
Impacts of Melatonin on Functionalities of Constructed Wetlands for Wastewater Treatment
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup and Synthetic Wastewater
2.2. Water Samplings and Analysis
2.3. Physiological Performance of Phragmites australis
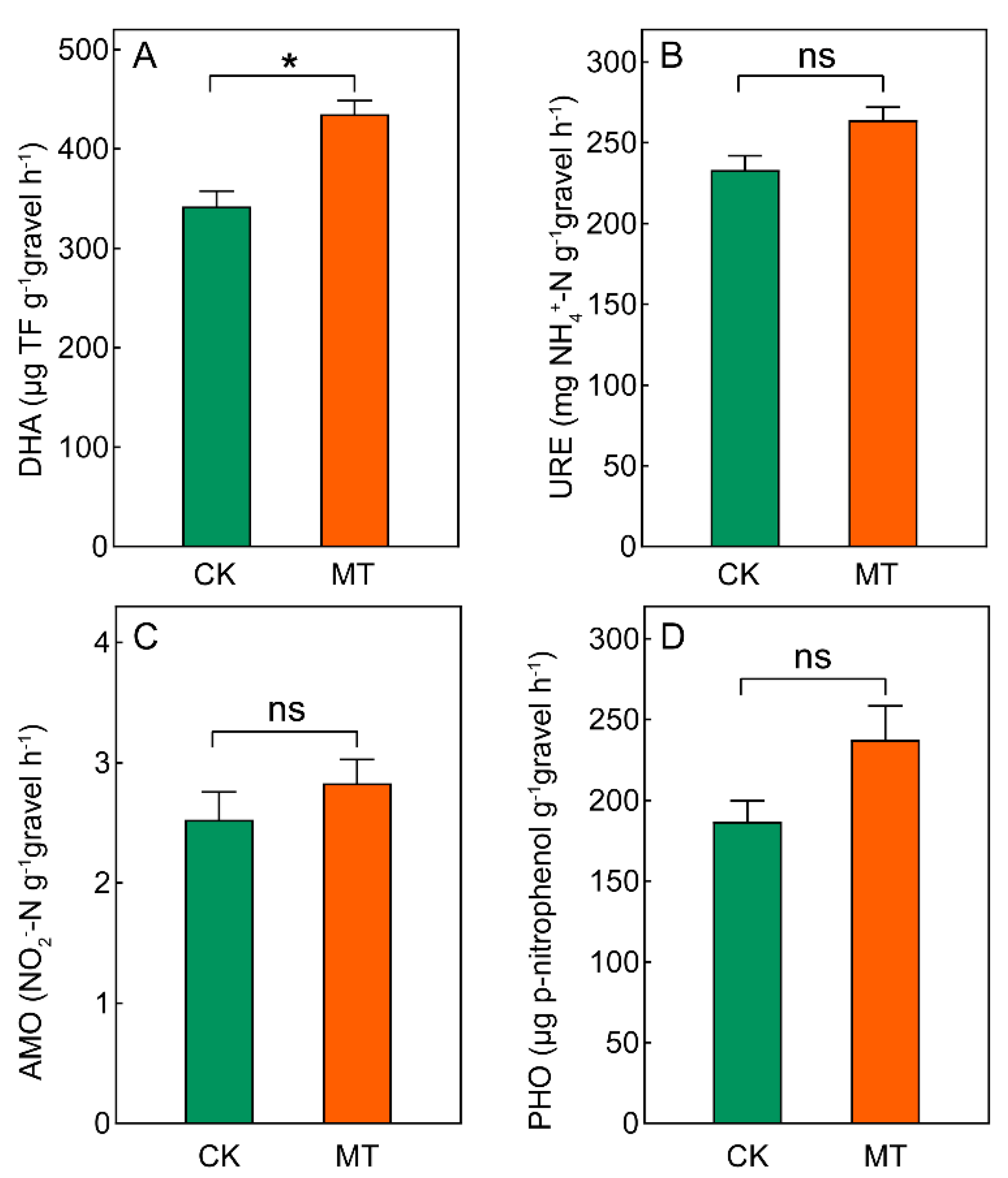
2.4. Microbial Enzyme Activities
2.5. DNA Extraction and 16S/ITS rRNA Gene Sequencing
2.6. Statistical Analysis
3. Results
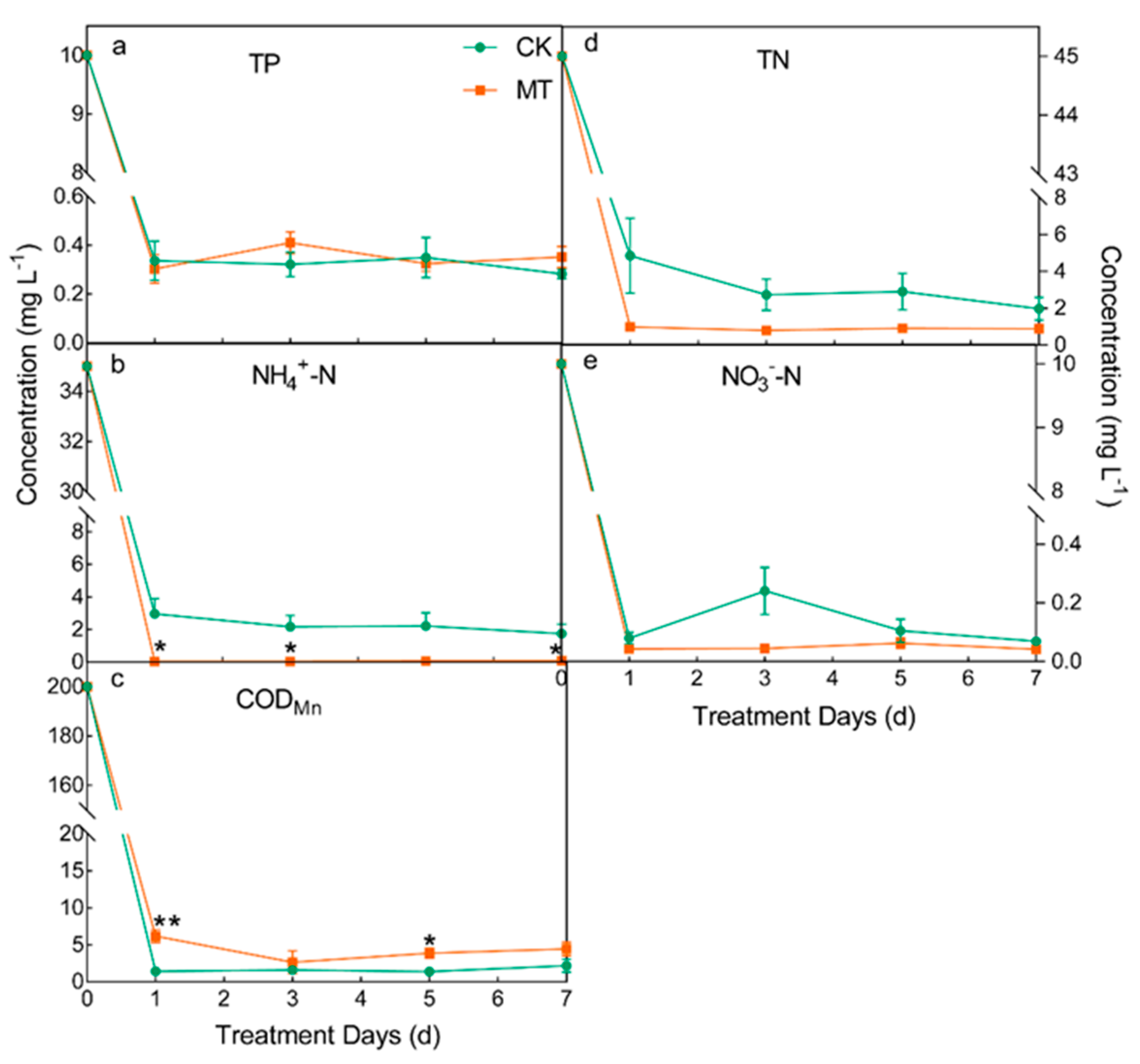
3.1. Treatment Performance of CWs as Affected by Melatonin
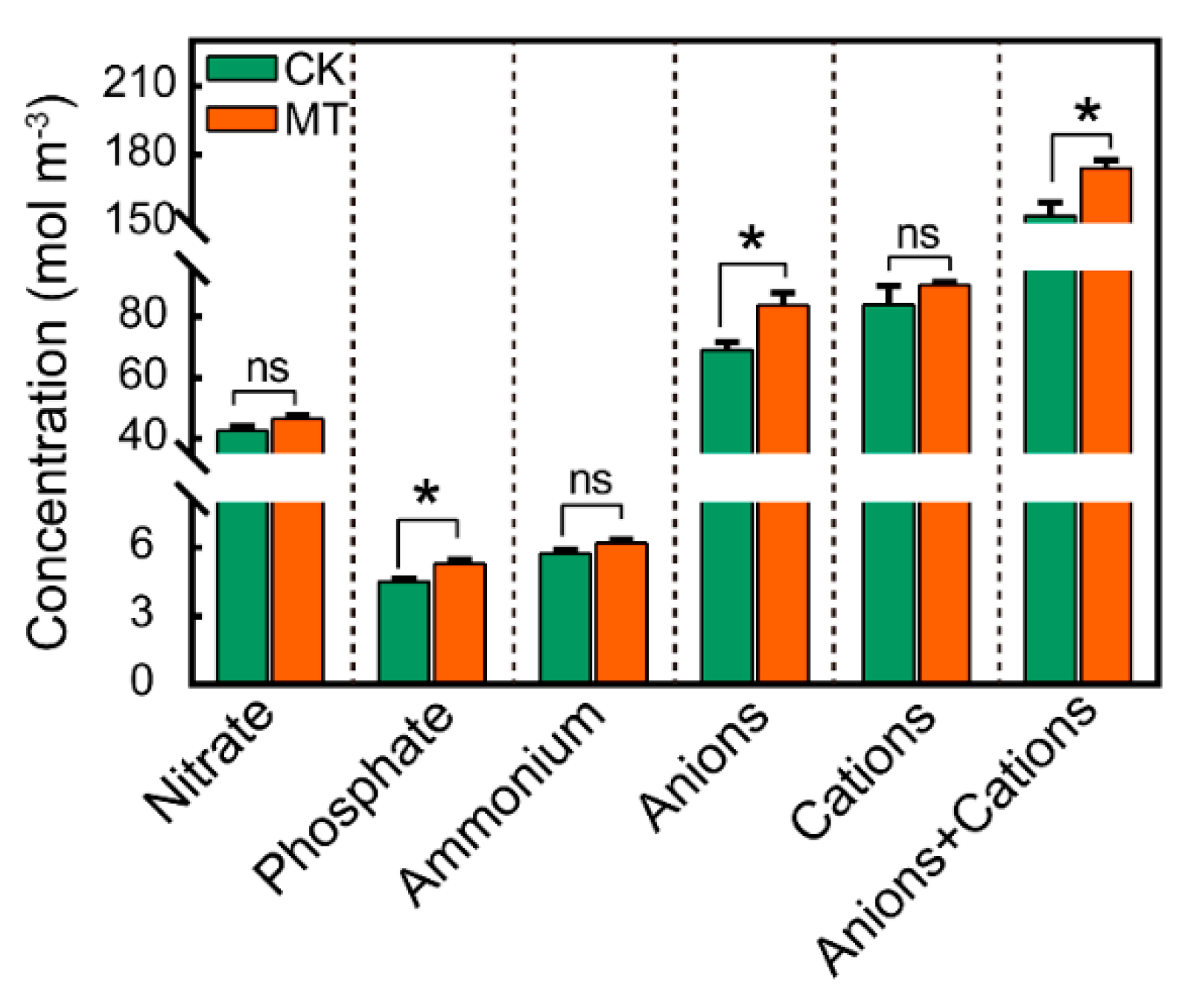
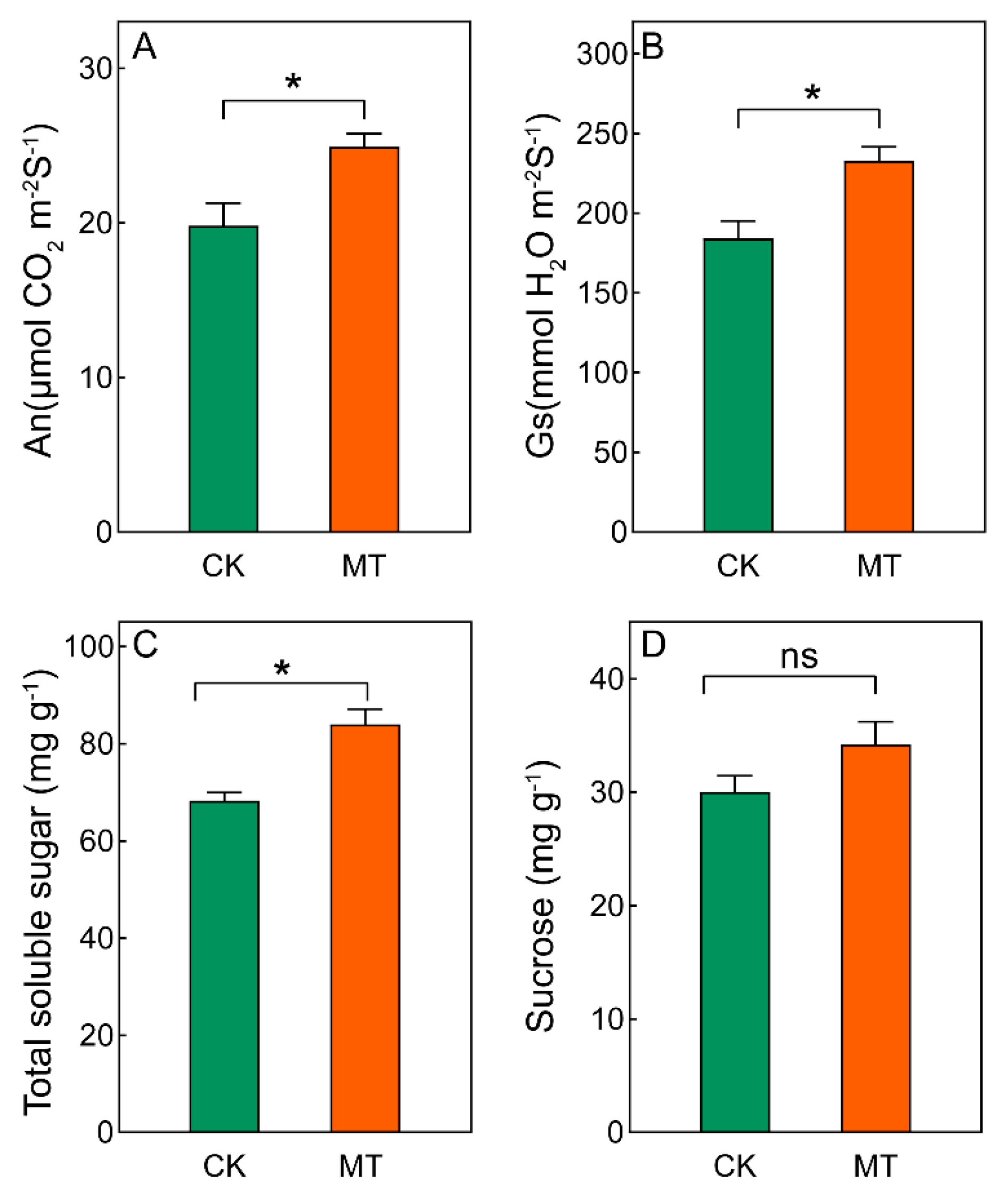
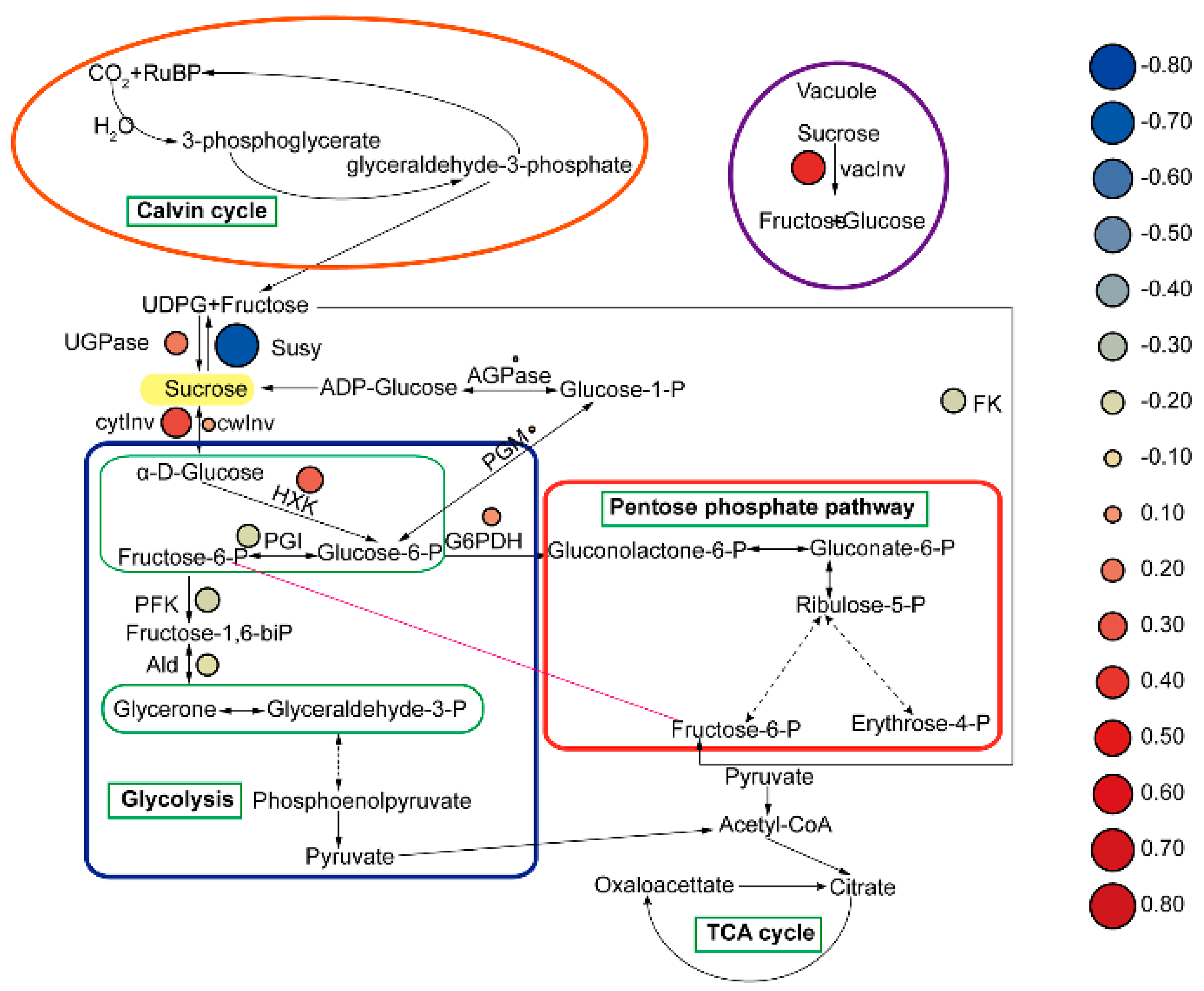
3.2. Physiological Response of Phragmites australis Aerial Portion as Affected by Melatonin
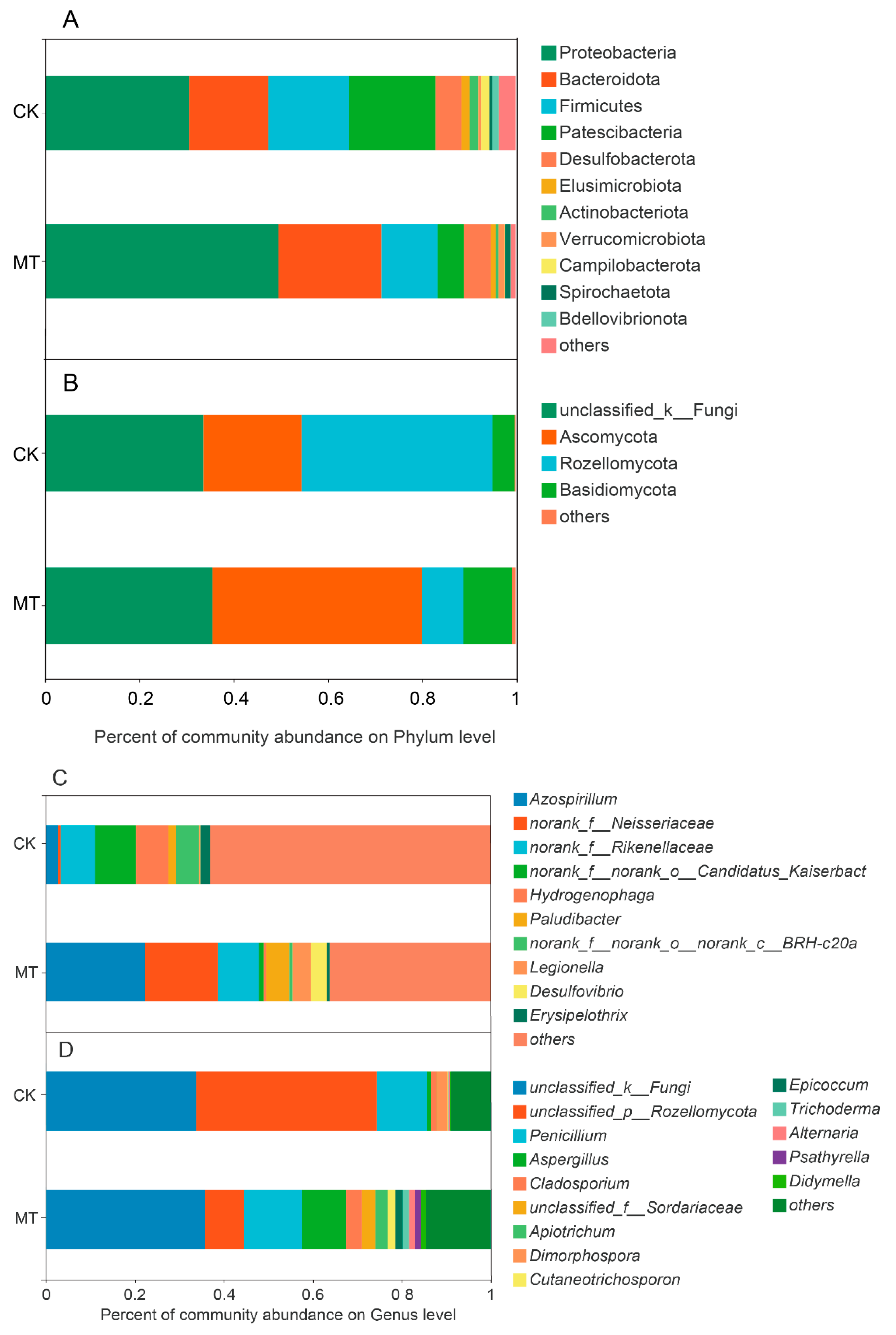
3.3. Microbiota of CWs as Affected by Melatonin
3.3.1. Microbial Community Structure
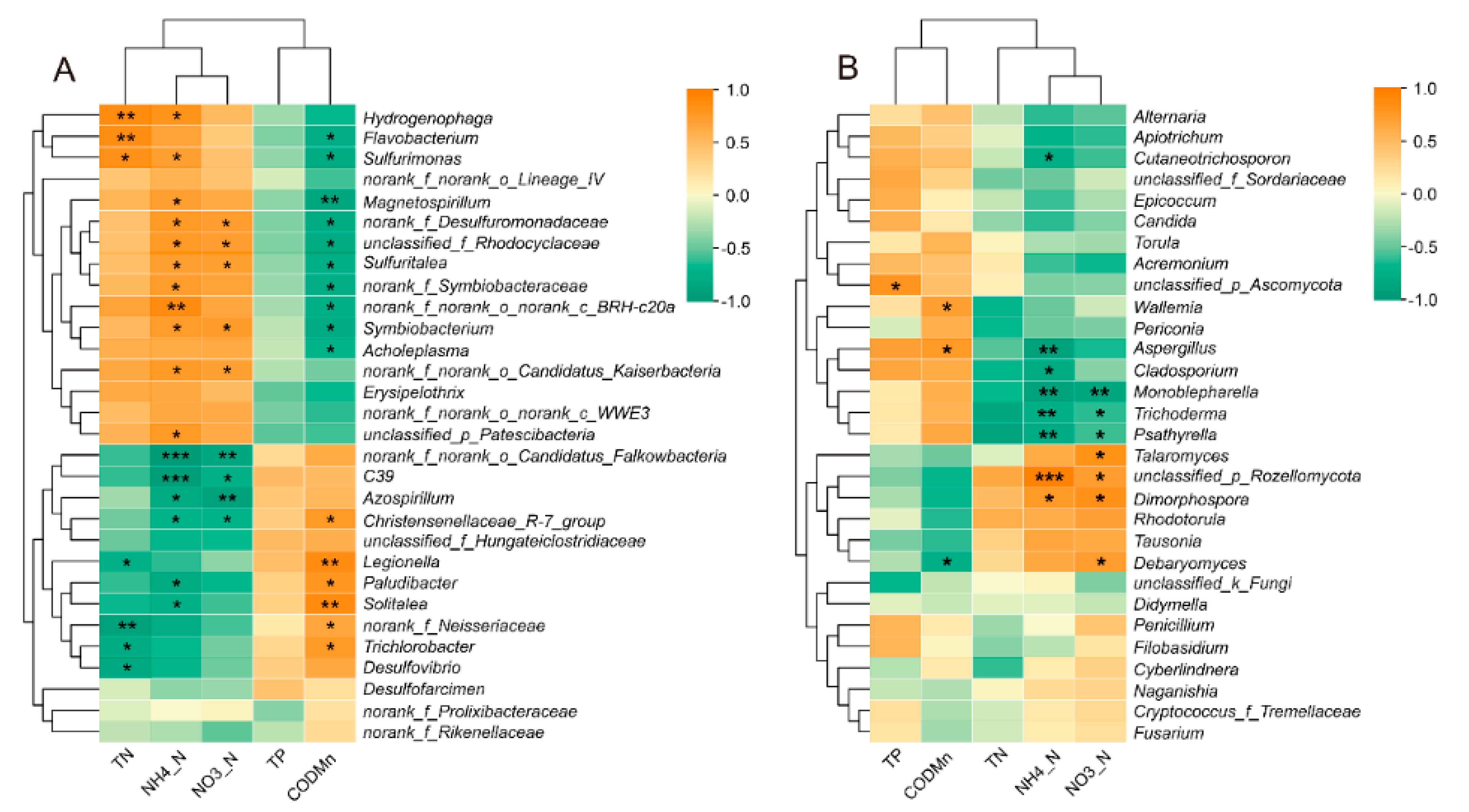
3.3.2. Correlation between Contaminants Concentration and Microbial Communities
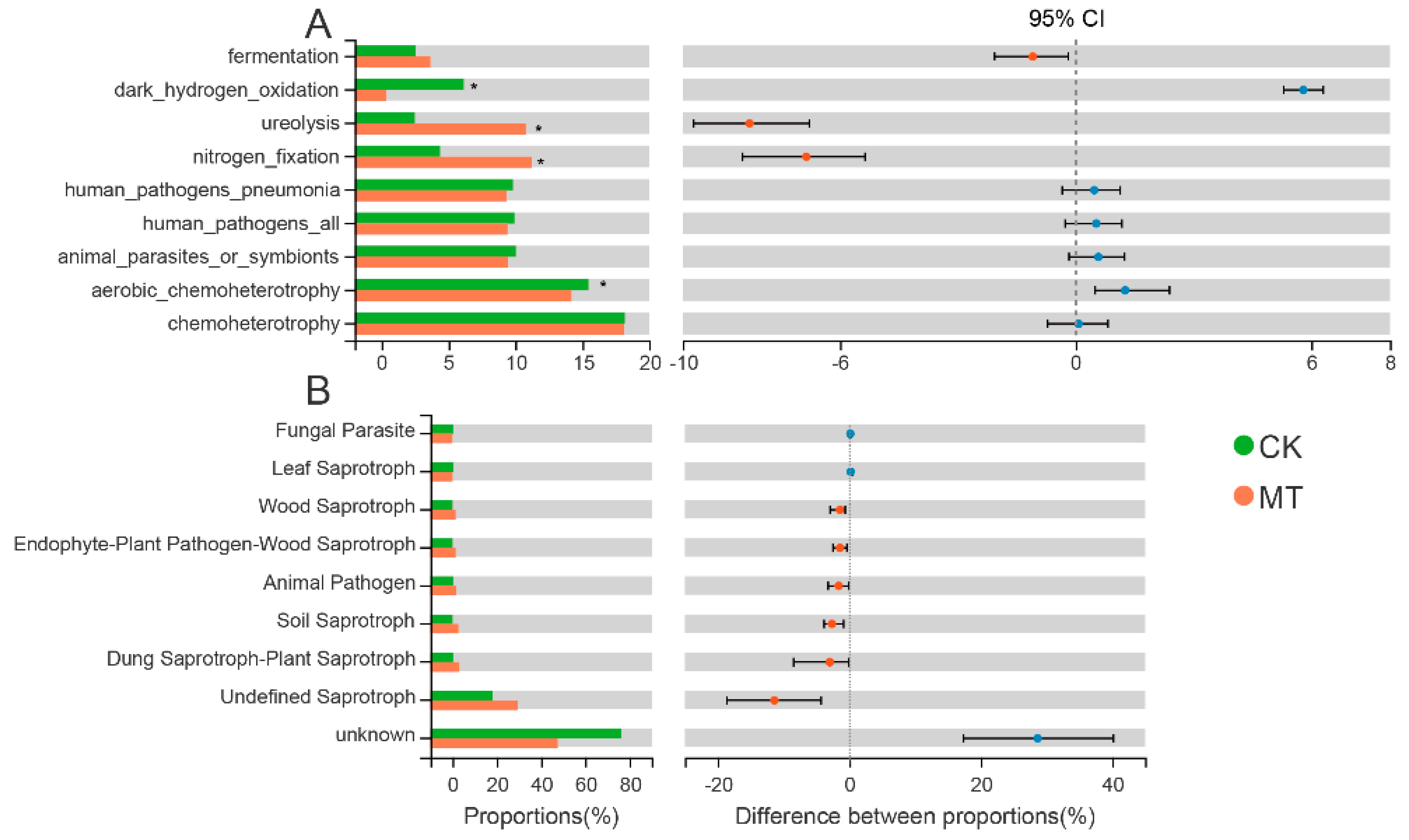
3.3.3. Prediction of Functional Composition in the Microbial Community
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kivaisi, A.K. The potential for constructed wetlands for wastewater treatment and reuse in developing countries: A review. Ecol. Eng. 2001, 16, 545–560. [Google Scholar] [CrossRef]
- Sundaravadivel, M.; Vigneswaran, S. Constructed wetlands for wastewater treatment. Crit. Rev. Environ. Sci. Technol. 2001, 31, 351–409. [Google Scholar] [CrossRef]
- Brix, H. Macrophyte-Mediated Oxygen Transfer in Wetlands: Transport Mechanisms and Rates. In Constructed Wetlands for Water Quality Improvement; CRC Press: Boca Raton, FL, USA, 1993; pp. 391–398. [Google Scholar]
- Wang, Q.; Hu, Y.; Xie, H.; Yang, Z. Constructed wetlands: A review on the role of radial oxygen loss in the rhizosphere by macrophytes. Water 2018, 10, 678. [Google Scholar] [CrossRef] [Green Version]
- Fu, G.; Wu, J.; Han, J.; Zhao, L.; Chan, G.; Leong, K. Effects of substrate type on denitrification efficiency and microbial community structure in constructed wetlands. Bioresour. Technol. 2020, 307, 123222. [Google Scholar] [CrossRef] [PubMed]
- Khalilzadeh, R.; Pirzad, A.; Sepehr, E.; Khan, S.; Anwar, S. Efficiency of Phragmites australis under different times of wastewater irrigation in the soil–plant–water system. Int. J. Environ. Sci. Technol. 2022, 19, 1957–1976. [Google Scholar] [CrossRef]
- Meng, P.; Pei, H.; Hu, W.; Shao, Y.; Li, Z. How to increase microbial degradation in constructed wetlands: Influencing factors and improvement measures. Bioresour. Technol. 2014, 157, 316–326. [Google Scholar] [CrossRef]
- Truu, M.; Juhanson, J.; Truu, J. Microbial biomass, activity and community composition in constructed wetlands. Sci. Total Environ. 2009, 407, 3958–3971. [Google Scholar] [CrossRef]
- Hu, B.; Hu, S.; Vymazal, J.; Chen, Z. Application of arbuscular mycorrhizal fungi for pharmaceuticals and personal care productions removal in constructed wetlands with different substrate. J. Clean. Prod. 2022, 339, 130760. [Google Scholar] [CrossRef]
- Tan, D.X.; Hardeland, R.; Manchester, L.C.; Korkmaz, A.; Ma, S.; Rosales-Corral, S.; Reiter, R.J. Functional roles of melatonin in plants, and perspectives in nutritional and agricultural science. J. Exp. Bot. 2012, 63, 577–597. [Google Scholar] [CrossRef]
- Huang, J.; Wang, S.H.; Yan, L.; Zhong, Q.S. Plant photosynthesis and its influence on removal efficiencies in constructed wetlands. Ecol. Eng. 2010, 36, 1037–1043. [Google Scholar] [CrossRef]
- Luo, W.; Wang, S.; Huang, J.; Yan, L.; Huang, J. Impact of photosynthesis and transpiration on nitrogen removal in constructed wetlands. Front. Environ. Sci. Eng. China 2007, 1, 316–319. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Role of melatonin to enhance phytoremediation capacity. Appl. Sci. 2019, 9, 5293. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Xiong, X.; Huang, Z.; Sun, L.; Ma, J.; Cai, S.; Yu, F.; Zhong, W.; Chen, S.; Li, X. Exogenous melatonin improves lead tolerance of bermudagrass through modulation of the antioxidant defense system. Int. J. Phytoremediation 2018, 20, 1408–1417. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Wang, P.; Chen, X.; Peng, Y.; Cai, B.; Song, J.; Yin, G.; Jia, S.; Zhang, H. Melatonin alleviates cadmium toxicity and abiotic stress by promoting glandular trichome development and antioxidant capacity in Nicotiana tabacum. Ecotoxicol. Environ. Saf. 2022, 236, 113437. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Li, A.; Yu, H.; Li, W.; Liang, C.; Guo, S.; Zhang, R.; Chu, C. Melatonin regulates root architecture by modulating auxin response in rice. Front. Plant Sci. 2017, 8, 134. [Google Scholar] [CrossRef] [Green Version]
- Madigan, A.P.; Egidi, E.; Bedon, F.; Franks, A.E.; Plummer, K.M. Bacterial and fungal communities are differentially modified by melatonin in agricultural soils under abiotic stress. Front. Microbiol. 2019, 10, 02616. [Google Scholar] [CrossRef] [Green Version]
- Ye, F.; Jiang, M.; Zhang, P.; Liu, L.; Liu, S.; Zhao, C.; Li, X. Exogenous Melatonin Reprograms the Rhizosphere Microbial Community to Modulate the Responses of Barley to Drought Stress. Int. J. Mol. Sci. 2022, 23, 9665. [Google Scholar] [CrossRef]
- Fan, H.X.; Guo, R.X.; Xin, G.Q.; Li, F.Q. Effects of exogenous melatonin on growth and physiological characteristics of reedings under salt stress. J. Agric. Sci. Technol. 2019, 21, 51–58. [Google Scholar]
- Wiessner, A.; Kappelmeyer, U.; Kuschk, P.; Kastner, M. Influence of the redox condition dynamics on the removal efficiency of a laboratory-scale constructed wetland. Water Res. 2005, 39, 248–256. [Google Scholar] [CrossRef]
- Administration, T.P. Water and Wastewater Monitoring and Analysis Methods, 4th ed.; China Environmental Science Press: Beijing, China, 2002. [Google Scholar]
- Jammer, A.; Gasperl, A.; Luschin-Ebengreuth, N.; Heyneke, E.; Chu, H.; Cantero-Navarro, E.; Grosskinsky, D.K.; Albacete, A.A.; Stabentheiner, E.; Franzaring, J. Simple and robust determination of the activity signature of key carbohydrate metabolism enzymes for physiological phenotyping in model and crop plants. J. Exp. Bot. 2015, 66, 5531–5542. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Liu, X.; Yang, X.; Guo, F.; Su, X.; Chen, Y. Acute and chronic responses of macrophyte and microorganisms in constructed wetlands to cerium dioxide nanoparticles: Implications for wastewater treatment. Chem. Eng. J. 2018, 348, 35–45. [Google Scholar] [CrossRef]
- Klose, S.; Tabatabai, M.A. Urease activity of microbial biomass in soils as affected by cropping systems. Biol. Fertil. Soils 2000, 31, 191–199. [Google Scholar] [CrossRef]
- Zheng, X.; Chen, Y.; Wu, R. Long-term effects of titanium dioxide nanoparticles on nitrogen and phosphorus removal from wastewater and bacterial community shift in activated sludge. Environ. Sci. Technol. 2011, 45, 7284–7290. [Google Scholar] [CrossRef]
- Wang, Z.; Li, S.; Liu, S.; Wang, F.; Kong, L.; Li, X.; Brestic, M. Effects of Elevated Atmospheric CO2 Concentration on Phragmites australis and Wastewater Treatment Efficiency in Constructed Wetlands. Water 2021, 13, 2500. [Google Scholar] [CrossRef]
- Saharimoghaddam, N.; Massoudinejad, M.; Ghaderpoori, M. Removal of pollutants (COD, TSS, and NO3−) from textile effluent using Gambusia fish and Phragmites australis in constructed wetlands. Environ. Geochem. Health 2019, 41, 1433–1444. [Google Scholar] [CrossRef]
- Huang, J.; Cai, W.; Zhong, Q.; Wang, S. Influence of temperature on micro-environment, plant eco-physiology and nitrogen removal effect in subsurface flow constructed wetland. Ecol. Eng. 2013, 60, 242–248. [Google Scholar] [CrossRef]
- Chen, Y.E.; Mao, J.J.; Sun, L.Q.; Huang, B.; Ding, C.B.; Gu, Y.; Liao, J.; Hu, C.; Zhang, Z.; Yuan, S. Exogenous melatonin enhances salt stress tolerance in maize seedlings by improving antioxidant and photosynthetic capacity. Physiol. Plant. 2018, 164, 349–363. [Google Scholar] [CrossRef]
- Zuo, Z.; Sun, L.; Wang, T.; Miao, P.; Zhu, X.; Liu, S.; Song, F.; Mao, H.; Li, X. Melatonin improves the photosynthetic carbon assimilation and antioxidant capacity in wheat exposed to nano-ZnO stress. Molecules 2017, 22, 1727. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Jing, T.; Wang, Y.; Ai, X.; Bi, H. Melatonin Delays Leaf Senescence and Improves Cucumber Yield by Modulating Chlorophyll Degradation and Photoinhibition of PSII and PSI. Environ. Exp. Bot. 2022, 200, 104915. [Google Scholar] [CrossRef]
- Liu, F.; Sun, L.; Wan, J.; Shen, L.; Yu, Y.; Hu, L.; Zhou, Y. Performance of different macrophytes in the decontamination of and electricity generation from swine wastewater via an integrated constructed wetland-microbial fuel cell process. J. Environ. Sci. 2020, 89, 252–263. [Google Scholar] [CrossRef]
- Tang, S.; Liao, Y.; Xu, Y.; Dang, Z.; Zhu, X.; Ji, G. Microbial coupling mechanisms of nitrogen removal in constructed wetlands: A review. Bioresour. Technol. 2020, 314, 123759. [Google Scholar] [CrossRef] [PubMed]
- Rosa, M.; Prado, C.; Podazza, G.; Interdonato, R.; González, J.A.; Hilal, M.; Prado, F.E. Soluble sugars: Metabolism, sensing and abiotic stress: A complex network in the life of plants. Plant Signal. Behav. 2009, 4, 388–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, J.; Zhao, X.; Song, X.; Gong, J. The inhibition and adaptability of four wetland plant species to high concentration of ammonia wastewater and nitrogen removal efficiency in constructed wetlands. Bioresour. Technol. 2016, 202, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Yang, X.; Ma, C.; Wang, Y.; Zhao, J. Melatonin enhances drought stress tolerance in maize through coordinated regulation of carbon and nitrogen assimilation. Plant Physiol. Biochem. 2021, 167, 958–969. [Google Scholar] [CrossRef]
- Sharma, A.; Wang, J.; Xu, D.; Tao, S.; Chong, S.; Yan, D.; Li, Z.; Yuan, H.; Zheng, B. Melatonin regulates the functional components of photosynthesis, antioxidant system, gene expression, and metabolic pathways to induce drought resistance in grafted Carya cathayensis plants. Sci. Total Environ. 2020, 713, 136675. [Google Scholar] [CrossRef]
- Marschnert, H.; Kirkby, E.A.; Engels, C. Importance of cycling and recycling of mineral nutrients within plants for growth and development. Bot. Acta 1997, 110, 265–273. [Google Scholar] [CrossRef]
- Noguchi, A.; Kageyama, M.; Shinmachi, F.; Schmidhalter, U.; Hasegawa, I. Potential for Using Plant Xylem Sap to Evaluate Inorganic Nutrient Availability in Soil: I. Influence of Inorganic Nutrients Present in the Rhizosphere on Those in the Xylem Sap of Luffa cylindrica Roem. Soil Sci. Plant Nutr. 2005, 51, 333–341. [Google Scholar] [CrossRef]
- Chen, Z.; Cao, X.; Niu, J. Effects of melatonin on morphological characteristics, mineral nutrition, nitrogen metabolism, and energy status in alfalfa under high-nitrate stress. Front. Plant Sci. 2021, 12, 694179. [Google Scholar] [CrossRef]
- Kvarnström, M.E.; Morel, C.A.L.; Krogstad, T. Plant-availability of phosphorus in filter substrates derived from small-scale wastewater treatment systems. Ecol. Eng. 2004, 22, 1–15. [Google Scholar] [CrossRef]
- Lee, C.; Fletcher, T.D.; Sun, G. Nitrogen removal in constructed wetland systems. Eng. Life Sci. 2009, 9, 11–22. [Google Scholar] [CrossRef]
- Vymazal, J. Removal of nutrients in various types of constructed wetlands. Sci. Total Environ. 2007, 380, 48–65. [Google Scholar] [CrossRef] [PubMed]
- Spieles, D.J.; Mitsch, W.J. The effects of season and hydrologic and chemical loading on nitrate retention in constructed wetlands: A comparison of low-and high-nutrient riverine systems. Ecol. Eng. 1999, 14, 77–91. [Google Scholar] [CrossRef]
- Xiao, R.; Han, Q.; Liu, Y.; Zhang, X.; Hao, Q.; Chai, Q.; Hao, Y.; Deng, J.; Li, X.; Ji, H. Melatonin Attenuates the Urea-Induced Yields Improvement Through Remodeling Transcriptome and Rhizosphere Microbial Community Structure in Soybean. Front. Microbiol. 2022, 13, 903467. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Lv, R.; Zhang, L.; Zi, X.; Zhou, H.; Tang, J. Melatonin is a promising silage additive: Evidence from microbiota and metabolites. Front. Microbiol. 2021, 12, 670764. [Google Scholar] [CrossRef]
- Ma, X.; Song, X.; Li, X.; Fu, S.; Li, M.; Liu, Y. Characterization of microbial communities in pilot-scale constructed wetlands with Salicornia for treatment of marine aquaculture effluents. Archaea 2018, 2018, 7819840. [Google Scholar] [CrossRef] [Green Version]
- Ansola, G.; Arroyo, P.; de Miera, L.S. Characterisation of the soil bacterial community structure and composition of natural and constructed wetlands. Sci. Total Environ. 2014, 473, 63–71. [Google Scholar] [CrossRef]
- Desta, A.F.; Assefa, F.; Leta, S.; Stomeo, F.; Wamalwa, M.; Njahira, M.; Appolinaire, D. Microbial community structure and diversity in an integrated system of anaerobic-aerobic reactors and a constructed wetland for the treatment of tannery wastewater in Modjo, Ethiopia. PLoS ONE 2014, 9, e115576. [Google Scholar] [CrossRef]
- Xie, F.; Ma, A.; Zhou, H.; Liang, Y.; Yin, J.; Ma, K.; Zhuang, G. Revealing fungal communities in alpine wetlands through species diversity, functional diversity and ecological network diversity. Microorganisms 2020, 8, 632. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, D.; Sun, Z.; Yang, Q.; Liu, Y.; Cao, T.; Chen, R.; Dzakpasu, M.; Wang, X.C. Stereoselective degradation pathway of amide chiral herbicides and its impacts on plant and bacterial communities in integrated vertical flow constructed wetlands. Bioresour. Technol. 2022, 351, 126997. [Google Scholar] [CrossRef]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Bashan, Y.; De-Bashan, L.E. How the plant growth-promoting bacterium Azospirillum promotes plant growth—A critical assessment. Adv. Agron. 2010, 108, 77–136. [Google Scholar]
- Shi, Y.; Liu, T.; Chen, S.; Quan, X. Accelerating anaerobic hydrolysis acidification of dairy wastewater in integrated floating-film and activated sludge (IFFAS) by using zero-valent iron (ZVI) composite carriers. Biochem. Eng. J. 2022, 177, 108226. [Google Scholar] [CrossRef]
- Willems, A.; Busse, J.; Goor, M.; Pot, B.; Falsen, E.; Jantzen, E.; Hoste, B.M.; Kersters, K.; Auling, G.; Ley, J. Hydrogenophaga, a new genus of hydrogen-oxidizing bacteria that includes Hydrogenophaga flava comb. nov. (Formerly Pseudomonas flava), Hydrogenophaga palleronii (Formerly Pseudomonas palleronii), Hydrogenophaga pseudoflava (Formerly Pseudomonas pseudoflava and “Pseudomonas carboxydoflava”), and Hydrogenophaga taeniospiralis (Formerly Pseudomonas taeniospiralis). Int. J. Syst. Evol. Microbiol. 1989, 39, 319–333. [Google Scholar]
- Chen, H.; Liu, G.; Wang, K.; Piao, C.; Ma, X.; Li, X.K. Characteristics of microbial community in EGSB system treating with oxytetracycline production wastewater. J. Environ. Manag. 2021, 295, 113055. [Google Scholar] [CrossRef] [PubMed]
- Koch, H.; Lücker, S.; Albertsen, M.; Kitzinger, K.; Herbold, C.; Spieck, E.; Nielsen, P.; Wagner, M.; Daims, H. Expanded metabolic versatility of ubiquitous nitrite-oxidizing bacteria from the genus Nitrospira. Proc. Natl. Acad. Sci. USA 2015, 112, 11371–11376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mekonnen, E.; Kebede, A.; Nigussie, A.; Kebede, G.; Tafesse, M. Isolation and characterization of urease-producing soil bacteria. Int. J. Microbiol. 2021, 2021, 8888641. [Google Scholar] [CrossRef]
- Chen, G.; Zhu, H.; Zhang, Y. Soil microbial activities and carbon and nitrogen fixation. Res. Microbiol. 2003, 154, 393–398. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Richness Index | Diversity Index | Coverage | ||||
|---|---|---|---|---|---|---|---|
| Sobs | Ace | Chao | Shannon | Simpson | |||
| Bacteria | CK | 1818 *** | 2235.7 *** | 2203.8 *** | 5.3004 *** | 0.020321 | 0.98911 |
| MT | 1056 | 1407.9 | 1399.1 | 3.9023 | 0.080602 *** | 0.9925 | |
| Fungi | CK | 358.25 ** | 377.81 ** | 375.95 ** | 3.0495 | 0.11156 * | 0.99945 |
| MT | 111.25 | 113.15 | 113.56 | 3.4637 * | 0.061382 | 0.99995 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, F.; Guo, J.; Hou, P.; Wang, Y.; Song, F.; Zhang, P.; Li, X. Impacts of Melatonin on Functionalities of Constructed Wetlands for Wastewater Treatment. Land 2022, 11, 2022. https://doi.org/10.3390/land11112022
Ye F, Guo J, Hou P, Wang Y, Song F, Zhang P, Li X. Impacts of Melatonin on Functionalities of Constructed Wetlands for Wastewater Treatment. Land. 2022; 11(11):2022. https://doi.org/10.3390/land11112022
Chicago/Turabian StyleYe, Fan, Junhong Guo, Pengfu Hou, Yongjun Wang, Fengbin Song, Peng Zhang, and Xiangnan Li. 2022. "Impacts of Melatonin on Functionalities of Constructed Wetlands for Wastewater Treatment" Land 11, no. 11: 2022. https://doi.org/10.3390/land11112022