
Ecological Niche Modelling and Potential Distribution of Artemisia sieberi in the Iranian Steppe Vegetation
 ,
, 
Abstract
:1. Introduction
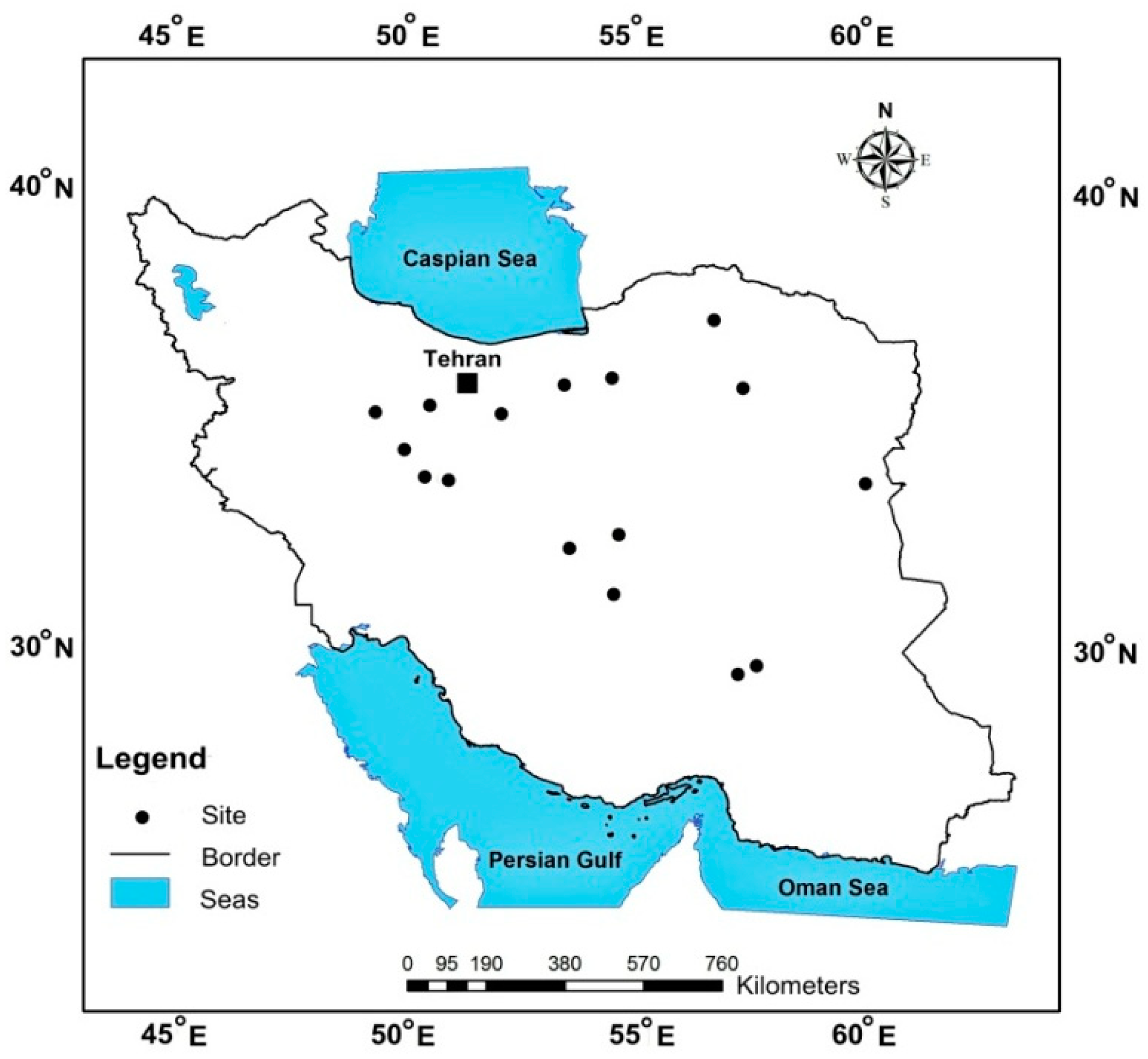
2. Materials and Methods
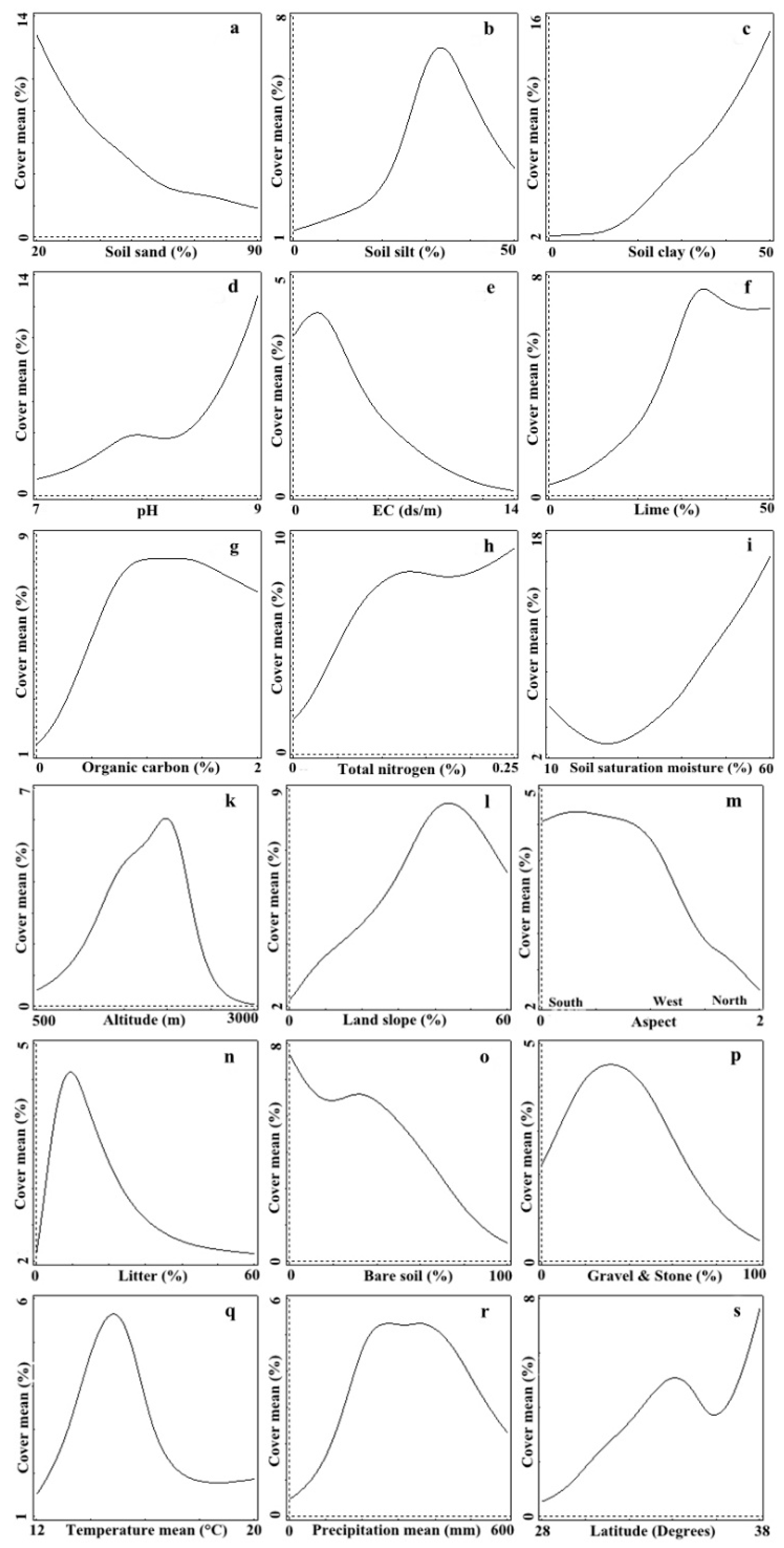
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Podlech, D. Artemisia. In Flora Iranica, Compositae, VI-Anthemideae, Akademische Druck-u; Rechinger, K.H., Ed.; Verlagsanstalt: Graz, Austria, 1986; Volume 158, pp. 159–223. [Google Scholar]
- Mahboubi, M. Artemisia sieberi Besser essential oil and treatment of fungal infections. Biomed. Pharmacother. 2017, 89, 1422–1430. [Google Scholar] [CrossRef] [PubMed]
- Jalili, A. Ecology, Evolution and Biogeography of Artemisia L.; Research Institute of Forests and Rangelands: Tehran, Iran, 2016; p. 493. [Google Scholar]
- Nigam, M.; Atanassova, M.; Mishra, A.P.; Pezzani, R.; Devkota, H.P.; Plygun, S.; Salehi, B.; Setzer, W.N.; Sharifi-Rad, J. Bioactive Compounds and Health Benefits of Artemisia Species. Nat. Prod. Commun. 2019, 14, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, S.; Kappas, M.; Chahouki, M.A.Z.; Gerold, G.; Erasmi, S.; Emam, A.R. Modelling potential habitats for Artemisia sieberi and Artemisia aucheri in Poshtkouh area, central Iran using the maximum entropy model and geostatistics. Ecol. Inform. 2013, 18, 61–68. [Google Scholar] [CrossRef]
- Akhani, H.; Mahdavi, P.; Noroozi, J.; Zarrinpour, V. Vegetation patterns of the Irano-Turanian steppe along a 3000 m al-titudinal gradient in the Alborz Mountains of Northern Iran. Folia. Geobot. 2013, 48, 229–255. [Google Scholar] [CrossRef]
- Whittaker, R.H. Direct Gradient Analysis. Handbook of Vegetation Science 5: Ordination and Classification of Communities; Whittaker, R.H., Ed.; Junk Publishers: Hague, The Netherlands, 1973; pp. 19–50. [Google Scholar]
- Traoré, S.; Zerbo, L.; Schmidt, M.; Thiombiano, L. Acacia communities and species responses to soil and climate gradients in the Sudano-Sahelian zone of West Africa. J. Arid. Environ. 2012, 87, 144–152. [Google Scholar] [CrossRef]
- Gogina, M. Investigation of Interrelations between Sediment and Near-Bottom Environmental Parameters and Macrozoobenthic Distribution Patterns for the Baltic Sea. Ph.D. Thesis, Ernst Moritz Arndt University of Greifswald, Greifswald, Germany, 2010. [Google Scholar]
- Moisen, G.G.; Frescino, T.S. Comparing five modelling techniques for predicting forest characteristics. Ecol. Model. 2002, 157, 209–225. [Google Scholar] [CrossRef] [Green Version]
- Cajo, J.F.; Ter Braak, C. Canonical Correspondence Analysis: A New Eigenvector Technique for Multivariate Direct Gradient Analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef] [Green Version]
- Austin, M.; Belbin, L.; Meyers, J.; Doherty, M.; Luoto, M. Evaluation of statistical models used for predicting plant species distributions: Role of artificial data and theory. Ecol. Model. 2006, 199, 197–216. [Google Scholar] [CrossRef]
- Hastie, T.; Tibshirani, R. Generalised Additive Models; Chapman and Hall: London, UK, 1990. [Google Scholar]
- Kosicki, J.Z. Generalised Additive Models and Random Forest Approach as effective methods for predictive species density and functional species richness. Environ. Ecol. Stat. 2020, 27, 273–292. [Google Scholar] [CrossRef]
- Yee, T.W.; Mitchell, N.D. Generalized additive models in plant ecology. J. Veg. Sci. 1991, 2, 587–602. [Google Scholar] [CrossRef]
- Austin, M.P. The potential contribution of vegetation ecology to biodiversity research. Ecography 1999, 22, 465–484. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Leathwick, J.R.; Whitehead, D. Soil and atmospheric water deficits and the distributions of New Zealand’s indigenous tree species. Funct. Ecol. 2001, 15, 233–242. [Google Scholar]
- Vetaas, O.R.; Grytnes, J.A. Distribution of vascular plant species richness and endemic richness along the Himalayan elevation gradient in Nepal. Glob. Ecol. Biogeogr. 2002, 11, 291–301. [Google Scholar] [CrossRef]
- Austin, M.P.; Meyers, J.A. Current approaches to modelling the environmental niche of eucalypts: Implication for management of forest biodiversity. For. Ecol. Manag. 1996, 85, 95–106. [Google Scholar] [CrossRef]
- Azarnivand, H.; Jafari, M.; Moghaddam, M.R.; Jalili, A.; Zare Chahouki, M.A. Investigation on environmental variables affecting habitat distribution of A. sieberi and A. aucheri in rangelands of Vardavard, Garmsar and Semnan. Iran. J. Nat. Resour. 2006, 56, 93–99. [Google Scholar]
- Yaghmaei, L.; Soltani Koupaei, S.; Khodagholami, M. Effect of climatic factors on distribution of Artemisia sieberi and Artemisia aucheri in Isfahan province using multivariate statistical methods. Water Soil Sci. 2008, 12, 359–371. [Google Scholar]
- Abdollahi, J.; Naderi, H. Soil and topographical variation influencing the growing factors of Artemisia sieberi in steppic rangeland, Nodoushan-Yazd. Watershed Manag. Res. Pajouhesh Sazandegi 2011, 97, 52–62. [Google Scholar]
- Sahragard, H.P.; Ajorlo, M.; Karami, P. Modeling habitat suitability of range plant species using random forest method in arid mountainous rangelands. J. Mt. Sci. 2018, 15, 2159–2171. [Google Scholar] [CrossRef]
- Chahouki, M.A.Z.; Azarnivand, H.; Jafari, M.; Tavili, A. Multivariate statistical methods as a tool for model-based prediction of vegetation types. Russ. J. Ecol. 2010, 41, 84–94. [Google Scholar] [CrossRef]
- Piri Sahragard, H.; Zare Chahouki, M.A. Modeling of Artemisia sieberi Besser habitat distribution using maximum entropy method in desert rangelands. J. Rangel. Sci. 2016, 6, 93–101. [Google Scholar]
- Amiri, M.; Tarkesh, M.; Jafari, R.; Jetschke, G. Bioclimatic variables from precipitation and temperature records vs. remote sensing-based bioclimatic variables: Which side can perform better in species distribution modeling? Ecol. Inform. 2020, 57, 101060. [Google Scholar] [CrossRef]
- Jalili, A.; Rabie, M.; Azarnivand, H.; Hodgson, J.G.; Arzani, H.; Jamzad, Z.; Asri, Y.; Hamzehee, B.; Ghasemi, F.; Hejazi, S.H.; et al. Distribution and ecological consequences of ploidy variation in Artemisia sieberi in Iran. Acta. Oecologica 2013, 53, 95–101. [Google Scholar] [CrossRef]
- Mousaei Sanjerehei, M.; Rundel, P.W. The Impact of Climate Change on Habitat Suitability for Artemisia sieberi and Artemisia aucheri (Asteraceae), a Modeling Approach. Pol. J. Ecol. 2017, 65, 97–109. [Google Scholar]
- Khodagholi, M.; Saboohi, R.; Esfahani, E.Z. A comparison of vegetative climate of Artemisia sieberi Besser and Artemisia aucheri Boiss in Iran. Arch. Meteorol. Geophys. Bioclimatol. Ser. B 2021, 146, 921–939. [Google Scholar] [CrossRef]
- Krebs, C.J. Ecological Methodology, 2nd ed.; Addison Wesley Longman: Menlo Park, CA, USA, 1999; p. 620. [Google Scholar]
- Arzani, H.; Abedi, M. Rangeland Assessment: Vegetation Measurement, 2nd ed.; University of Tehran: Tehran, Iran, 2015. [Google Scholar]
- Küchler, A.W.; Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology. Geogr. Rev. 1976, 66, 114. [Google Scholar] [CrossRef]
- Iranica, F.; Druck-u, A.; Rechinger, K.H. (Eds.) Flora Iranica: Facts and Figures and a List of Publications by k.h. Rechinger on Iran and Adjacent Areas; Verlagsanstalt: Graz, Austria, 2015; Volume 181. [Google Scholar]
- Research Institute of Forests and Rangelands. Flora of Iran; Assadi, M., Ed.; Research Institute of Forests and Rangelands: Tehran, Iran, 2019; Volume 149, Available online: https://irannature.areeo.ac.ir/article_119036.html?lang=en (accessed on 25 November 2022).
- Palmer, M.W. Putting Things in Even Better Order: The Advantages of Canonical Correspondence Analysis. Ecology 1993, 74, 2215–2230. [Google Scholar] [CrossRef]
- Beers, T.W.; Dress, P.E.; Wensel, L.C. Aspect trans-formation in site productivity research. J. For. Res. 1966, 64, 691–692. [Google Scholar]
- Bouyoucos, G.J. Hydrometer Method Improved for Making Particle Size Analyses of Soils. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Black, C.A.; Evans, D.D.; Dinauer, R.C. Methods of Soil Analysis; American Society of Agronomy: Madison, NC, USA, 1965. [Google Scholar]
- McLean, E.; Ph, S.; Requirement, L. Methods of Soil Analysis: Part 2; Chemical and Microbiological Properties: Agronomy Monographs, 1982; pp. 199–224. Available online: https://acsess.onlinelibrary.wiley.com/doi/book/10.2134/agronmonogr9.2.2ed (accessed on 25 November 2022).
- Walkley, A.; Black, I.A. Estimation of soil organic carbon by the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Ryan, J.G.; Estefan, G.; Rashid, A. Soil and Plant Analysis Laboratory Manual; ICARDA: Aleppo, Syria, 2002. [Google Scholar]
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar] [PubMed]
- McCune, B.; Mefford, M.J. Multivariate Analysis of Ecological Data, version 4.17; MjM Software; Gleneden, OR, USA, 1999. Available online: https://www.scirp.org/(S(351jmbntvnsjt1aadkposzje))/reference/ReferencesPapers.aspx?ReferenceID=551732 (accessed on 25 November 2022).
- Smilauer, P.; Leps, J. Multivariate Analysis of Ecological Data Using CANOCO 5, 2nd ed.; Cambridge University: Cambridge, UK, 2014. [Google Scholar]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Dawson, C.W.; Abrahart, R.J.; See, L.M. HydroTest: A web-based toolbox of evaluation metrics for the standardized assessment of hydrological forecasts. Environ. Model. Softw. 2007, 22, 1034–1052. [Google Scholar] [CrossRef] [Green Version]
- ter Braak, C.J.F.; Smilauer, P. Canoco, Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination, version 4.5; Microcomputer Power: Ithaca, Greece, 2002. [Google Scholar]
- Gaston, K.J.; Blackburn, T.M. A Critique for Macroecology. Oikos 1999, 84, 353. [Google Scholar] [CrossRef]
- Toner, M.; Keddy, P. River hydrology and riparian wetlands: A predictive model for ecological assembly. Ecol. Appl. 1997, 7, 236–246. [Google Scholar] [CrossRef]
- Wiser, S.K.; Peet, R.K.; White, P.S. Prediction of rare-plant occurrence: A southern Appalachian example. Ecol. Appl. 1998, 8, 909–920. [Google Scholar] [CrossRef]
- Witkowski, E.T.F.; Lamont, B. Does the rare Banksia goodii have inferior vegetative, reproductive or ecological attributes compared with its widespread co-occurring relative B. gardneri? J. Biogeogr. 1997, 24, 469–482. [Google Scholar] [CrossRef]
- Olden, J.D.; Jacksona, D.A. Comparison of statistical approaches for modelling fish species distributions. Freshw. Biol. 2002, 47, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Bagheri, R.; Chaichi, M.R.; Mohseni-Saravi, M.; Amin, G.R.; Zahedi, G. Grazing affects essential of compositions of Artemisis sieberi Besser. Pak. J. Biol. Sci. 2007, 5, 810–813. [Google Scholar] [CrossRef]
- Khosravi Mashizi, A.; Sharafatmandrad, M. Assessing Impact of Anthropogenic Disturbances on Forage Production in Arid and Semiarid Rangelands. J. Rangel. Sci. 2019, 9, 234–245. [Google Scholar]
- Vereecken, H.; Diels, J.; Van Orshoven, J.; Feyen, J.; Bouma, J. Functional evaluation of pedotransfer functions for the estimtion of soil hydraulic properties. Soil Sci. Soc. Am. J. 1992, 56, 1371–1379. [Google Scholar] [CrossRef]
- Aryanpour, H.; Shorafa, M. Cultivation impact on soil available water in different soil textures using pore size distribution. J. Soil Manag. Sustain. Prod. 2013, 3, 131–148. [Google Scholar]
- Wahba, S.A.; Abdel Rahman, S.I.; Cairo, M.Y.T.; Matyn, M.A. Soil moisture, salinity, water use efficiency and sunflower growth as influenced by irrigation bitumen mulch and plant density. Soil Technol. 1990, 3, 33–44. [Google Scholar] [CrossRef]
- Rangel, T.F.L.V.B.; Diniz-Filho, J.A.F.; Bini, L.M. Towards an integrated computational tool water use efficiency and sun-flower growth as influenced by irrigation bitumen mulch and plant density. Soil Technol. 2006, 3, 33–44. [Google Scholar]
- Moradi, P.; Aghajanloo, F.; Moosavi, A.; Monfared, H.; Khalafi, J.; Taghiloo, M.; Khoshzaman, T.; Shojaee, M.; Mastinu, A. Anthropic Effects on the Biodiversity of the Habitats of Ferula gummosa. Sustainability 2021, 13, 7874. [Google Scholar] [CrossRef]
- Janisova, M. Vegetation-environment relationship in dry calcareous grassland. Ekológia-Bratisl 2005, 24, 25–44. [Google Scholar]
- Mahmodi, S.; Hakymian, M. Fundamental of Soil Science; Tehran University: Tehran, Iran, 2007. [Google Scholar]
- Mohtashamnia, S.; Zahedi, G.H.; Arzani, H. An investigation on synecology of semi-steppe vegetation in relation to edaphic and physiographical factors (case study: Eghlid rangelands of Fars). J. Agric. Nat. Resour. 2008, 14, 111–123. [Google Scholar]
- Mseddi, K.; Al-Shammari, A.; Sharif, H.; Chaieb, M. Plant diversity and relationships with environmental factors after rangeland exclosure in arid Tunisia. Turk. J. Bot. 2016, 40, 287–297. [Google Scholar] [CrossRef]
- Brahim, N.; Blavet, D.; Gallali, T.; Bernoux, M. Application of structural equation modeling for assessing relationships between organic carbon and soil properties in semiarid Mediterranean region. Int. J. Environ. Sci. Technol. 2011, 8, 305–320. [Google Scholar] [CrossRef] [Green Version]
- Lado, M.; Paz, A.; Ben-Hur, M. Organic Matter and Aggregate-Size Interactions in Saturated Hydraulic Conductivity. Soil Sci. Soc. Am. J. 2004, 68, 234–242. [Google Scholar] [CrossRef]
- Davies, K.W.; Bates, J.D.; Miller, R.F. Vegetation Characteristics Across Part of the Wyoming Big Sagebrush Alliance. Rangel. Ecol. Manag. 2006, 59, 567–575. [Google Scholar] [CrossRef]
- Alavi, S.J.; Nouri, Z.; Zahedi Amiri, G.H. The response curve of beech tree (Fagus orientalis lipsky.) in relation to enviromental variables using generalized additive model in Khayroud forest, Nowshahr. J. Wood For. Sci. Technol. 2017, 24, 29–42. [Google Scholar]
- Rahimi, A. Autecology of Onobrychis Chorassanica Bunge. In Khorasan Razavi Province. Master’s Thesis, Ferdowsi University of Mashhad, Mashad, Iran, 1999. [Google Scholar]
- Balent, G.; Stafford Smith, D.M. Conceptual model for evaluating the consequences of management practices on the use of pastoral resources. In Proceedings of the fourth International Rangeland Congress, Montpellier, France, 22–26 April 1991; pp. 1158–1164. [Google Scholar]
- Wang, G.; Zhou, G.; Yang, L.; Li, Z. Distribution, species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slopes of Qilianshan Mountains, Gansu, China. Plant Ecol. 2002, 165, 169–181. [Google Scholar] [CrossRef]
- Grytnes, J.A. Ecological interpretations of the mid-domain effect. Ecol. Lett. 2003, 6, 883–888. [Google Scholar] [CrossRef]
- Moghaddam, M.R. Ecology of Terrestrial Plants; Tehran University: Tehran, Iran, 2005. [Google Scholar]
- Chaplygin, V.A.; Rajput, V.D.; Mandzhieva, S.S.; Minkina, T.M.; Nevidomskaya, D.G.; Nazarenko, O.G.; Kalinitchenko, V.P.; Singh, R.; Maksimov, A.Y.; Popova, V.A. Comparison of Heavy Metal Content in Artemisia austriaca in Various Impact Zones. ACS Omega 2020, 5, 23393–23400. [Google Scholar] [CrossRef] [PubMed]
- Mark, A.F.; Dickinson, K.J.M.; Hofstede, R.G.M. Alpine vegetation, plant distribution, life forms, and environments in a humid New Zealand region: Oceanic and tropical high mountain affinities. Arct. Antarct. Alp. Res. 2018, 32, 240–254. [Google Scholar] [CrossRef]
- Ranjbar, A.; Vali, A.; Mokarram, M.; Taripanah, F. Investigating variations of vegetation: Climatic, geological substrate, and topographic factors-a case study of Kharestan area, Fars Province, Iran. Arab. J. Geosci. 2020, 13, 597–614. [Google Scholar] [CrossRef]
- Zare, M.; Mohammady, M.; Pradhan, B. Modeling the effect of land use and climate change scenarios on future soil loss rate in Kasilian watershed of northern Iran. Environ. Earth Sci. 2017, 76, 305. [Google Scholar] [CrossRef]
- Childs, E.C. The Use of Soil Moisture Characteristics in Soil Studies. Soil Sci. 1940, 50, 239–252. [Google Scholar] [CrossRef]
- Vogel, H.J. A numerical experiment on pore size, pore connectivity, water retention, permeability, and solute transport using network models. Eur. J. Soil Sci. 2000, 51, 99–105. [Google Scholar] [CrossRef]
- Carcey Hincz, P.A.; Diaz Aguilar, I. Impact of Grazing on Soil Mesofauna Diversity and Community Composition in Deciduous Forested Rangelands of Northwest Alberta. Available online: https://open.alberta.ca/dataset/303a7691-a457-4b7e-a32b-f39b034060ec/resource/1883d8df-abe6-4848-a1bb-ba6d46cc6f66 (accessed on 7 April 2016).
- Wassie, A.; Sterck, F.J.; Teketay, D.; Bongers, F. Effects of livestock exclusion on tree regeneration in church forests of Ethiopia. For. Ecol. Manag. 2009, 257, 765–772. [Google Scholar] [CrossRef]
- Laris, P.; Wardell, D.A. Good, bad or ‘necessary evil’? Reinterpreting the colonial durning experiment in savanna landscapes of West Africa. Geogr. J. 2006, 172, 271–290. [Google Scholar] [CrossRef]
- Olliff-yang, R.L.; Case, E.J. The effects of leaf litter on germination in the serpentin endemic Boechera constancei. Madroño 2018, 65, 159–167. [Google Scholar]
- Bakåsmoen, H. Endemic Alpine Plant Facing Local Extinction in Context of Climate and Landuse Change in Hardanger, Norway, Nich Modeling of Artemisis Norvegica Fr. Master’s Thesis, University of Bergen, Bergen, Norway, 2021; p. 102. [Google Scholar]
- Austin, M. Spatial prediction of species distribution: An interface between ecological theory and statistical modelling. Ecol. Model. 2002, 157, 101–118. [Google Scholar] [CrossRef]
- Osborne, P.; Alonso, J.; Bryant, R. Modelling landscape-scale habitat use using GIS and remote sensing: A case study with great bustards. J. Appl. Ecol. 2001, 38, 458–471. [Google Scholar] [CrossRef] [Green Version]
- Gustafson, E.J.; Lietz, S.M.; Wright, J.L. Predicting the spatial distribution of aspen growth potential in the upper Great Lakes region. For. Sci. 2003, 49, 499–508. [Google Scholar]
- Holloway, G.J.; Griffiths, G.H.; Richardson, P. Conservation strategy maps: A tool to facilitate biodiversity action planning illustrated using the heath fritillary butterfly. J. Appl. Ecol. 2003, 40, 413–421. [Google Scholar] [CrossRef]
- Cabeza, M.; Araújo, M.B.; Wilson, R.J.; Thomas, C.D.; Cowley, M.J.R.; Moilanen, A. Combining probabilities of occurrence with spatial reserve design. J. Appl. Ecol. 2004, 41, 252–262. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Environmental Factors | Mean ± Stdev. | Environmental Factors | Mean ± Stdev. |
|---|---|---|---|
| Sand (%) | 63.6 ± 12.8 | Altitude (m) | 1796.6 ± 525.7 |
| Silt (%) | 20.6 ± 8.7 | Aspect (converted value of azimuth) | 1.07 ± 0.75 |
| Clay (%) | 17.1 ± 7.5 | Slope (%) | 11.4 ± 9.3 |
| EC (dsm-1) | 2.4 ± 3.9 | Bare soil (%) | 45.4 ± 29.1 |
| pH | 7.9 ± 0.3 | Stone and gravel (%) | 33.1 ± 22.2 |
| Lime (%) | 21.8 ± 8.6 | Litter (%) | 5.4 ± 5.3 |
| Organic carbon (%) | 0.32 ± 0.3 | Mean annual temperature (°C) | 15.3 ± 1.7 |
| Total nitrogen (%) | 0.03 ± 0.3 | Mean annual precipitation (mm) | 185.7 ± 90.3 |
| Soil saturation moisture (%) | 27.7 ± 6.7 | Latitude (Decimal degrees) | 33.69 ± 2.68 |
| Environmental Variable | F * | P * | AIC Value | Environmental Variable | F * | P * | AIC Value |
|---|---|---|---|---|---|---|---|
| Sand (%) | 18.9 | 0.0000 ** | 1840.3 | Altitude (m) | 44.5 | 0.0000 ** | 1590.4 |
| Silt (%) | 87.2 | 0.0000 ** | 1262.3 | Aspect | 4.2 | 0.0001 ** | 2066.8 |
| Clay (%) | 23.2 | 0.0000 ** | 1812.4 | Slope (%) | 24.1 | 0.0000 ** | 1754.8 |
| pH | 6.6 | 0.0003 ** | 2035.7 | Bare soil (%) | 19.2 | 0.0000 ** | 1847.2 |
| EC (dsm-1) | 11.5 | 0.0000 ** | 1966.2 | Stone and gravel (%) | 3.1 | 0.027 * | 2095 |
| Lime percentage (%) | 80.8 | 0.0000 ** | 1296.4 | Litter (%) | 6.4 | 0.0003 ** | 2034.8 |
| Organic carbon (%) | 24.3 | 0.0000 ** | 1771.4 | Mean annual temperature (°C) | 52.4 | 0.0000 ** | 2178.9 |
| Total nitrogen (%) | 24.5 | 0.0000 ** | 1761.6 | Mean annual precipitation (mm) | 125.7 | 0.0000 ** | 1572.6 |
| Soil saturation moisture (%) | 16.1 | 0.0000 ** | 1769.3 | Latitude (Decimal degrees) | 17 | 0.0000 ** | 2594.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirdavoudi, H.; Ghorbanian, D.; Zarekia, S.; Soleiman, J.M.; Ghonchepur, M.; Sweeney, E.M.; Mastinu, A. Ecological Niche Modelling and Potential Distribution of Artemisia sieberi in the Iranian Steppe Vegetation. Land 2022, 11, 2315. https://doi.org/10.3390/land11122315
Mirdavoudi H, Ghorbanian D, Zarekia S, Soleiman JM, Ghonchepur M, Sweeney EM, Mastinu A. Ecological Niche Modelling and Potential Distribution of Artemisia sieberi in the Iranian Steppe Vegetation. Land. 2022; 11(12):2315. https://doi.org/10.3390/land11122315
Chicago/Turabian StyleMirdavoudi, Hamidreza, Darush Ghorbanian, Sedigheh Zarekia, Javad Miri Soleiman, Mashaalaah Ghonchepur, Eileen Mac Sweeney, and Andrea Mastinu. 2022. "Ecological Niche Modelling and Potential Distribution of Artemisia sieberi in the Iranian Steppe Vegetation" Land 11, no. 12: 2315. https://doi.org/10.3390/land11122315