1. Introduction
Mountainous rangelands provide a wide range of valuable ecosystem services [
1]. They are a source of high-quality forage for livestock [
2], especially for the transhumance livestock farming system [
3,
4]. Moreover, mountainous rangelands have a crucial role in the conservation of biodiversity and landscape preservation [
5] as well as in climate mitigation and water regulation [
6]. In addition, rangelands as cultural landscapes [
7] and protected natural reserves are attractive for recreational and touristic activities [
8]. For all these reasons, rangeland ecosystems have substantial direct or indirect impact on local economies in mountainous areas.
The management status of these ecosystems has changed rapidly in recent decades due to social and economic changes that have led to land use/land cover changes. On the one hand, the transhumant livestock activity has decreased [
9,
10], while on the other hand, tourist pressure has increased. All these changes have environmental and economic impacts on the ecosystem, its services, and local communities [
11].
Transhumance livestock grazing activities in mountainous areas have declined significantly in recent decades due to various socioeconomic reasons [
9,
10,
12,
13]. As a result, the extent and structure of mountain rangelands have changed [
14,
15], as livestock grazing has maintained them for centuries [
16]. Woody species encroachment due to transhumance livestock grazing abandonment is among the essential changes that has occurred in these rangelands’ lands [
17,
18,
19,
20] which negatively affects biodiversity [
21].
Since the 1970s, human pressure on mountain ecosystems has increased in several developed countries due to the development of ski resorts [
22] to cover the demands for recreational activities by the urban population. These activities generally have a positive economic impact on local communities in mountainous regions [
23], but ski resorts were significantly correlated with adverse changes in the rangelands. Their development includes using heavy machinery to construct runs and constructing and maintaining access roads and other infrastructure. Harsh conditions in high altitudes and the mechanical damage caused to plants by the construction and maintenance of ski-resort infrastructure retards the recovery of the vegetation cover [
24]. As a result, these infrastructures have been reported to cause a reduction in species richness and plant cover in the rangelands [
25], increasing the risk of soil erosion as well as changing soil properties [
26,
27], which in turn may have negative impacts on ecosystem functioning and stability [
28]. On the other hand, Allegrezza and coworkers [
16] did not find any differences in floristic diversity among undisturbed alpine grassland and grasslands with ski runs covered with natural and artificial snow. It seems that differences in altitude, slope, different management practices applied, and time passed after the ski resort was built are among the factors affecting the floristic diversity in these ecosystems [
29].
In some mountainous areas in the Mediterranean region, ski resorts coexist with extensive pasture-based livestock farming. However, there are a limited number of studies investigating both the effects of ski resorts and livestock grazing on plant communities and ecosystem function [
30]. In this regard, Goñi and Gúzman [
31] proposed that livestock grazing can prevent reductions in plant diversity caused due to ski resorts. A useful tool for assessing the impact of different management regimes on the rangeland ecosystem’s function is the indices of rangeland health [
32,
33,
34].
Recently, the goal of public governance and local communities is the sustainable development of rural areas, which includes three axes: the economic viability of local communities, social cohesion, and environmental sustainability. These three axes collide in some cases but are recognized as having the same weight and importance in ensuring sustainable development [
35].
In this respect, the coexistence of tourism with pasture-based livestock is desirable and essential for the sustainability of the less favorable mountainous areas [
36]. Moreover, tourist activities related to skiing and transhumance livestock are two non-rivalrous activities in terms of time, since skiing is carried out during the winter months, while livestock farming is carried out from spring to autumn. The question that arises is to what extent the creation and operation of ski centers conflict with transhumance livestock spatially and to what extent the use of rangelands by both activities ultimately leads to their degradation.
In as much as, to the best of our knowledge, there are no similar studies on the combined effect of pastoralism and touristic activities on rangeland ecosystems, we conducted the current study to gain insights on the impact of these activities on both floristic diversity and ecosystem function. The outcome of our analyses, as far as the interaction of pastoralism and touristic activities is regarded and their effects on the ecosystem as a whole, can provide a basis for designing and establishing strategies for the sustainable development of mountainous areas. This research was conducted in a mountainous area of Southern Greece, traditionally used by transhumance, while a ski resort has been present since the 2000s. We assessed the impact of the ski-resort infrastructure and livestock grazing on the rangeland ecosystems; specifically, as far as (1) floristic composition and diversity and (2) rangeland health indices related to ecosystem stability and function are concerned. We tested the hypothesis that ski resorts and transhumance livestock farming coexistence in Mediterranean mountainous areas results in severe degradation of the ecosystem.
3. Results
Significant differences (
p ≤ 0.05) between sites were recorded for plant cover, the functional groups legumes and forbs, the Simpson index, and the Berger–Parker dominance index (
Table 2). Additionally, significant differences (
p ≤ 0.05) were recorded among the years for the functional groups grasses, legumes, and forbs, the species richness, the Simpson, Shannon, Margalef indices, and the Berger–Parker dominance index. The interaction of site and year was significant (
p ≤ 0.05) for forbs, the species richness, the Simpson, Shannon, and Margalef diversity indices, and the Berger–Parker dominance index (
Table 2).
The plant cover (across years) was higher in Sarantapicho rangeland. Functional group composition was differentiated between sites. The percentage of legumes was higher in Sarantapicho, while more forbs were presented in Ano Trikala. There was a slight trend of higher floristic diversity in Sarantapicho, without significant differences. Only the Simpson index was significantly higher, while the Berger–Parker dominance index was significantly lower in Sarantapicho (
Table 3).
The percentage of forbs progressively increased, and it was significantly higher in 2019 compared to those recorded in 2014 and 2015 (
Table 4). An opposite trend was recorded for grasses and woody species but without producing significant results. The percentage of legumes was significantly lower in 2014. Floristic diversity indices (species richness, Simpson, Shannon, Margalef) were significantly higher in 2015 compared to 2014, while in 2019, these indices had intermediate values without significantly differentiating from the other years. The Berger–Parker index of dominance followed an opposite trend.
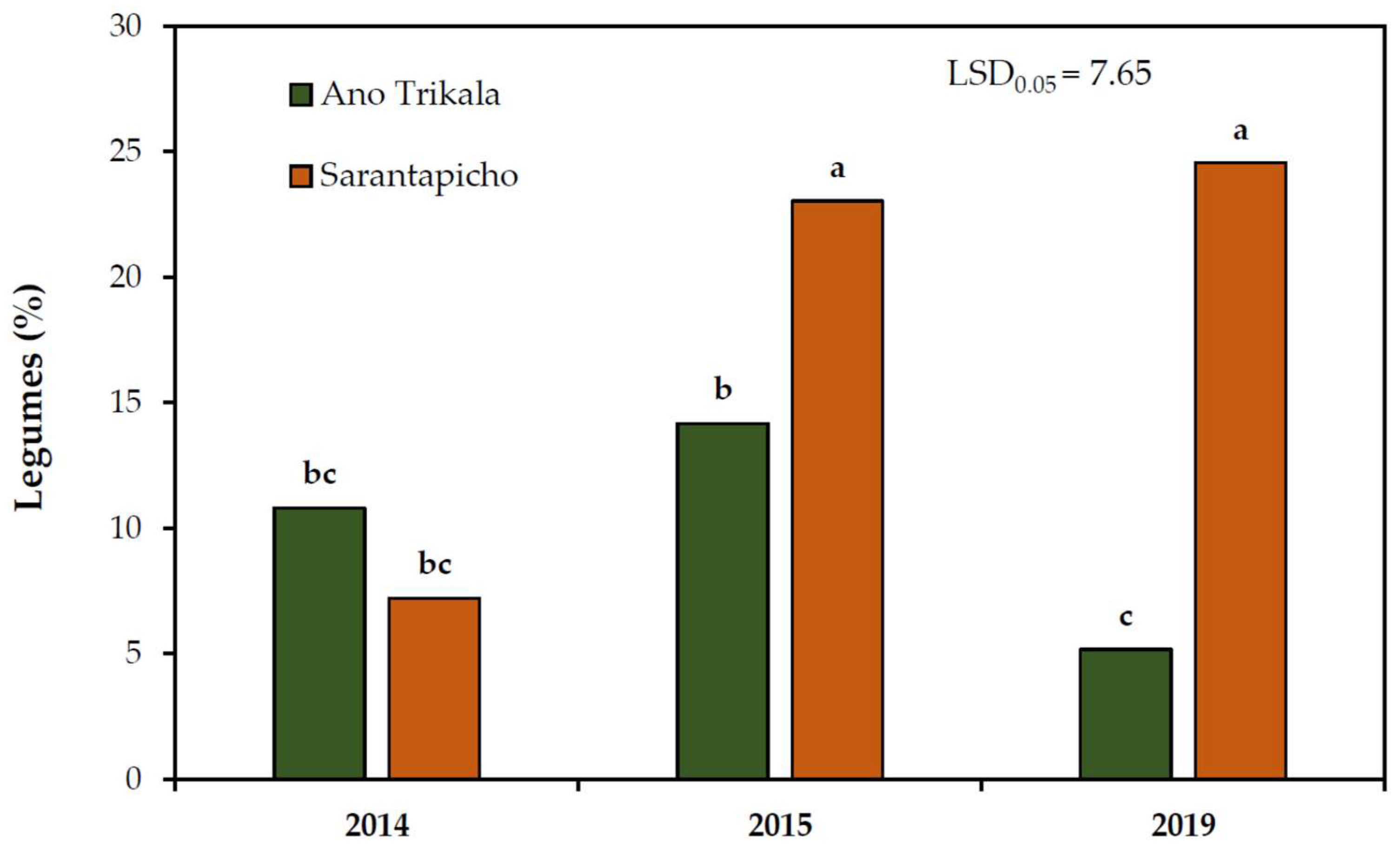
The percentage of legumes in Sarantapicho was significantly lower in 2014 compared to 2015 and 2019, while it was higher in 2015 compared to 2019 in Ano Trikala. The percentages of legumes were significantly higher in Sarantapicho than in Ano Trikala in 2015 and 2019, while no significant differences (
p > 0.05) were detected between sites in 2014 (
Figure 2).
The species richness, Simpson, Shannon, and Margalef indices in Ano Trikala were significantly lower in 2014 compared to 2015 and 2019, while in Sarantapicho, no differences were recorded among years. The values of the Berger–Parker dominance index followed the opposite trend. Those indices were significantly (higher in Sarantapicho than in Ano Trikala only in 2014, while no significant differences (
p > 0.05) were detected among sites in 2015 and 2019 (
Table 5). Berger–Parker dominance index was significantly lower in Sarantapicho than in Ano Trikala in 2014 and 2015.
Significant differences (
p ≤ 0.05) between sites were recorded for all the indices of landscape composition, function, stability, and forage value index (
Table 6). Additionally, significant differences (
p ≤ 0.05) for the landscape function index and the total score of indices were recorded over the years. The interaction of site and year was significant (
p ≤ 0.05) for the landscape function index and the forage value index (
Table 5).
All the indices of landscape composition, function, stability, and forage value index were significantly (
p ≤ 0.05) higher in Sarantapicho compared to Ano Trikala (
Table 7).
The landscape function index and the total score of indices were significantly lower in 2014 compared to those recorded in 2015 and 2019 (
Table 8).
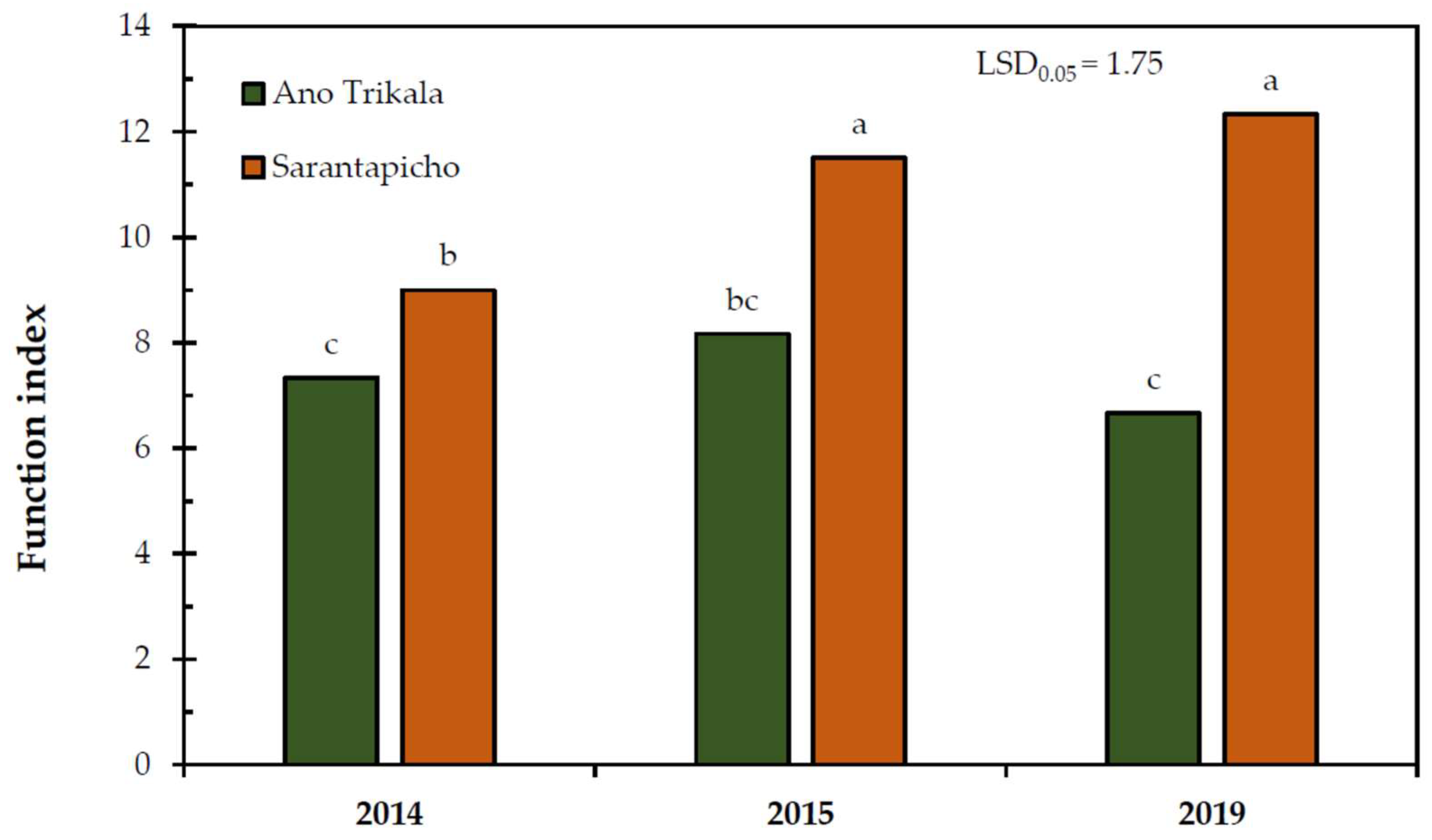
The landscape function index in Sarantapicho was significantly lower in 2014 compared to 2015 and 2019, while in Ano Trikala, no significant differences (
p > 0.05) were detected among years. The landscape function index was significantly higher in Sarantapicho than in Ano Trikala in all the years (
Figure 3).
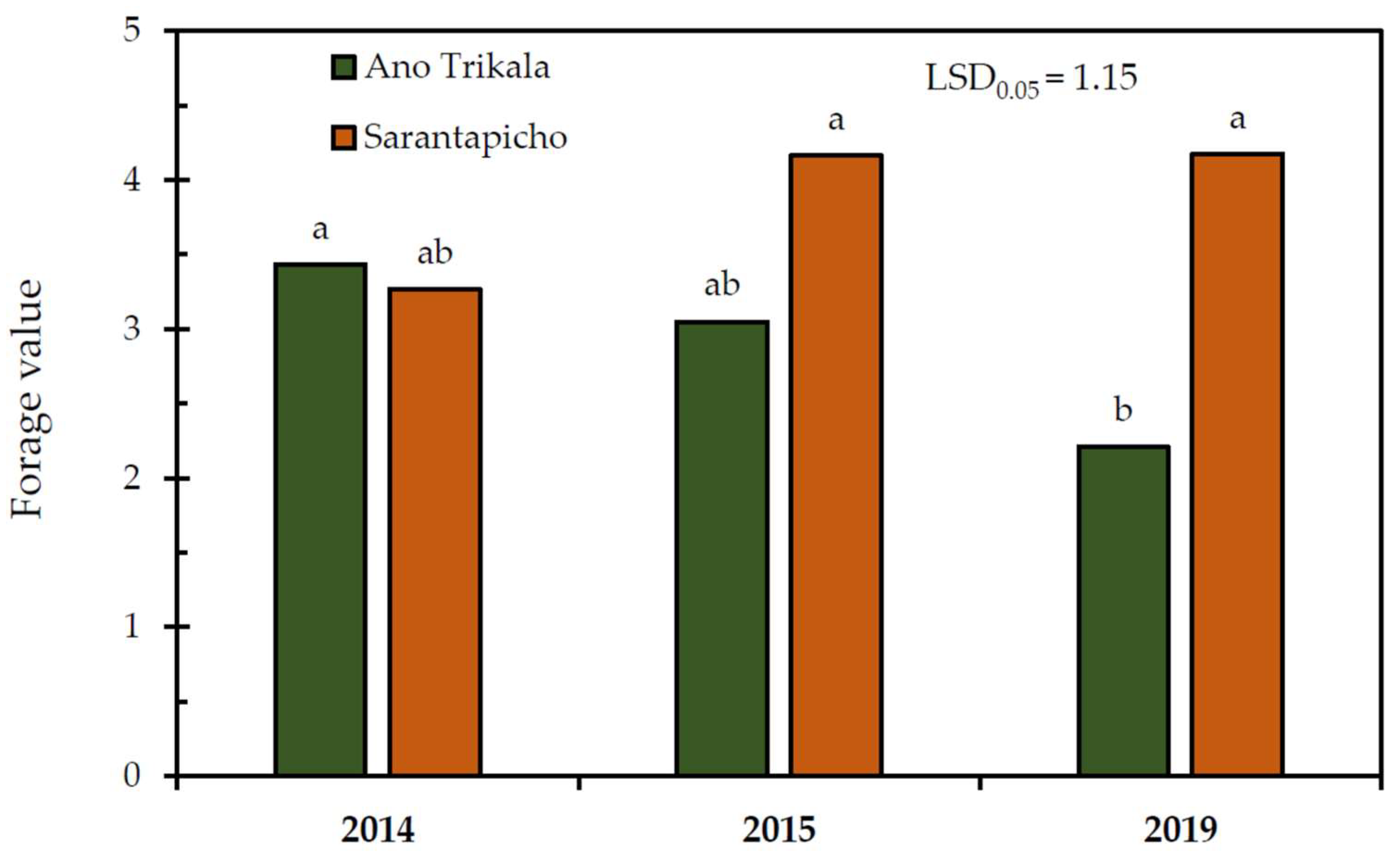
The forage value index in Ano Trikala was significantly lower in 2019 compared to 2014, while in Sarantapicho no significant differences (
p > 0.05) were detected among years. The forage value index was significantly higher in Sarantapicho than in Ano Trikala only in 2019, while no significant differences were detected between sites in 2014 and 2019 (
Figure 4).
4. Discussion
Ski resorts and transhumance livestock farming are activities that often coexist in mountainous areas, which may strongly affect the structure and function of rangeland ecosystems. Nevertheless, our data and analyses cannot support the negative impact hypothesis for a representative mountain of South Mediterranean area.
The current study indicates that the development of the ski resort in Ano Trikala had a minimal negative impact on rangelands’ plant cover. Ski-resort infrastructure and the increased number of visitors usually cause considerable trampling and other disturbances, leading to a decrease in plant cover [
26,
30,
50]. In such sites, the soils are compacted, and as a result, plant growth is limited, infiltration rates and water-storage capacity are reduced, and soil erosion risk is high [
51]. However, this was not the case in the ski resort in Ano Trikala, possibly because of the low-intensity touristic activities in the specific site. Transhumance livestock grazing has been reported to positively impact maintaining plant cover in high values [
38]. This result is in accordance with those of Goñi and Gúzman [
31], who also reported a minimum increase in bare soil in a grazed ski resort area in Spain. A slight reduction in plant cover was detected in Ano Trikala compared to Sarantapicho; plant cover was generally high in both mountainous rangelands, remaining stable over the years.
Regarding the vegetation composition, the percentage of legumes was lower in the mountainous rangeland close to the ski resort of Ano Trikala, while more forbs were present compared to the rangeland located away from the ski resort. These results are in accordance with previous findings in Spain [
30]. Legumes have deeper root systems in general than grasses and forbs. The presence of ice, the reduced soil microporosity that causes poor aeration, and the trampling caused by visitors had increased negative effects on the legume roots compared to those of the other plant functional groups [
30,
52]. Thus, legumes were significantly less in the rangeland close to the ski resort. The negative impact of the ski resort on legumes is further confirmed by their significant reduction from 2014 to 2019 when the opposite trend was recorded for the rangelands located away from the ski resort. Forbs (across sites) were significantly more in 2019 compared to those recorded in 2014 and 2015, while an opposite trend was recorded for grasses. The differences in the contribution of these plant groups are probably related to grazing and the relative higher preference of sheep for legumes than for forbs when they are available [
53], as well as to the differences in climatic over the years [
9,
38,
54,
55].
The ski-resort development in Ano Trikala generally had minimal effects on floristic diversity. However, it has been noted that the Simpson index was significantly lower, while the Berger–Parker dominance index was significantly higher compared to those recorded in the mountainous rangeland located away from the ski resort, indicating a slight decrease in floristic diversity at the expense of the increasing abundance of the dominant species near the ski resort. Previous studies [
24,
56] have reported reduced species richness and diversity close to ski resorts. However, there is evidence that livestock grazing can counterbalance the adverse effects of ski vehicles and visitors through micro-depressions due to trampling and exozoochoria, which can maintain diversity [
22]. Thus, Goñi and Gúzman [
31] recorded higher plant diversity in grazed ski runs than in non-grazed ones. Barrantes and coworkers [
30] found that floristic diversity in grazed skiing areas increased between 1972 and 2005. The present study confirms this result as floristic diversity was also recorded to increase in 2019 compared to 2014 in rangelands close to the Ano Trikala ski resort, while in Sarantapicho biodiversity indices did not change among years. Floristic diversity (across sites) was higher in 2015 compared to 2014 and 2019, probably because of the annual fluctuations in rainfall and temperature [
30].
The landscape was negatively affected by the ski resort and related activities. All the indices of landscape composition, function, and stability were significantly higher in the rangeland located in Sarantapicho, away from the ski resort. The increased soil erosion, reduced plant cover and herbage production, and the decreased legume percentage recorded in rangelands close to the ski resort in Ano Trikala constitute the main reasons for this result. Increased risk of soil erosion close to ski resorts has been reported in previous studies [
26,
57]. Moreover, significant decreases in productivity in rangelands in proximity to ski resorts have been reported by Gartzia and coworkers [
58]. It has to be noted that the landscape function index remained low in this site during the years, while it progressively increased in the rangeland located away from the ski resort.
Forage value was significantly lower close to the ski resort. This result is related to the significantly lower percentage of legumes, which have high forage value, and the higher percentage of less palatable forbs. The lower forage value implies this area’s decreased grazing capacity and a need for a reduced stocking rate. It is in accordance with the results of previous studies [
30,
31] that also reported lower pasture quality close to ski resorts. The result of the present study is further confirmed by the fact that forage value remained stable from 2014 to 2019, when it increased in the rangelands located away from the ski resort.
5. Conclusions
Proximity to ski resorts did not reduce plant cover seriously, while it had a minor impact on species richness and floristic diversity. These results suggest that livestock grazing can counterbalance the effects of ski resorts and related activities on plant cover and floristic diversity. On the other hand, the floristic composition was modified in the ski center area. These differences in species composition are reflected in forage value, which is reduced in the ski-resort area. Besides the relatively limited effects on the vegetation community, the ski resort had a significant negative impact on landscape composition, function, and stability.
Transhumance livestock grazing should be used as a management tool in ski-resort areas as it benefits floristic diversity. Low-intensity touristic activities have relatively limited effects on such areas, but some restoration activities may be needed in the future. Furthermore, the two activities could be complementary for the employment of the residents of local communities, as tourism is an activity most preferred by young people, while livestock farming is mainly employed by elderly people. Finally, monitoring these areas at both levels of landscape and vegetation is necessary for managers to make the appropriate decisions at the right time. In this respect, the landscape indices that were used in the present study could be a useful tool for future monitoring and management decisions.

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}