
LABs Fermentation Side-Product Positively Influences Rhizosphere and Plant Growth in Greenhouse Lettuce and Tomatoes
 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Eluate Preparation and Composition
2.2. Experimental Design and Treatments
2.3. Agronomic Paramenters
2.4. Rhizosphere DNA Extraction and Amplification
2.5. Amplicon Sequences Data Preparation
2.6. Statistical Analysis
3. Results
3.1. Effect of the Eluate on Plant Growth and Production
3.1.1. Enhancement of Lettuce Growth
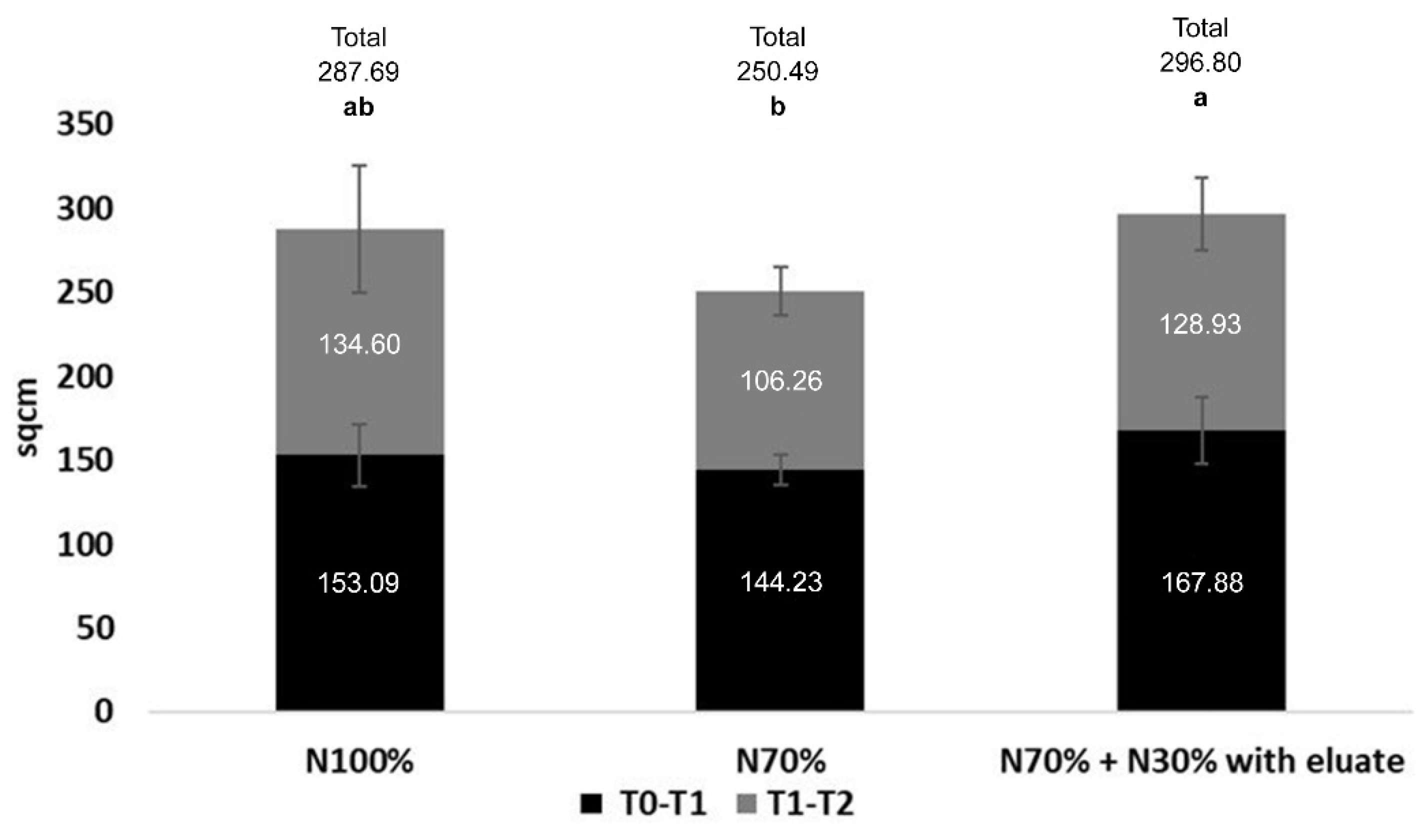
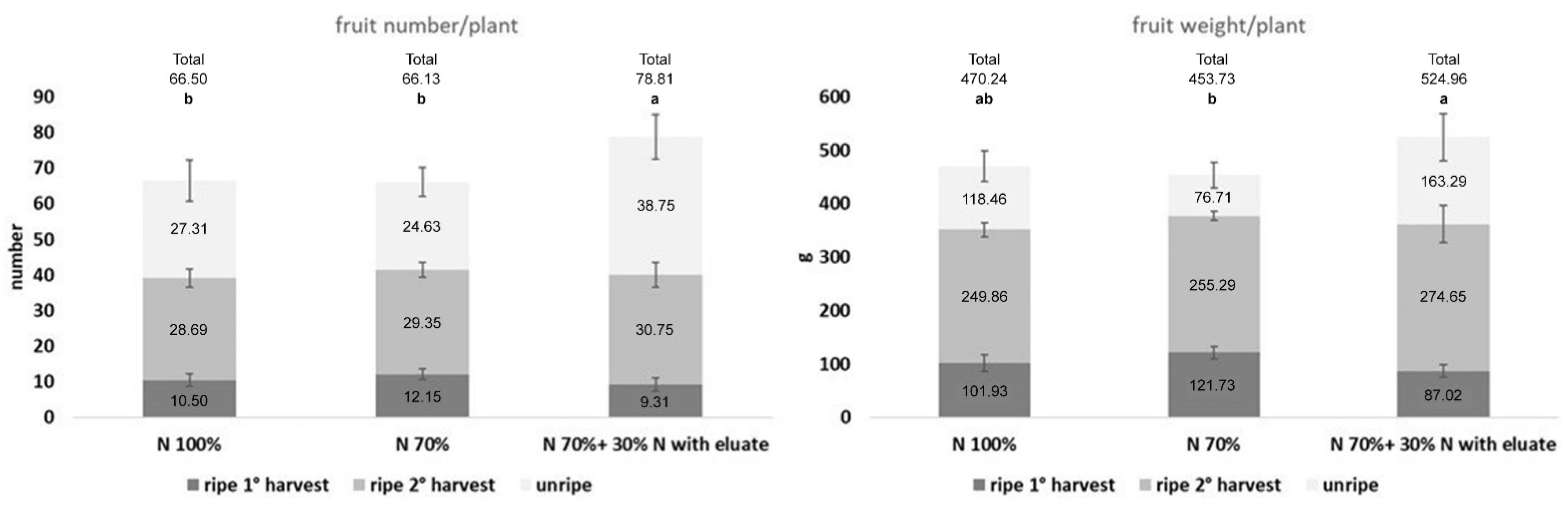
3.1.2. Enhancement of Tomato Growth and Production
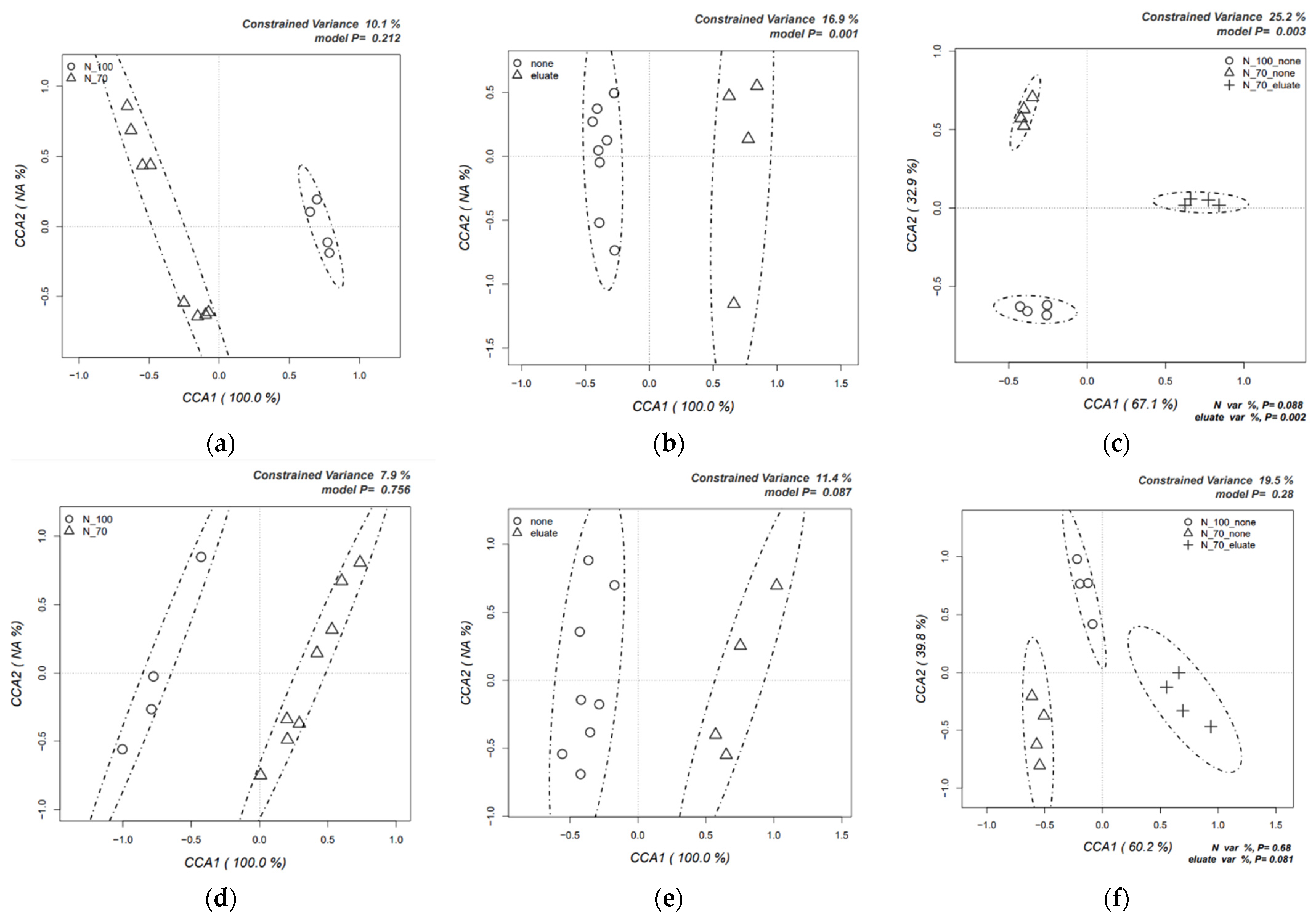
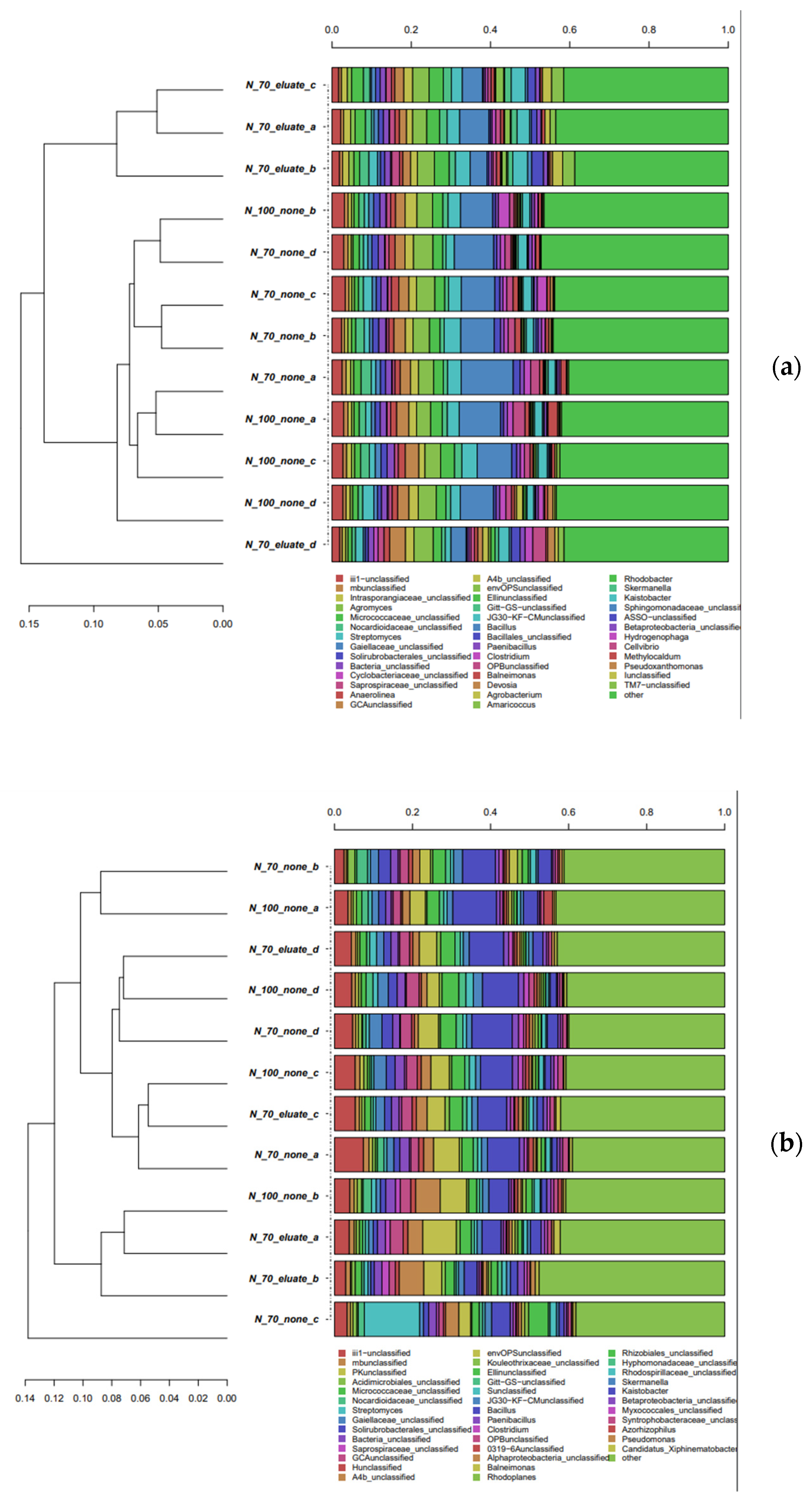
3.2. Dynamics of Rhizosphere Microbial Communities
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Eisa, M.; Ragauskaite, D.; Adhikari, S.; Bella, F.; Baltrusaitis, J. Role and Responsibility of Sustainable Chemistry and Engineering in Providing Safe and Sufficient Nitrogen Fertilizer Supply at Turbulent Times. ACS Sustain. Chem. Eng. 2022, 10, 8997–9001. [Google Scholar] [CrossRef]
- Tyagi, J.; Ahmad, S.; Malik, M. Nitrogenous fertilizers: Impact on environment sustainability, mitigation strategies, and challenges. Int. J. Environ. Sci. Technol. 2022, 1–24. [Google Scholar] [CrossRef]
- Wezel, A.; Herren, B.G.; Kerr, R.B.; Barrios, E.; Gonçalves, A.L.R.; Sinclair, F. Agroecological principles and elements and their implications for transitioning to sustainable food systems. A review. Agron. Sustain. Dev. 2020, 40, 1–13. [Google Scholar]
- Morra, L.; Bilotto, M.; Baldantoni, D.; Alfani, A.; Baiano, S. A seven-year experiment in a vegetable crops sequence: Effects of replacing mineral fertilizers with Biowaste compost on crop productivity, soil organic carbon and nitrates concentrations. Sci. Hortic. 2021, 290, 110534. [Google Scholar] [CrossRef]
- Chirinda, N.; Trujillo, C.; Loaiza, S.; Salazar, S.; Luna, J.; Tong Encinas, L.A.; Lavalle, L.A.B.L.; Tran, T. Nitrous oxide emissions from cassava fields amended with organic and inorganic fertilizers. Soil Use Manag. 2021, 37, 257–263. [Google Scholar] [CrossRef]
- Chen, H.; Zhao, J.; Jiang, J.; Zhao, Z.; Guan, Z.; Chen, S.; Chen, F.; Fang, W.; Zhao, S. Effects of inorganic, organic and bio-organic fertilizer on growth, rhizosphere soil microflora and soil function sustainability in chrysanthemum monoculture. Agriculture 2021, 11, 1214. [Google Scholar] [CrossRef]
- Ganugi, P.; Fiorini, A.; Ardenti, F.; Caffi, T.; Bonini, P.; Taskin, E.; Puglisi, E.; Tabaglio, V.; Trevisan, M.; Lucini, L. Nitrogen use efficiency, rhizosphere bacterial community, and root metabolome reprogramming due to maize seed treatment with microbial biostimulants. Physiol. Plant 2022, 174, e13679. [Google Scholar] [CrossRef]
- Kanwal, Q.; Li, J.; Zeng, X. Mapping recyclability of industrial waste for anthropogenic circularity: A circular economy approach. ACS Sustain. Chem. Eng. 2021, 9, 11927–11936. [Google Scholar] [CrossRef]
- Ren, A.-T.; Abbott, L.K.; Chen, Y.; Xiong, Y.-C.; Mickan, B.S. Nutrient recovery from anaerobic digestion of food waste: Impacts of digestate on plant growth and rhizosphere bacterial community composition and potential function in ryegrass. Biol. Fertil. Soils 2020, 56, 973–989. [Google Scholar] [CrossRef]
- Caballero, P.; Rodríguez-Morgado, B.; Macías, S.; Tejada, M.; Parrado, J. Obtaining plant and soil biostimulants by waste whey fermentation. Waste Biomass Valorization 2020, 11, 3281–3292. [Google Scholar] [CrossRef]
- Arora, N.K.; Mishra, I. United Nations Sustainable Development Goals 2030 and environmental sustainability: Race against time. Environ. Sustain. 2019, 2, 339–342. [Google Scholar] [CrossRef] [Green Version]
- Bastida, F.; Hernández, T.; Garcia, C. Soil degradation and rehabilitation: Microorganisms and functionality. In Microbes at Work; Springer: Berlin/Heidelberg, Germany, 2010; pp. 253–270. [Google Scholar]
- du Jardin, P. The Science of Plant Biostimulants—A Bibliographic Analysis, Ad hoc Study Report; European Commission: Brussels, Belgium, 2012; Available online: https://op.europa.eu/it/publication-detail/-/publication/5c1f9a38-57f4-4f5a-b021-cad867c1ef3c (accessed on 7 September 2017).
- du Jardin, P.; Xu, L.; Geelen, D. Agricultural Functions and Action Mechanisms of Plant Biostimulants (PBs) an Introduction. Chem. Biol. Plant Biostimulants 2020, 1–30. [Google Scholar]
- Bellotti, G.; Taskin, E.; Guerrieri, M.C.; Beone, G.M.; Remelli, S.; Menta, C.; Remelli, S.; Bandini, F.; Tabaglio, V.; Fiorini, A.; et al. Agronomical valorization of eluates from the industrial production of microorganisms: Chemical, microbiological and ecotoxicological assessment of a novel putative biostimulant. Front. Plant Sci. 2022, 13, 2513. [Google Scholar] [CrossRef] [PubMed]
- Sut, S.; Ferrarese, I.; Shrestha, S.S.; Kumar, G.; Slaviero, A.; Sello, S.; Altissimo, A.; Pagni, L.; Gattesco, F.; Dall’Acqua, S. Comparison of biostimulant treatments in Acmella oleracea cultivation for alkylamides production. Plants 2020, 9, 818. [Google Scholar] [CrossRef]
- Taskin, E.; Misci, C.; Bandini, F.; Fiorini, A.; Pacini, N.; Obiero, C.; Ndaka Sila, D.; Tabaglio, V.; Puglisi, E. Smallholder Farmers’ Practices and African Indigenous Vegetables Affect Soil Microbial Biodiversity and Enzyme Activities in Lake Naivasha Basin, Kenya. Biology 2021, 10, 44. [Google Scholar] [CrossRef]
- Bandini, F.; Misci, C.; Taskin, E.; Cocconcelli, P.S.; Puglisi, E. Biopolymers modulate microbial communities in municipal organic waste digestion. FEMS Microbiol. Ecol. 2020, 96, fiaa183. [Google Scholar] [CrossRef]
- Vasileiadis, S.; Puglisi, E.; Trevisan, M.; Scheckel, K.G.; Langdon, K.A.; McLaughlin, M.J.; Lombi, E.; Donner, E. Changes in soil bacterial communities and diversity in response to long-term silver exposure. FEMS Microbiol. Ecol. 2015, 91, fiv114. [Google Scholar] [CrossRef]
- Berry, D.; ben Mahfoudh, K.; Wagner, M.; Loy, A. Barcoded primers used in multiplex amplicon pyrosequencing bias amplification. Appl. Env. Microbiol. 2011, 77, 7846–7849. [Google Scholar] [CrossRef]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 1–7. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Paulson, J.N.; Pop, M.; Bravo, H.C. Metastats: An improved statistical method for analysis of metagenomic data. Genome Biol. 2011, 12, 1–27. [Google Scholar] [CrossRef]
- White, J.R.; Nagarajan, N.; Pop, M. Statistical methods for detecting differentially abundant features in clinical metagenomic samples. PLoS Comput. Biol. 2009, 5, e1000352. [Google Scholar] [CrossRef]
- Tsouvaltzis, P.; Kasampalis, D.S.; Aktsoglou, D.-C.; Barbayiannis, N.; Siomos, A.S. Effect of reduced nitrogen and supplemented amino acids nutrient solution on the nutritional quality of baby green and red lettuce grown in a floating system. Agronomy 2020, 10, 922. [Google Scholar] [CrossRef]
- Castro Marín, I.; Loef, I.; Bartetzko, L.; Searle, I.; Coupland, G.; Stitt, M.; Osuna, D. Nitrate regulates floral induction in Arabidopsis, acting independently of light, gibberellin and autonomous pathways. Planta 2011, 233, 539–552. [Google Scholar] [CrossRef] [PubMed]
- Fageria, N.K.; Moreira, A. The role of mineral nutrition on root growth of crop plants. Adv. Agron. 2011, 110, 251–331. [Google Scholar]
- Blake, C.; Christensen, M.N.; Kovács, Á.T. Molecular aspects of plant growth promotion and protection by Bacillus subtilis. Mol. Plant-Microbe Interact. 2021, 34, 15–25. [Google Scholar] [CrossRef]
- Akinrinlola, R.J.; Yuen, G.Y.; Drijber, R.A.; Adesemoye, A.O. Evaluation of Bacillus strains for plant growth promotion and predictability of efficacy by in vitro physiological traits. Int. J. Microbiol. 2018, 2018. [Google Scholar] [CrossRef]
- Li, J.; Luo, C.; Zhang, D.; Cai, X.; Jiang, L.; Zhao, X.; Zhang, G. Diversity of the active phenanthrene degraders in PAH-polluted soil is shaped by ryegrass rhizosphere and root exudates. Soil Biol. Biochem. 2019, 128, 100–110. [Google Scholar] [CrossRef]
- Zhong, Z.; Huang, X.; Feng, D.; Xing, S.; Weng, B. Long-term effects of legume mulching on soil chemical properties and bacterial community composition and structure. Agric. Ecosyst. Environ. 2018, 268, 24–33. [Google Scholar] [CrossRef]
- Meidute, S.; Demoling, F.; Bååth, E. Antagonistic and synergistic effects of fungal and bacterial growth in soil after adding different carbon and nitrogen sources. Soil Biol. Biochem. 2008, 40, 2334–2343. [Google Scholar] [CrossRef]
- Li, J.-G.; Shen, M.-C.; Hou, J.-F.; Li, L.; Wu, J.-X.; Dong, Y.-H. Effect of different levels of nitrogen on rhizosphere bacterial community structure in intensive monoculture of greenhouse lettuce. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Mishra, V.; Rau, N.; Sharma, R.S. Increased iron-stress resilience of maize through inoculation of siderophore-producing Arthrobacter globiformis from mine. J. Basic Microbiol. 2016, 56, 719–735. [Google Scholar] [CrossRef] [PubMed]
- Lucini, L.; Colla, G.; Moreno, M.B.M.; Bernardo, L.; Cardarelli, M.; Terzi, V.; Bonini, P.; Rouphael, Y. Inoculation of Rhizoglomus irregulare or Trichoderma atroviride differentially modulates metabolite profiling of wheat root exudates. Phytochemistry 2019, 157, 158–167. [Google Scholar] [CrossRef]
- Luo, Y.; Zhou, M.; Zhao, Q.; Wang, F.; Gao, J.; Sheng, H.; An, L. Complete genome sequence of Sphingomonas sp. Cra20, a drought resistant and plant growth promoting rhizobacteria. Genomics 2020, 112, 3648–3657. [Google Scholar] [CrossRef]
- Groth, I.; Schumann, P.; Schütze, B.; Augsten, K.; Stackebrandt, E. Knoellia sinensis gen. nov., sp. nov. and Knoellia subterranea sp. nov., two novel actinobacteria isolated from a cave. Int. J. Syst. Evol. Microbiol. 2002, 52, 77–84. [Google Scholar] [CrossRef]
- Zhang, X.; Myrold, D.D.; Shi, L.; Kuzyakov, Y.; Dai, H.; Hoang, D.T.T.; Dippold, M.A.; Meng, X.; Song, X.; Li, Z.; et al. Resistance of microbial community and its functional sensitivity in the rhizosphere hotspots to drought. Soil Biol. Biochem. 2021, 161, 108360. [Google Scholar] [CrossRef]
- Biedendieck, R.; Knuuti, T.; Moore, S.J.; Jahn, D. The “beauty in the beast”—the multiple uses of Priestia megaterium in biotechnology. Appl. Microbiol. Biotechnol. 2021, 105, 5719–5737. [Google Scholar] [CrossRef]
- Jiang, H.; Feng, Y.; Zhao, F.; Lin, X. Characteristics and Complete Genome Analysis of Bacillus asahii OM18, a Bacterium in Relation to Soil Fertility in Alkaline Soils Under Long-Term Organic Manure Amendment. Curr. Microbiol. 2019, 76, 1512–1519. [Google Scholar] [CrossRef]
- Schlatter, D.C.; Yin, C.; Hulbert, S.; Paulitz, T.C. Core rhizosphere microbiomes of dryland wheat are influenced by location and land use history. Appl. Environ. Microbiol. 2020, 86, e02135-19. [Google Scholar] [CrossRef] [PubMed]
- Ozimek, E.; Hanaka, A. Mortierella species as the plant growth-promoting fungi present in the agricultural soils. Agriculture 2020, 11, 7. [Google Scholar] [CrossRef]
- Feng, J.; Huang, Z.; Zhang, Y.; Rui, W.; Lei, X.; Li, Z. Beneficial effects of the five isolates of Funneliformis mosseae on the tomato plants were not related to their evolutionary distances of SSU rDNA or PT1 sequences in the nutrition solution production. Plants 2021, 10, 1948. [Google Scholar] [CrossRef] [PubMed]
- Azcón, R.; Rodríguez, R.; Amora-Lazcano, E.; Ambrosano, E. Uptake and metabolism of nitrate in mycorrhizal plants as affected by water availability and N concentration in soil. Eur. J. Soil Sci. 2008, 59, 131–138. [Google Scholar] [CrossRef]
- Miransari, M. Arbuscular mycorrhizal fungi and nitrogen uptake. Arch. Microbiol. 2011, 193, 77–81. [Google Scholar] [CrossRef]
- Pozo, M.J.; Azcón-Aguilar, C. Unraveling mycorrhiza-induced resistance. Curr. Opin. Plant Biol. 2007, 10, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.A.; Liu, R.C.; Xiao, Z.Y.; Hashem, A.; Abd_Allah, E.F.; Alsayed, M.F.; Harsonowati, W.; Wu, Q.S. Symbiotic Fungi Alter the Acquisition of Phosphorus in Camellia oleifera through Regulating Root Architecture, Plant Phosphate Transporter Gene Expressions and Soil Phosphatase Activities. J. Fungi 2022, 8, 800. [Google Scholar] [CrossRef]
- Murillo-Roos, M.; Abdullah, H.S.M.; Debbar, M.; Ueberschaar, N.; Agler, M.T. Cross-feeding niches among commensal leaf bacteria are shaped by the interaction of strain-level diversity and resource availability. ISME J. 2022, 16, 2280–2289. [Google Scholar] [CrossRef]
- Lamont, J.R.; Wilkins, O.; Bywater-Ekegärd, M.; Smith, D.L. From yogurt to yield: Potential applications of lactic acid bacteria in plant production. Soil Biol. Biochem. 2017, 111, 1–9. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Content % |
|---|---|
| Alanine | 0.96 |
| Arginine | 0.40 |
| Asparagine (incl. Aspartic acid) | 1.07 |
| Hydroxyproline | 0.25 |
| Cysteine | 0.24 |
| Glutamic acid | 1.83 |
| Glycine | 0.55 |
| Histidine | 0.20 |
| Isoleucine | 0.46 |
| Leucine | 0.74 |
| Lysine | 0.77 |
| Methionine | 0.13 |
| Phenylalanine | 0.41 |
| Proline | 0.45 |
| Serine | 0.46 |
| Threonine | 0.41 |
| Tryptophan | 0.08 |
| Tyrosine | 0.29 |
| Valine | 0.62 |
| Fertilization Level | Shoot Fresh Biomass (g) | Root Fresh Biomass (g) |
|---|---|---|
| N100% | 173.57 (±14.23) ab | 17.96 (±2.31) |
| N70% | 159.82 (±3.75) b | 19.01 (±1.93) |
| N70% + N30% from eluate | 183.99 (±13.77) a | 17.47 (±1.72) |
| Fertilization Level | Shoot Fresh Biomass (g) | Root Fresh Biomass (g) |
|---|---|---|
| N100% | 107.69 (±7.26) | 22.53 (±4.31) |
| N70% | 99.24 (±8.51) | 17.30 (±2.62) |
| N70% + N30% from eluate | 120.65 (±14.22) | 20.74 (±4.85) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellotti, G.; Taskin, E.; Sello, S.; Sudiro, C.; Bortolaso, R.; Bandini, F.; Guerrieri, M.C.; Cocconcelli, P.S.; Vuolo, F.; Puglisi, E. LABs Fermentation Side-Product Positively Influences Rhizosphere and Plant Growth in Greenhouse Lettuce and Tomatoes. Land 2022, 11, 1544. https://doi.org/10.3390/land11091544
Bellotti G, Taskin E, Sello S, Sudiro C, Bortolaso R, Bandini F, Guerrieri MC, Cocconcelli PS, Vuolo F, Puglisi E. LABs Fermentation Side-Product Positively Influences Rhizosphere and Plant Growth in Greenhouse Lettuce and Tomatoes. Land. 2022; 11(9):1544. https://doi.org/10.3390/land11091544
Chicago/Turabian StyleBellotti, Gabriele, Eren Taskin, Simone Sello, Cristina Sudiro, Rossella Bortolaso, Francesca Bandini, Maria Chiara Guerrieri, Pier Sandro Cocconcelli, Francesco Vuolo, and Edoardo Puglisi. 2022. "LABs Fermentation Side-Product Positively Influences Rhizosphere and Plant Growth in Greenhouse Lettuce and Tomatoes" Land 11, no. 9: 1544. https://doi.org/10.3390/land11091544