1. Introduction
The Great Basin is a region of north and south running mountain ranges and valleys that extend from the Wasatch Mountains of Utah in the east to the Sierra Nevada Mountains in the west. The northern border is the Snake River Plain extending south to the Mohave Desert. It is characterized as a cold desert with hot summers and freezing winters. Precipitation ranges from 125 to 500 mm annually [
1]. Two common habitat types found throughout the region are pinyon/juniper (
Pinus edulis Engelm. or
P. monophylla Torr. and Frem. and
Juniperus osteosperma (Torr.) Little) woodland and sagebrush (
Artemisia tridentata Nutt.) steppe. In Utah, these two habitat types compose nearly 40 percent of the semi-arid region of the state [
2]. These habitats have become fragmented due to frequent wildfires, leaving a habitat mosaic across the region. Subsequent to wildfire, they are often replaced by introduced exotic species, especially annual cheatgrass (
Bromus tectorum L.), which is native to Eurasia. A regional model suggests that nearly one-third of the Great Basin (210,000 km
2) has cheatgrass cover of at least 15 percent [
3]. Crested wheatgrass (
Agropyron cristatum (L.) Gaertn.), a perennial grass native to Russian and Siberia, has been used extensively in post wildfire reseeding efforts across the region. While it is an introduced species, it is considered preferable to cheatgrass. Like cheatgrass, it forms monotypic stands. The impact of these habitat alterations from native to nonnative plant communities on insect communities is poorly understood. We are interested in the effect this might have on chalcidid wasps that occur in these habitats.
The family Chalcididae is a taxon of parasitic Hymenoptera. These wasps most often parasitize pupae of Lepidoptera [
4]. As such, they are typically studied for their potential use as biological control agents in areas where their lepidopteran hosts are important economic insect pests. The taxonomy of Chalcididae has changed very little in the past 150 years, including divisions into subfamilies and genera [
4], a 1992 revision of the new world Chalcididae being the most recent [
5]. Recently, three species of chalcidid wasps were described from rangelands of the eastern Great Basin in Utah, two of which were collected in enough abundance to warrant further investigation [
6]. These two species belong to the subfamily Haltichellinae.
Haltichellinae is comprised of six chalcidid genera that occur in the United States and Canada [
4]. The most recent report of Haltichellinae species that occur in the Great Basin describes the distribution of species in the genus
Psilochalcis Kieffer [
6].
The Universal Chalcidoidea Database [
7] currently reports 60 species of
Psilochalcis worldwide. Old world and new world distributions are not known to overlap [
8]. The literature suggests that less than 20 species of
Psilochalcis are known from the Western Hemisphere [
5]. In general,
Psilochalcis species occur in arid to semi-arid areas across the southwest and western United States [
6]. Biological observations are rare, with only a few host species reported [
8,
9].
Of the eight species of
Psilochalcis wasps known to occur in the United States [
6], four are associated with agricultural environments.
Psilochalcis brevialata Grissell and Johnson is known from a culled fig warehouse in California [
8].
Psilochalcis deceptor (Grissell and Schauff),
P. threa (Grissell and Schauff), and
P. usta (Grissell and Schauff) occur in cultivated peanut crop in Oklahoma and Texas [
9].
Psilochalcis hesphenheidei (Boŭcek) occurs in natural areas across the western United States [
10], but no habitat associations are reported.
Psilochalcis adenticulata Petersen is known from multiple natural habitat types based on label information from collection sites in New Mexico, Nevada, and Utah [
6].
The paucity of ecological data associated with Psilochalcis is understandable, due to their being rarely collected and the lack of interest in adding ecological data to collection labels. This paper is unique, relative to all other publications on North American Psilochalcis, because it links two Psilochalcis species with specific ecological data. Using Malaise trap capture data, we evaluate the changes in seasonal abundance over time for P. minuta Petersen and P. quadratis Petersen and their associations within distinct habitat types of the eastern Great Basin.
2. Materials and Methods
In 2019, we were able to examine and extract chalcid wasps from historic Malaise trap samples collected in 2006 and 2007. These samples were originally used to study different insect groups with the remaining material stored in 500 ml Nalgene bottles with 70% ethanol at 2 °C. The sample sites were originally selected in areas where native and non-native plant communities formed contiguous boundaries representing; native shrubland, native woodland, introduced annual grassland, and introduced perennial grassland.
2.1. Site Descriptions
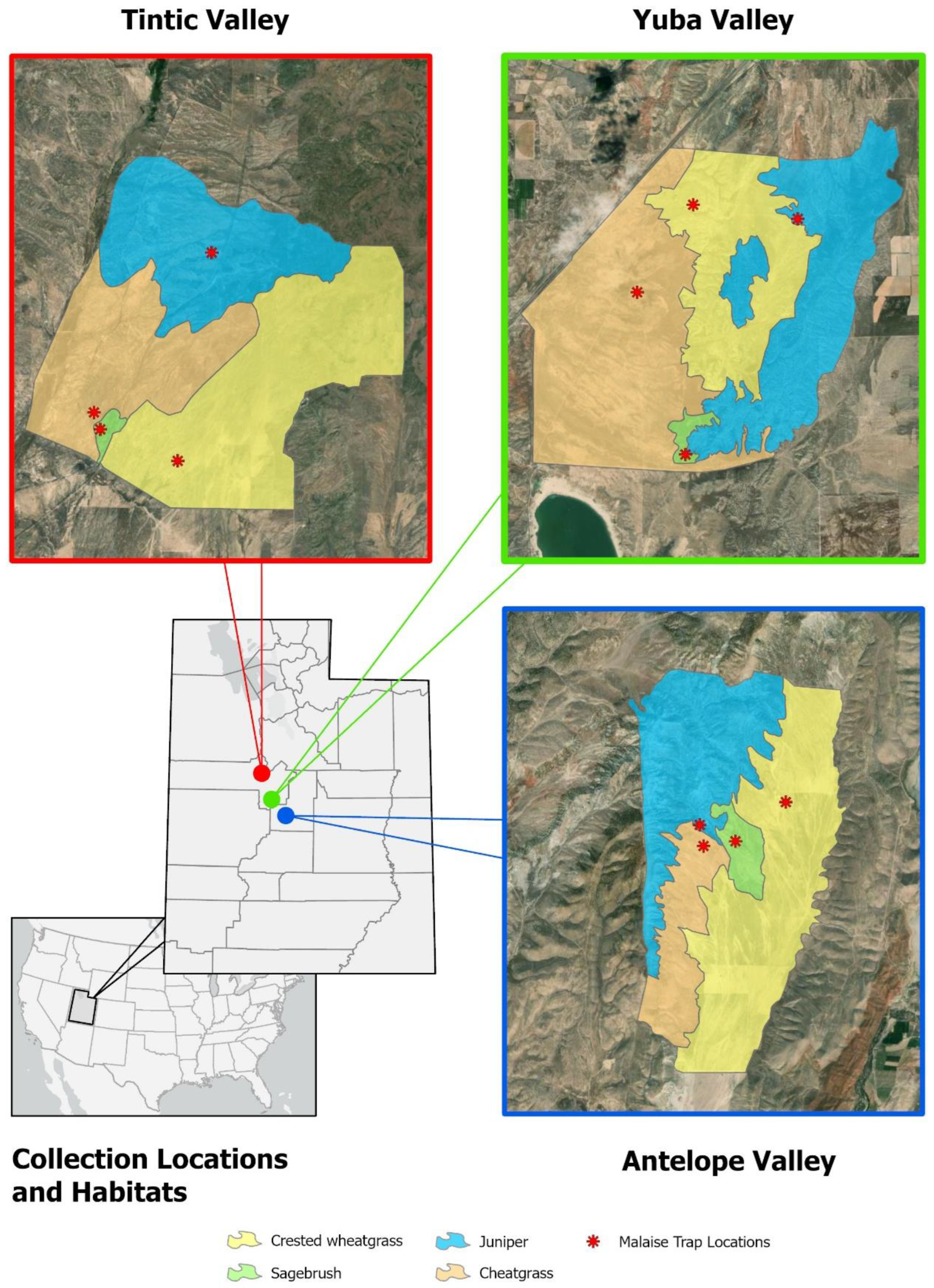
In 2006, a study was established to examine insect diversity in native pinyon/juniper and sagebrush habitats relative to those in the non-native conversion habitats of cheatgrass or crested wheatgrass. From these samples, we are able to test the effect of habitat on the presence of
Psilochalcis wasps. Malaise traps were set up in four contiguous habitat types common to the Great Basin at three different locations. Locations are (1) Utah, Juab County, Tintic Valley; Utah, Juab County, Yuba Valley; and Utah, Sanpete County, Antelope Valley. The habitat types at these locations are defined by the dominant plant species, namely (1) pinyon/juniper (
Pinus edulis or
P. monophylla and
Juniperus osteosperma), (2) sagebrush (
Artemisia tridentata), (3) cheatgrass (
Bromus tectorum) and (4) crested wheatgrass (
Agropyron cristatum) (
Figure 1). Both the cheatgrass and crested wheatgrass sites were either native shrubland or woodland prior to wildfires. Crested wheatgrass sites were reseeded after fire disturbance whereas cheatgrass sites were untreated or failed reseedings. These two altered plant communities formed the dominant habitat type at each location with sagebrush comprising much smaller remnant patches. The overall topography was relatively flat except where dry washes bisected the study site or where slopes gradually gain elevation. The Antelope Valley site has abundant pinyon/juniper on the adjacent slopes to the west and north. At the Tintic Valley site, pinyon/juniper is abundant to the north, and at the Yuba Valley site, pinyon/juniper is abundant to the east (
Figure 1). Within habitat types, exact trap locations were based partly on road accessibility and the logistics of regular retrieval of trap samples. Though some trap sites appear close to habitat boundaries, they were still placed a minimum distance of 100 meters from the habitat edge, to reduce the effect of edge bias.
2.2. Sampling Method
Townes-style malaise traps [
11,
12] were installed at three different sites. At each site, three traps were set up in each of the four habitats. Within each habitat, the three traps were installed 120° opposing each other thus effecting a full 360° sample orientation. A total of 36 traps were installed in 2006 and repeated in 2007. Samples from each trap were retrieved biweekly from 1 April to 1 October (Spring through Fall). Trap setup required two eight-foot T-posts pounded into the soil on either side of trap, securing trap to posts, and staking the trap to the ground (
Figure 2).
Malaise trapping is the prevailing method for collecting chalcidid wasps [
13,
14,
15]. Malaise traps have been shown to be more effective in trapping chalcidoids than other style traps and are preferred when time and/cost are major constraints [
16]. Due to the remoteness of our trap locations, it was not feasible to check traps more frequent than biweekly. Malaise traps passively capture through flight interception, which allows to them to be left unattended between sample retrieval for longer periods relative to other style traps without the concern of sample degradation.
Preliminary examination of chalcidid wasp captures indicated two Psilochalcis species almost exclusively occurring in pinyon/juniper and cheatgrass habitats. This discovery prompted us to set up a few additional traps during the summers of 2020 through 2022 to see if we could predictably recapture the two wasp species in the same habitats but from different locations. Eleven traps were set up in pinyon/juniper and four traps in cheatgrass.
2.3. Plant Composition
In 2007, aerial plant cover was estimated at each trap location using a half-square meter quadrat placed every 5 m (excluding point 0) along a 45 m transect in each cardinal direction from the Malaise trap center. This yielded 32 sample quadrats for each trap with the sample area being a 45-m radius around each trap.
2.4. Seasonal Abundance
Malaise trap data were used to calculate species trap abundance by location, habitat type and collection year. For each Psilochalcis species, we produced annual species abundance graphs using Microsoft Excel 2016 to visualize seasonal trends.
2.5. Statistical Analysis
The trap abundance data for both P. minuta and P. quadratis were highly skewed having a large number of zeros from several traps across habitat type, site, and collection year. The data were transformed to a log scale before analysis to account for this. A separate two-way analysis of variance (ANOVA) and logistic regression were conducted for P. minuta and P. quadratis using the Statistical Analysis System (SAS) version 9.4 to determine the likelihood of each species occurring at each location and habitat type.
2.6. Climate Data Analysis
To assess any impact of climate on seasonal abundance, we analyzed both county [
17] and local [
18] temperature and precipitation data to visualize climate trends at collection sites. The weather stations in closest proximity to each location were chosen. Stations are Little Sahara for Tintic Valley, Scipio for Yuba Valley, and Manti for Antelope Valley.
3. Results
3.1. Seasonal Trap Abundance
In 2006, 99.24 percent of
P. minuta specimens were collected in pinyon/juniper habitat, while 0.76 percent of specimens were collected in crested wheatgrass habitat. Total number of specimens collected was 131, and only at the Yuba Valley site. In 2006, 96.75 percent of
P. quadratis specimens were collected in cheatgrass habitat, 3.0 percent in crested wheatgrass habitat and 0.25 percent in sagebrush habitat. Total number of specimens collected was 401, with 400 specimens collected at Yuba Valley, and 1 collected at Tintic Valley. The 2006 seasonal trap abundance for both
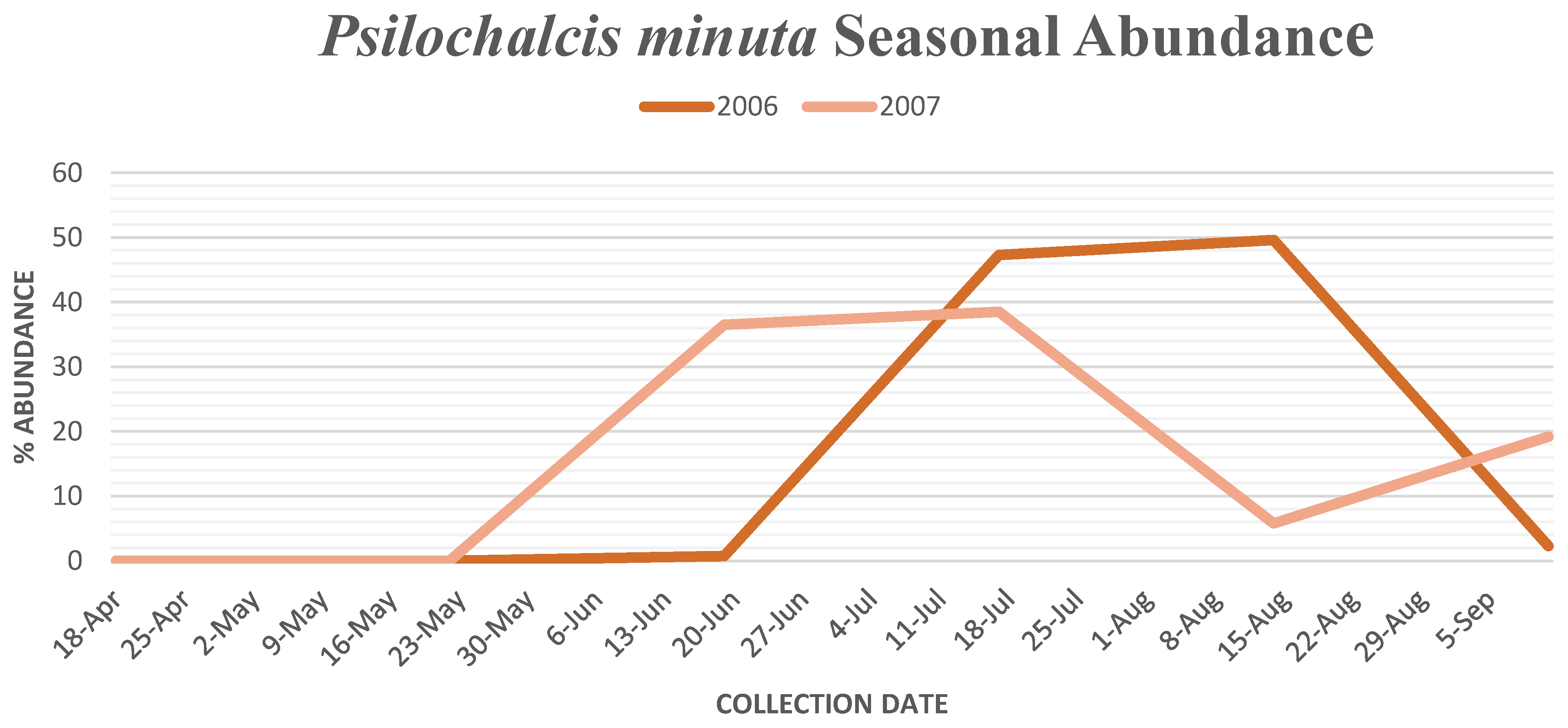
P. minuta (
Figure 3) and
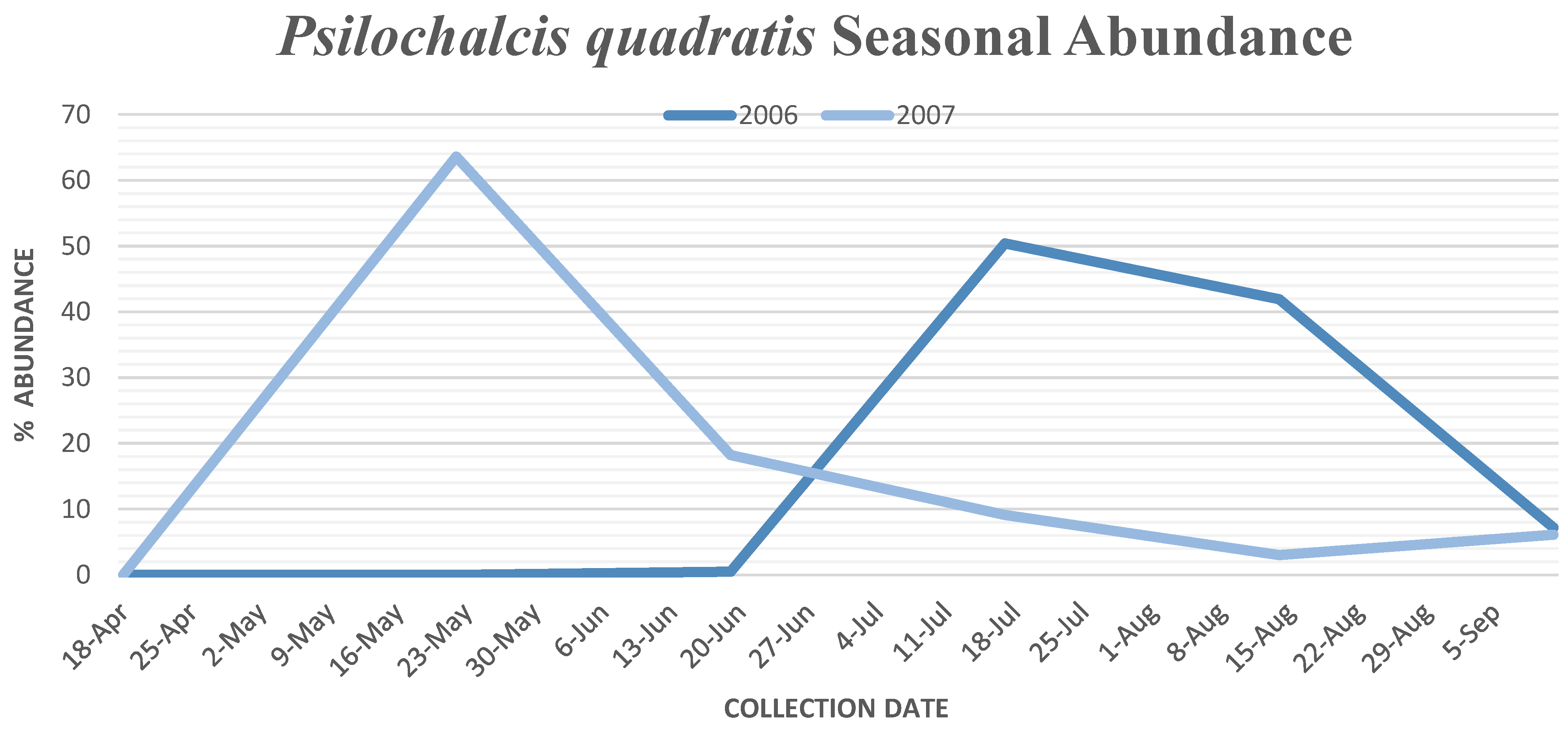
P. quadratis (
Figure 4) peaked from mid-July to early August.
Collection data for 2007 showed a decrease in seasonal abundance and a time shift in the peak abundance for both
Psilochalcis species. The total number of
P. minuta specimens collected in 2007 decreased to 52, with all specimens collected in pinyon/juniper habitat at Yuba Valley. The seasonal abundance for
P. minuta peaked one month earlier in mid-June (
Figure 3). The total number of
P. quadratis specimens collected decreased to 33, with 75.8 percent of specimens collected in cheatgrass habitat, 12.1 percent in crested wheatgrass habitat and 12.1 percent in sagebrush habitat. Thirty
P. quadratis specimens were collected at Yuba Valley, with 2 specimens collected at Antelope Valley and 1 specimen collected at Tintic Valley. The seasonal abundance for
P. quadratis peaked two months earlier in mid-May (
Figure 4).
Subsequent Malaise trapping from other locations in 2020 through 2022 revealed similar Psilochalcis affinities to habitat. Psilochalcis minuta was predominantly found in traps placed in pinyon/juniper habitat. All eleven traps placed in pinyon/juniper yielded specimens of P. minuta totaling 348. Five P. minuta specimens were retrieved from two traps placed in cheatgrass habitat. Three of four traps placed in cheatgrass habitat yielded specimens of P. quadratis totaling 10. Additionally, 18 P. quadratis specimens were retrieved from five traps placed in pinyon/juniper.
3.2. Plant Composition
Pinyon/juniper habitat sites are characterized by the dominant species Utah juniper (
Juniperus osteosperma). The percent cover of Utah Juniper was 10.9% at Tintic Valley, 19.4% at Yuba, and 17.3% at Antelope Valley. Only the Antelope Valley site had cover of pinyon pine (Pinus edulis) at 6.0%. Cheatgrass is present in the understory at all three locations but with extremely low cover. Tintic Valley had cheatgrass cover of 0.1%, Yuba Valley had 1.5% and Antelope Valley had 0.1%. Other plants varied amongst the three locations (
Table 1).
Cheatgrass habitat sites are characterized by the dominant species cheatgrass (
Bromus tectorum). The percent cover of cheatgrass at Tintic Valley was 16.1%, 1.3% at Yuba, and 42% at Antelope Valley.
Eremopyrum triticeum (Gaertn.) Nevski had a slightly greater percent cover than cheatgrass at the Yuba site. It should be noted that the Yuba cheatgrass site experienced cheatgrass die-off in 2007, a phenomenon that occurs sporadically but regularly throughout the Great Basin [
19]. No other species were sampled at the Antelope Valley site. Other plants varied amongst the other two locations (
Table 2).
In general, sagebrush habitat sites are characterized by the dominant species Big Sagebrush (Artemisia tridentata) comprising slightly more than 12% of the total plant cover. Cheatgrass (Bromus tectorum) is present in the understory comprising approximately 7.5% total cover. Other forbes found were Elymus elymoides and Chrysothamnus viscidiflorus, both approximately 0.3% total cover.
Crested wheatgrass habitat sites are characterized by the dominant species Crested wheatgrass (Agropyron cristatum) comprising slightly more than 4% of the total plant cover. Cheatgrass (Bromus tectorum) is present in the understory comprising approximately 0.7% total cover. Other forbes found were Stipa hymenoides, slightly less than 1%, and Elymus elymoides, less than 0.5 % total cover.
3.3. Statistical Results
Psilochalcis minuta demonstrated a significant interaction in trap abundance for habitat type by location (F6, 60 = 120.97 (p < 0.0001). The least square means analysis shows a significant interaction with pinyon/juniper habitat at Yuba Valley (p < 0.0001, t value 38.53). All other habitat type and location combinations were insignificant. An odds ratio estimate was not obtained for P. minuta due to it being collected only at the Yuba Valley location.
Psilochalcis quadratis also demonstrated a significant interaction in trap abundance for habitat type by location (F6, 60 = 20.06 (p < 0.0001). The least square means combination of cheatgrass at Yuba Valley shows a significant interaction of these two variables (p < 0.0001, t value 16.83). Crested wheatgrass at Yuba Valley also shows significance (p < 0.0001) but with a much lower t value (t value 5.07). With zeros so prevalent in our data set, this significant interaction between Crested wheatgrass habitat at Yuba Valley is to be expected. The number of P. quadratis specimens collected from Crested wheatgrass habitat at Yuba Valley were 16 out of 401. Psilochalcis quadratis was collected at all three locations. From the logistic regression we obtained an odds ratio estimate that P. quadratis is 63.46 times more likely to be present at Yuba Valley than Antelope Valley (confidence interval from 6.13 to 656.88), and 29.73 times more likely to be present at Yuba Valley than Tintic Valley (confidence interval from 4.53 to 195.08).
3.4. Climate Data Results
For Juab County Utah, monthly mean temperatures for 2006 and 2007 were compared to the 20th century average. In 2006, February and March temperatures were at or slightly below the 20th century average. April through July temperatures were consistently warmer. In 2007, the February temperature was slightly above average and rose more above average in March. April through July temperatures were consistently above average. The temperatures for these months were slightly higher when compared to the same months in 2006. In 2006, monthly precipitation was greater from February through September when compared to 2007.
The three weather stations closest to each of our collection sites showed the same trends with minor differences becoming apparent. In 2006, all three stations were lower than the 30-year normal, the greatest deviation occurred at Little Sahara, which was 5.5 degrees below the 30-year normal. Scipio recorded a difference of 3.8 degrees and Manti was 4.1 degrees lower. In 2007, all three stations were higher than the 30-year normal. Temperatures ranged from 4.6 degrees higher at Manti up to 5.0 higher at Little Sahara. April through July in both years were consistently above the 30-year normal. The 30-year normal, 2006, and 2007 mean maximum temperatures from July through December were very similar with only minor fluctuations. The three weather stations reported lower monthly precipitation than the 30-year normal. All three stations also reported slightly higher precipitation in March and April of 2006 compared to the same months in 2007. Scipio showed the greatest difference between years, followed by Manti. Overall, the precipitation records for Little Sahara were spotty for both years, so a comparison was not easily made.
4. Discussion
We collected P. minuta predominantly from areas of pinyon/juniper habitat, with rare collections in cheatgrass. Psilochalcis quadratis was collected primarily from cheatgrass habitat, with occasional collections in pinyon/juniper. The occasional occurrence of Psilochalcis species in adjacent habitat types is either due to their close proximities or the host plant of their host moth co-occurring in both habitats. Both Psilochalcis species are highly mobile. There is no reason not to expect flight across plant community boundaries. Our results suggest biological and/or ecological factors are driving the associations of P. minuta within pinyon/juniper habitat and P. quadratis within cheatgrass habitat and not trap placement within the habitat, but we are uncertain what the key host plant is within either habitat.
Plant composition in the same habitat across sites was not always uniform. The variability seen in trap capture between sites is likely due to differences in plant community compositions. The presence or absence of a particular plant species in a given habitat could be the primary factor influencing the presence of P. minuta and P. quadratis in those areas. This might account for the stark absence of P. minuta and P. quadratis in Antelope Valley, it being the most different in plant composition.
Psilochalcis are known parasitoids of lepidopterans in the families Pyralidae and Gelechiidae [
4]. One particular species,
P. brevialata, has been documented parasitizing the pupal stage of pyralid moths infesting stored figs [
20]. Reports of host associations between dominant plant species occurring in our sampled habitats and pyralid moths are lacking, however; it is interesting to note that the pyralid
Dioryctria albovitella (Hulst) was reported as attacking pinyon pine [
21]. The pupal stage of
D. albovitella has been documented as occurring from mid-July through September [
22]. This timing of pupal abundance coincides with the peak abundance of
P. minuta and
P. quadratis adult wasps observed in our study. We observed both pyralid and gelichiid moth species in the same Malaise trap samples in which
P. minuta and
P. quadratis specimens were retrieved though they are yet to be quantified or identified to species. They were similarly observed in our subsequent Malaise trap samples. It is therefore likely that the abundance of
P. minuta in pinyon/juniper habitat and
P. quadratis in cheatgrass habitat is directly related to the abundance of their lepidopteran hosts occurring in each of these habitat types.
There are many variables affecting the phenology of living organisms, in particular environmental factors. Temperature and precipitation have been shown to play a role in the phenology of plants [
23]. This pattern extends to plant/insect associations. The countywide differences in temperature and precipitation between 2006 and 2007 could account for the shift in observed phenology in
P. minuta and
P. quadratis. In 2007, warmer than average temperatures were recorded in February and March as well as from July through September when compared to 2006. Less than average precipitation was recorded from January through October when compared to 2006. We hypothesize that the warmer and drier spring of 2007 accelerated the phenology of the plant communities at the Yuba location. This could have altered the timing of the peak abundance of the host moth pupae within those habitats, and in turn resulted in the shift in seasonal abundance observed for both
P. minuta and
P. quadratis that parasitize those moths.
5. Conclusions
Psilochalcis minuta and P. quadratis are the first North American Psilochalcis species for which seasonal abundance has been associated with a specific habitat type. While P. minuta is strongly associated with pinyon/juniper habitat and P. quadratis is with cheatgrass habitat, the exact plant species accounting for these relationships remains unknown. The associations with habitat type were however, further confirmed by the additional Malaise traps placed in multiple areas beyond the original trap sites in multiple subsequent years.
We recognize there are still many unknown ecological relationships of Psilochalcis species. While Malaise traps can be a useful tool for determining species/habitat associations, trap costs and trap maintenance are often unfeasible. The simple addition of habitat information to collection labels would significantly contribute to our increased understanding of Psilochalcis and our ability to predict future occurrences through the development of species distribution models. We encourage collectors to include ecological data along with location data to increase our understanding of the ecological relationships of Psilochalcis species in their natural habitats. This study uniquely ties two Psilochalcis species’ seasonal abundance to particular habitats of the Great Basin.






{kind=link}
{kind=link}
{kind=link}
{kind=link}