307. [Google Scholar] [CrossRef] [Green Version]
- Vanhove, M.; Pina-Martins, F.; Coelho, A.C.; Branquinho, C.; Costa, A.; Batista, D.; Príncipe, A.; Sousa, P.; Henriques, A.; Marques, I.; et al. Using Gradient Forest to Predict Climate Response and Adaptation in Cork Oak. J. Evol. Biol. 2021, 34, 910–923. [Google Scholar] [CrossRef]
- Hernández-Morcillo, M.; Burgess, P.; Mirck, J.; Pantera, A.; Plieninger, T. Scanning Agroforestry-Based Solutions for Climate Change Mitigation and Adaptation in Europe. Environ. Sci. Policy 2018, 80, 44–52. [Google Scholar] [CrossRef] [Green Version]
- Regato, P.; IUCN Centre for Mediterranean Cooperation; Food and Agriculture Organization of the United Nations. Adapting to Global Change: Mediterranean Forests; IUCN Centre for Mediterranean Cooperation: Malaga, Spain, 2008; ISBN 978-2-8317-1098-3. [Google Scholar]
- Vilà-Cabrera, A.; Coll, L.; Martínez-Vilalta, J.; Retana, J. Forest Management for Adaptation to Climate Change in the Mediterranean Basin: A Synthesis of Evidence. For. Ecol. Manag. 2018, 407, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Vizinho, A.; Avelar, D.; Fonseca, A.L.; Carvalho, S.; Sucena-Paiva, L.; Pinho, P.; Nunes, A.; Branquinho, C.; Vasconcelos, A.C.; Santos, F.D.; et al. Framing the Application of Adaptation Pathways for Agroforestry in Mediterranean Drylands. Land Use Policy 2021, 104, 105348. [Google Scholar] [CrossRef]
- Aranda, I.; Castro, L.; Alía, R.; Pardos, J.A.; Gil, L. Low Temperature during Winter Elicits Differential Responses among Populations of the Mediterranean Evergreen Cork Oak (Quercus suber). Tree Physiol. 2005, 25, 1085–1090. [Google Scholar] [CrossRef] [Green Version]
- Durrant, T.H.; De Rigo, D.; Caudullo, G. Quercus Suber in Europe: Distribution, Habitat, Usage and Threats; European Atlas of Forest Tree Species; Publications Office of the EU: Luxembourg, 2016; pp. 164–165.
- Aronson, J.; Pereira, J.S.; Pausas, J.G. Cork Oak Woodlands on the Edge: Ecology, Adaptive Management, and Restoration; Island Press: Washington, DC, USA, 2012. [Google Scholar]
- Ferreira, A.G.; Gonçalves, A.C.; Pinheiro, A.C.; Pinto Gomes, C.; Ilhéu, M.; Neves, N.; Ribeiro, N.; Santos, P. Plano Específico de Ordenamento Florestal Para o Alentejo; Universidade de Évora: Évora, Portugal, 2001. [Google Scholar]
- Ghouil, H.; Montpied, P.; Epron, D.; Ksontini, M.; Hanchi, B.; Dreyer, E. Thermal Optima of Photosynthetic Functions and Thermostability of Photochemistry in Cork Oak Seedlings. Tree Physiol. 2003, 23, 1031–1039. [Google Scholar] [CrossRef] [Green Version]
- Aubard, V.; Paulo, J.A.; Silva, J.M.N. Long-Term Monitoring of Cork and Holm Oak Stands Productivity in Portugal with Landsat Imagery. Remote Sens. 2019, 11, 525. [Google Scholar] [CrossRef] [Green Version]
- Lecomte, X.; Paulo, J.A.; Tomé, M.; Veloso, S.; Firmino, P.N.; Faias, S.P.; Caldeira, M.C. Shrub Understorey Clearing and Drought Affects Water Status and Growth of Juvenile Quercus suber Trees. For. Ecol. Manag. 2022, 503, 119760. [Google Scholar] [CrossRef]
- Leite, C.; Oliveira, V.; Lauw, A.; Pereira, H. Effect of a Drought on Cork Growth Along the Production Cycle. In Theory and Practice of Climate Adaptation; Alves, F., Leal Filho, W., Azeiteiro, U., Eds.; Climate Change Management; Springer International Publishing: Cham, Switzerland, 2018; pp. 127–136. ISBN 978-3-319-72874-2. [Google Scholar]
- Catalão, J.; Navarro, A.; Calvão, J. Mapping Cork Oak Mortality Using Multitemporal High-Resolution Satellite Imagery. Remote Sensing 2022, 14, 2750. [Google Scholar] [CrossRef]
- Pérez-Devesa, M.; Cortina, J.; Vilagrosa, A.; Vallejo, R. Shrubland Management to Promote Quercus suber L. Establishment. For. Ecol. Manag. 2008, 255, 374–382. [Google Scholar] [CrossRef]
- Pulido, F.J.; Díaz, M. Regeneration of a Mediterranean Oak: A Whole-Cycle Approach. Écoscience 2005, 12, 92–102. [Google Scholar] [CrossRef]
- Plieninger, T.; Rolo, V.; Moreno, G. Large-Scale Patterns of Quercus ilex, Quercus suber, and Quercus pyrenaica Regeneration in Central-Western Spain. Ecosystems 2010, 13, 644–660. [Google Scholar] [CrossRef]
- Trubat, R.; Cortina, J.; Vilagrosa, A. Nursery Fertilization Affects Seedling Traits but Not Field Performance in Quercus suber L. J. Arid Environ. 2010, 74, 491–497. [Google Scholar] [CrossRef]
- Vizinho, A.; Avelar, D.; Branquinho, C.; Capela Lourenço, T.; Carvalho, S.; Nunes, A.; Sucena-Paiva, L.; Oliveira, H.; Fonseca, A.L.; Duarte Santos, F.; et al. Framework for Climate Change Adaptation of Agriculture and Forestry in Mediterranean Climate Regions. Land 2021, 10, 161. [Google Scholar] [CrossRef]
- Príncipe, A.; Nunes, A.; Pinho, P.; Aleixo, C.; Neves, N.; Branquinho, C. Local-Scale Factors Matter for Tree Cover Modelling in Mediterranean Drylands. Sci. Total Environ. 2022, 831, 154877. [Google Scholar] [CrossRef]
- Eskelson, B.N.; Anderson, P.D.; Temesgen, H. Sampling and Modeling Riparian Forest Structure and Riparian Microclimate. In Density Management for the 21st Century: West Side Story; General Technical Report PNW-GTR-880; USDA Forest Service: Washington, DC, USA; Pacific Northwest Research Station: Portland, OR, USA, 2013; pp. 126–135. [Google Scholar]
- Aussenac, G. Interactions between Forest Stands and Microclimate: Ecophysiological Aspects and Consequences for Silviculture. Ann. For. Sci. 2000, 57, 287–301. [Google Scholar] [CrossRef]
- Chirkov, Y.I. Microclimate and Phytoclimate. In Agrometeorology; Seemann, J., Chirkov, Y.I., Lomas, J., Primault, B., Eds.; Springer: Berlin/Heidelberg, Germany, 1979; pp. 139–141. ISBN 978-3-642-67288-0. [Google Scholar]
- Wilken, G.C. Microclimate Management by Traditional Farmers. Geogr. Rev. 1972, 62, 544–560. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L.; Zamora, R.; Gómez, J.M.; Hódar, J.A.; Castro, J.; Baraza, E. Applying Plant Facilitation to Forest Restoration: A Meta-Analysis of the Use of Shrubs as Nurse Plants. Ecol. Appl. 2004, 14, 1128–1138. [Google Scholar] [CrossRef] [Green Version]
- Simões, M.P.; Belo, A.F.; Fernandes, M.; Madeira, M. Regeneration Patterns of Quercus suber According to Montado Management Systems. Agrofor. Syst. 2016, 90, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Smit, C.; den Ouden, J.; Díaz, M. Facilitation of Quercus ilex Recruitment by Shrubs in Mediterranean Open Woodlands. J. Veg. Sci. 2007, 19, 193–200. [Google Scholar] [CrossRef]
- Skidmore, E.L.; Hagen, L.J. Evaporation in Sheltered Areas as Influenced by Windbreak Porosity. Agricult. Meteorol. 1970, 7, 363–374. [Google Scholar] [CrossRef]
- Callaway, R.M.; Walker, L.R. Competition and Facilitation: A Synthetic Approach to Interactions in Plant Communities. Ecology 1997, 78, 1958–1965. [Google Scholar] [CrossRef]
- Caldeira, M.C.; Lecomte, X.; David, T.S.; Pinto, J.G.; Bugalho, M.N.; Werner, C. Synergy of Extreme Drought and Shrub Invasion Reduce Ecosystem Functioning and Resilience in Water-Limited Climates. Sci. Rep. 2015, 5, 15110. [Google Scholar] [CrossRef] [Green Version]
- Haberstroh, S. Impact of Extreme Drought Events and Shrub Invasion on Mediterranean Cork Oak (Quercus suber L.) Ecosystem Functioning and Recovery. Dissertation, Albert-Ludwigs-Universität Freiburg im Breisgau, Breisgau, Germany, January 2021. Available online: https://www.cep.uni-freiburg.de/forschungsprojekte/idi (accessed on 26 July 2016).
- Arosa, M.L.; Ceia, R.S.; Costa, S.R.; Freitas, H. Factors Affecting Cork Oak (Quercus suber) Regeneration: Acorn Sowing Success and Seedling Survival under Field Conditions. Plant Ecol. Divers. 2015, 8, 519–528. [Google Scholar] [CrossRef]
- Köbel, M.; Listopad, C.M.C.S.; Príncipe, A.; Nunes, A.; Branquinho, C. Temporary Grazing Exclusion as a Passive Restoration Strategy in a Dryland Woodland: Effects over Time on Tree Regeneration and on the Shrub Community. For. Ecol. Manag. 2021, 483, 118732. [Google Scholar] [CrossRef]
- Listopad, C.; Köbel, M.; Príncipe, A.; Goncalves, P.; Branquinho, C. The Effect of Grazing Exclusion over Time on Structure, Biodiversity, and Regeneration of High Nature Value Farmland Ecosystems in Europe. Sci. Total Environ. 2017, 610–611, 926–936. [Google Scholar] [CrossRef]
- González, J.; Vallejo, J.R.; Amich, F.; Cistus Ladanifer, L. Inventario Español de los Conocimientos Tradicionales Relativos a la Biodiversidad; Ministerio de Agricultura y Pesca, Alimentación y Medio Ambiente (MAPAMA): Madrid, Spain, 2018; Volume Fase II (Tomo 2), pp. 47–55. ISBN 978-84-491-1472-4.
- Mendes, P.; Meireles, C.; Vila-Viçosa, C.; Musarella, C.; Pinto-Gomes, C. Best Management Practices to Face Degraded Territories Occupied by Cistus ladanifer Shrublands—Portugal Case Study. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2015, 149, 494–502. [Google Scholar] [CrossRef] [Green Version]
- EEA European Forest Types. Categories and Types for Sustainable Forest Management Reporting and Policy. EEA Tech. Rep. 2006, 9, 1–114. [Google Scholar]
- Carvalho, S. Ficha Informativa do Clima da Herdade da Ribeira Abaixo. In Life Montado Adapt; FCUL: Lisboa, Portugal, 2017. [Google Scholar]
- IPMA Portal do Clima. Available online: http://www.portaldoclima.pt/pt/ (accessed on 26 July 2016).
- DGT. COS-Carta de Uso e Ocupação Do Solo Para 2018; Format Shapefile (Scale 1:25000); Direcção Geral do Território: Lisboa, Portugal, 2018.
- Chen, J.; Saunders, S.C.; Crow, T.R.; Naiman, R.J.; Brosofske, K.D.; Mroz, G.D.; Brookshire, B.L.; Franklin, J.F. Microclimate in Forest Ecosystem and Landscape Ecology: Variations in Local Climate Can Be Used to Monitor and Compare the Effects of Different Management Regimes. BioScience 1999, 49, 288–297. [Google Scholar] [CrossRef] [Green Version]
- Foken, T. Micrometeorology; Springer: Berlin/Heidelberg, Germany, 2017; ISBN 978-3-642-25439-0. [Google Scholar]
- Crowther, T.W.; Glick, H.B.; Covey, K.R.; Bettigole, C.; Maynard, D.S.; Thomas, S.M.; Smith, J.R.; Hintler, G.; Duguid, M.C.; Amatulli, G.; et al. Mapping Tree Density at a Global Scale. Nature 2015, 525, 201–205. [Google Scholar] [CrossRef]
- Príncipe, A.; Matos, P.; Sarris, D.; Gaiola, G.; do Rosário, L.; Correia, O.; Branquinho, C. In Mediterranean Drylands Microclimate Affects More Tree Seedlings than Adult Trees. Ecol. Indic. 2019, 106, 105476. [Google Scholar] [CrossRef]
- Príncipe, A.; Nunes, A.; Pinho, P.; do Rosário, L.; Correia, O.; Branquinho, C. Modeling the Long-Term Natural Regeneration Potential of Woodlands in Semi-Arid Regions to Guide Restoration Efforts. Eur. J. For. Res. 2014, 133, 757–767. [Google Scholar] [CrossRef]
- Chalker-Scott, L. Impact of Mulches on Landscape Plants and the Environment—A Review. J. Environ. Horticult. 2007, 25, 239. [Google Scholar] [CrossRef]
- Jiménez, M.N.; Pinto, J.R.; Ripoll, M.A.; Sánchez-Miranda, A.; Navarro, F.B. Restoring Silvopastures with Oak Saplings: Effects of Mulch and Diameter Class on Survival, Growth, and Annual Leaf-Nutrient Patterns. Agrofor. Syst. 2014, 88, 935–946. [Google Scholar] [CrossRef]
- Rodríguez-Rojo, M.P.; Roig, S.; López-Carrasco, C.; Redondo García, M.M.; Sánchez-Mata, D. Which Factors Favour Biodiversity in Iberian Dehesas? Sustainability 2022, 14, 2345. [Google Scholar] [CrossRef]
- Chaves, N.; Escudero, J.C. Allelopathic Effect of Cistus ladanifer on Seed Germination. Funct. Ecol. 1997, 11, 432–440. [Google Scholar] [CrossRef]
- Alías, J.C.; Sosa, T.; Escudero, J.C.; Chaves, N. Autotoxicity against Germination and Seedling Emergence in Cistus ladanifer L. Plant Soil 2006, 282, 327–332. [Google Scholar] [CrossRef]
- Yeomans, P.A. Water for Every Farm Using the Keyline Plan; Second Back Row Press: Adelaide, Australia, 1981; ISBN 0 90932 529 4. [Google Scholar]
- Raeissi, S.; Taheri, M. Energy Saving by Proper Tree Plantation. Build. Environ. 1999, 34, 565–570. [Google Scholar] [CrossRef]
- Seemann, J.; Chirkov, Y.I.; Lomas, J.; Primault, B. Agrometeorology; Springer: Berlin/Heidelberg, Germany, 1979; ISBN 978-3-642-67288-0. [Google Scholar]
- Silva, J.S.; Catry, F. Forest Fires in Cork Oak (Quercus suber L.) Stands in Portugal. Int. J. Environ. Stud. 2006, 63, 235–257. [Google Scholar] [CrossRef]
- CIFOR Forests and Climate Change Adaptation: What Policymakers Should Know. Available online: https://www.cifor.org/knowledge/publication/4059/ (accessed on 7 February 2021).
- Santos, F.D.; Miranda, P. Alterações Climáticas em Portugal. Cenários, Impactos e Medidas de Adaptação—Projecto SIAM II; Gradiva: Lisboa, Portugal, 2006. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable/Year | 1971–2000 | 2070–2100 (RCP8.5) | 2019 ** | 2020 * | 2021 * |
|---|---|---|---|---|---|
| Annual precipitation (mm) | 828 | 649 | 404.8 | 529.0 | 390.2 |
| Average temperature (°C) | 15.1 | 18.5 | 17.0 | 16.8 | 17.4 |
| Maximum temperature (°C) | - | - | 38.9 | 40.8 | 42.4 |
| Precipitation (mm) | Average Maximum Temperature (°C) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Month | 1971–2000 | 2070–2100 | 2019 ** | 2020 * | 2021 * | 1971–2000 | 2070–2100 | 2019 ** | 2020 * | 2021 * |
| (RCP8.5) | (RCP8.5) | |||||||||
| January | - | - | 43.7 | 35.8 | 53 | - | - | 16.7 | 16.1 | 16.9 |
| February | - | - | 35.1 | 10.8 | 93.2 | - | - | 18.4 | 20.1 | 16.8 |
| March | - | - | 14.4 | 33.4 | 19.6 | - | - | 21.3 | 20.1 | 20.3 |
| April | - | - | 82.0 | 90.8 | 62.6 | - | - | 20.6 | 21.3 | 22.7 |
| May | - | - | 8.0 | 52.2 | 13.2 | - | - | 28.0 | 27.7 | 26.5 |
| June | - | - | 5.4 | 2.4 | 10.2 | - | - | 26.7 | 28.4 | 28.9 |
| July | - | - | 2.8 | 1.0 | 1.6 | - | - | 29.1 | 36.0 | 32.1 |
| August | - | - | 18.8 | 2.6 | 2.4 | 29.1 | 33.1 | 30.9 | 32.0 | 32.5 |
| September | - | - | 5.8 | 11.2 | 26.2 | - | - | 28.5 | 30.1 | 25.9 |
| October | - | - | 19.1 | 101.8 | 32.6 | - | - | 23.3 | 24.0 | 23.7 |
| November | - | - | 112.6 | 93.0 | 19 | - | - | 17.5 | 19.8 | 16.9 |
| December | - | - | 94.5 | 94.2 | 56.6 | - | - | 16.7 | 15.5 | 15.8 |
| Samples of Q. suber | Experimental Design | Plantation | Final Monitoring | |
|---|---|---|---|---|
| Experiment 1 topographic microclimate | 80 saplings planted and monitored (0 damaged) | 4 sites: north, south, water line, ridge. 4 blocks each. | 17 February 2019 | 5 March 2022 (36 months) |
| Experiment 2 shrub rows microclimate | 1200 saplings planted, 972 monitored (228 damaged by wild boar) | 2 sites: 16 blocks without C. ladanifer (A1–4, B1–4, C1–4, D1–4). 16 blocks with C. ladanifer (A1–4, B1–4, C1–4, D1–4) | 21–23 December 2020 | 29 May 2022 (17 months) |
| Planting Sites | Number of Blocks | Total Number of Trees | Survival Percentage | KW | df | p-Value |
|---|---|---|---|---|---|---|
| Water line | 4 | 20 | 45 ± 19.15 | 8.84 | 3 | 0.032 |
| North | 4 | 20 | 35 ± 10 | |||
| South | 4 | 20 | 15 ± 19.15 | |||
| Ridge | 4 | 20 | 5 ± 10 |
| N of Blocks (with C. ladanifer/ without C. ladanifer) | N of Total Samples (with C. ladanifer/without C. ladanifer) | Measurements | |||||
|---|---|---|---|---|---|---|---|
| With C. ladanifer | Without C. ladanifer | W | p-Value | ||||
| Survival percentage (%) | All treatments | 16/16 | 438/534 | 46.08 ± 14.09 [38.32, 53.84, 3.64] | 34.68 ± 15.48 [26.16, 43.2, 4] | 69 | 0.027 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vizinho, A.; Príncipe, A.; Vasconcelos, A.C.; Rebelo, R.; Branquinho, C.; Penha-Lopes, G. Using and Creating Microclimates for Cork Oak Adaptation to Climate Change. Land 2023, 12, 531. https://doi.org/10.3390/land12030531
Vizinho A, Príncipe A, Vasconcelos AC, Rebelo R, Branquinho C, Penha-Lopes G. Using and Creating Microclimates for Cork Oak Adaptation to Climate Change. Land. 2023; 12(3):531. https://doi.org/10.3390/land12030531
Chicago/Turabian StyleVizinho, André, Adriana Príncipe, Ana Cátia Vasconcelos, Rui Rebelo, Cristina Branquinho, and Gil Penha-Lopes. 2023. "Using and Creating Microclimates for Cork Oak Adaptation to Climate Change" Land 12, no. 3: 531. https://doi.org/10.3390/land12030531