
Vegetation Degradation of Guanshan Grassland Suppresses the Microbial Biomass and Activity of Soil



Abstract
:1. Introduction
2. Material and Methods
2.1. Study Areas
2.2. Experimental Design and Soil Sampling Collection
2.3. Analyses of the Soil Physicochemical Properties
2.4. Analyses of the Soil Microbial Biomass
2.5. Analyses of the Soil Microbial Activities
2.6. Statistics Analysis
3. Results
3.1. Changes of the Soil Physicochemical Properties Related to Vegetation Types
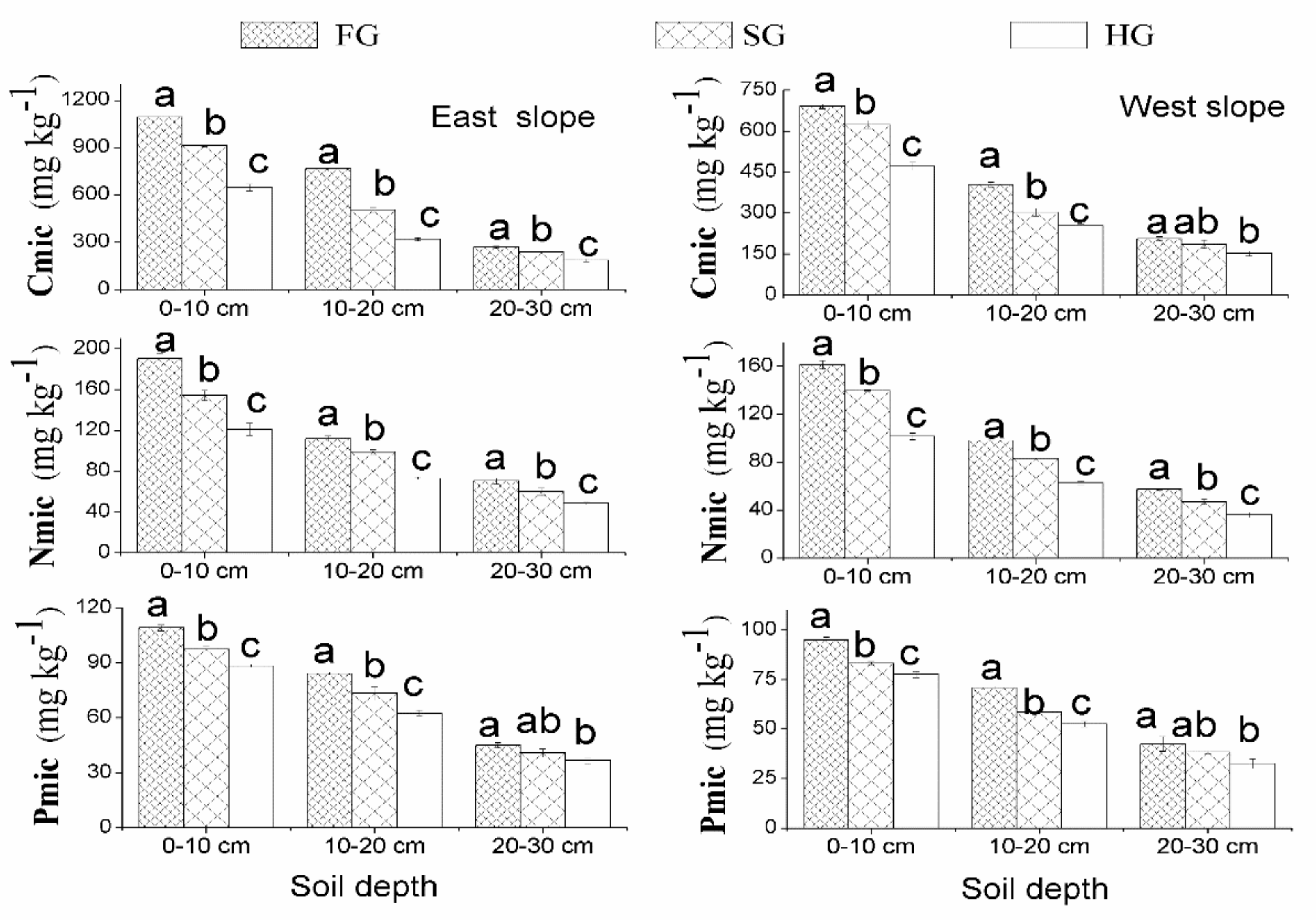
3.2. Changes in the Soil Microbial Biomass Related to Vegetation Types
3.3. Changes in the Soil Enzymatic Activities, BR, and qCO2 Related to Vegetation Types
3.4. Factors Affecting the Soil Microbial Parameters
4. Discussion
4.1. Impact of the Vegetation Types on the Soil Physicochemical Parameters, Microbial Biomass, Enzymatic Activities, BR, and qCO2
4.2. Impact of the Slope Aspect and Soil Depth on the Soil Microbial Biomass, Enzymatic Activities, BR, and qCO2
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ma, W.J.; Li, J.; Jimoh, S.O.; Zhang, Y.J.; Guo, F.H.; Ding, Y.; Li, X.L.; Hou, X.Y. Stoichiometric ratios support plant adaption to grazing moderated by soil nutrients and root enzymes. PeerJ 2019, 7, 15. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.C.; Wang, W.; Liu, H.Y.; Xu, X.T.; Zeng, H. Temperate grassland shifted from nitrogen to phosphorus limitation induced by degradation and nitrogen deposition: Evidence from soil extracellular enzyme stoichiometry. Ecol. Indic. 2019, 101, 453–464. [Google Scholar] [CrossRef]
- Yuan, Z.Q.; Jiang, X.J.; Liu, G.J.; Jin, H.J.; Chen, J.; Wu, Q.B. Responses of soil organic carbon and nutrient stocks to human-induced grassland degradation in a Tibetan alpine meadow. Catena 2019, 178, 40–48. [Google Scholar] [CrossRef]
- Zhou, W.; Gang, C.; Zhou, L.; Chen, Y.; Li, J.; Ju, W.; Odeh, I. Dynamic of grassland vegetation degradation and its quantitative assessment in the northwest China. Acta. Oecol. 2014, 55, 86–96. [Google Scholar] [CrossRef]
- Hu, Y.B.; Guo, T.; Chen, J. Research on characteristics of vegetation community structure at different altitudes in Guanshan grasslands. Pratacult. Sci. 2013, 30, 190–195. (In Chinese) [Google Scholar]
- Ren, Z.Y.; Song, B.P.; Zhang, H. Ecological environmental protection and construction in the development of Guanshan Grassland Tourism Scenic Area. J. Dese. Rese. 2000, 1, 87–90. (In Chinese) [Google Scholar]
- Dong, S.K.; Wen, L.; Yi, Y.Y.; Wang, X.X.; Zhu, L.; Li, X.Y. Soil-quality effects of grassland degradation and restoration on the Qinghai–Tibetan plateau. Soil Sci. Soc. Am. J. 2012, 76, 2256–2264. [Google Scholar] [CrossRef]
- Liu, S.B.; Zamanian, K.; Schleuss, P.M.; Zarebanadkouki, M.; Kuzyakov, Y. Degradation of Tibetan grasslands: Consequences for carbon and nutrient cycles. Agric. Ecosyst. Environ. 2018, 252, 93–104. [Google Scholar] [CrossRef]
- Feng, C.; Ma, Y.H.; Jin, X.; Wang, Z.; Ma, Y.; Fu, S.L.; Chen, H.Y.H. Soil enzyme activities increase following restoration of degraded subtropical forests. Geoderma 2019, 351, 180–187. [Google Scholar] [CrossRef]
- Bargali, K.; Manral, V.; Padalia, K.; Bargali, S.S.; Upadhyay, V.P. Effect of vegetation type and season on microbial biomass carbon in Central Himalayan forest soils, India. Catena 2018, 171, 125–135. [Google Scholar] [CrossRef]
- Kuscu, I.S.K. Changing of soil properties and urease–catalase enzyme activity depending on plant type and shading. Environ. Monit. Assess. 2019, 191, 178. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.H.; Domsch, K.H. Determination of ecophysiological maintenance carbon requirements of soil microorganisms in a dormant stare. Biol. Fert. Soils 1985, 1, 81–89. [Google Scholar] [CrossRef]
- Zhao, C.; Long, J.; Liao, H.K.; Zheng, C.L.; Li, J.; Liu, L.F.; Zhang, M.J. Dynamics of soil microbial communities following vegetation succession in a karst mountain ecosystem, southwest China. Sci. Rep. 2019, 9, 2160. [Google Scholar] [CrossRef]
- Ramirez, K.S.; Craine, J.M.; Fierer, N. Consistent effects of nitrogen amendments on soil microbial communities and processes across biomes. Glob. Chang. Biol. 2012, 18, 1918–1927. [Google Scholar] [CrossRef]
- Pan, F.J.; Zhang, W.; Liang, Y.M.; Liu, S.J.; Wang, K.L. Increased associated effects of topography and litter and soil nutrients on soil enzyme activities and microbial biomass along vegetation successions in karst ecosystem, southwestern China. Environ. Sci. Pollut. Res. 2018, 25, 16979–16990. [Google Scholar] [CrossRef]
- Rasouli-Sadaghiani, M.H.; Barin, M.; Siavash, S.M.; Damalas, C.A.; Ghodrat, K. Soil quality of an Iranian forest ecosystem after conversion to various types of land use. Environ. Monit. Assess. 2018, 190, 447. [Google Scholar] [CrossRef]
- Lungmuana; Singh, S.B.; Choudhury, B.U.; Vanthawmliana; Saha, S.; Hnamte, V. Transforming jhum to plantations: Effect on soil microbiological and biochemical properties in the foot hills of North Eastern Himalayas, India. Catena 2019, 177, 84–91. [Google Scholar]
- Wu, J.; Joergensen, R.G.; Pommerening, B.; Chaussod, R.; Brookes, P.C. Measurement of soil microbial biomass C by fumigation-extraction-an automated procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- Lu, R.K. Analytical Methods for Soil and Agricultural Chemistry; China Agricultural Science and Technology Press: Beijing, China, 1999; pp. 107–150. (in Chinese) [Google Scholar]
- Bremner, J.M. Nitrogen-Total. In Methods of Soil Analysis. Part 3. Soil Science Society of America; Sparks, D.L., Ed.; American Society of Agronomy: Madison, WI, USA, 1996; pp. 1085–1086. [Google Scholar]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, N.C. Estimation of Available Phosphous in Soil by Extraction with Sodium Bicorbonate; United States Department of Agriculture: Washington, DC, USA, 1954; p. 939.
- Bao, S.D. Agro-Chemical Analysis of Soil; China Agriculture Press: Beijing, China, 2000; pp. 22–108. (In Chinese) [Google Scholar]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil organic nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soils. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Brookes, P.C.; Powlson, D.S.; Jenkinson, D.S. Measurement of microbial biomass phosphorus in soil. Soil Biol. Biochem. 1982, 14, 319–329. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Liu, Y.M.; Xing, Z.S.; Yang, H.Y. Effect of biological soil crusts on microbial activity in soils of the Tengger Desert (China). J. Arid Environ. 2017, 144, 201–211. [Google Scholar] [CrossRef]
- Cui, Y.X.; Fang, L.C.; Guo, X.B.; Wang, X.; Wang, Y.Q.; Zhang, Y.J.; Zhang, X.C. Responses of soil bacterial communities, enzyme activities, and nutrients to agricultural-to-natural ecosystem conversion in the Loess Plateau, China. J. Soils Sediments 2019, 19, 1427–1440. [Google Scholar] [CrossRef]
- Muhammad, N.; Dai, Z.M.; Xiao, K.C.; Meng, J.; Brookes, P.C.; Liu, X.M.; Wang, H.Z.; Wu, J.J.; Xu, J.M. Changes in microbial community structure due to biochars generated from different feedstocks and their relationships with soil chemical properties. Geoderma 2014, 226–227, 270–278. [Google Scholar] [CrossRef]
- Balota, E.L.; Yada, I.F.; Amaral, H.; Nakatani, A.S.; Dick, R.P.; Coyne, M.S. Long-term land use influences soil microbial biomass P and S, phosphatase and arylsulfatase activities, and S mineralization in a Brazilian Oxisol. Land Degrad. Dev. 2014, 25, 397–406. [Google Scholar] [CrossRef]
- Chandregowda, H.; Murthy, K.; Bagchi, S. Woody shrubs increase soil microbial functions and multifunctionality in a tropical semi-arid grazing ecosystem. J. Arid Environ. 2018, 155, 65–72. [Google Scholar] [CrossRef]
- Tiwari, S.; Singh, C.; Boudh, S.; Rai, P.K.; Gupta, V.K.; Singh, J.S. Land use change: A key ecological disturbance declines soil microbial biomass in dry tropical uplands. J. Environ. Manag. 2019, 242, 1–10. [Google Scholar] [CrossRef]
- Sun, B.; Hallett, P.D.; Caul, S.; Daniell, T.J.; Hopkins, D.W. Distribution of soil carbon and microbial biomass in arable soils under different tillage regimes. Plant Soil 2010, 338, 17–25. [Google Scholar] [CrossRef]
- Xiao, L.; Bi, Y.L.; Du, S.Z.; Wang, Y.; Guo, C. Effects of re-vegetation type and arbuscular mycorrhizal fungal inoculation on soil enzyme activities and microbial biomass in coal mining subsidence areas of Northern China. Catena 2019, 177, 202–209. [Google Scholar] [CrossRef]
- Liu, W.X.; Xu, W.H.; Hong, J.P.; Wan, S.Q. Interannual variability of soil microbial biomass and respiration in responses to topography, annual burning and N addition in a semiarid temperate steppe. Geoderma 2010, 158, 259–267. [Google Scholar] [CrossRef]
- Tian, Y.W.; Qiao, D.; Xu, S.J.; Wang, N. Effects of tree species and topography on soil and microbial biomass stoichiometry in Funiu Mountain, China. BMC Ecol. 2020, 20, 67. [Google Scholar] [CrossRef]
- Johnson, D.; Leake, J.R.; Read, D.J. Liming and nitrogen fertilization affects phosphatase activities, microbial biomass and mycorrhizal colonisation in upland grassland. Plant Soil 2005, 271, 157–164. [Google Scholar] [CrossRef]
- Zhang, W.J.; Li, R.R.; Ai, X.Y.; Chen, J.; Xu, W.N.; Li, W.; Ai, Y.W. Enzyme activity and microbial biomass availability in artificial soils on rock-cut slopes restored with outside soil spray seeding (OSSS): Influence of topography and season. J. Environ. Manag. 2018, 211, 287–295. [Google Scholar] [CrossRef]
- Ghee, C.; Neilson, R.; Hallet, P.D.; Robinson, D.; Paterson, E. Priming of soil organic matter mineralisation is intrinsically insensitive to temperature. Soil Biol. Biochem. 2013, 66, 20–28. [Google Scholar] [CrossRef]
- Huang, Y.M.; Liu, D.; An, S.S. Effects of slope aspect on soil nitrogen and microbial properties in the Chinese Loess region. Catena 2015, 125, 135–145. [Google Scholar] [CrossRef]
- Joergensen, R.G.; Kubler, H.; Meyer, B.; Wolters, V. Microbial biomass phosphorus in soils of beech (fagus-sylvatica l) forests. Biol. Fert. Soils 1995, 19, 215–219. [Google Scholar]
- Wardle, D.A.; Ghani, A. A critique of the microbial metabolic quotient (qCO2) as a bio-indicator of disturbance and ecosystem development. Soil Biol. Biochem. 1995, 27, 1601–1610. [Google Scholar] [CrossRef]
- Wang, H.Y.; Wu, J.Q.; Li, G.; Yan, L.J. Changes in soil carbon fractions and enzyme activities under different vegetation types of the northern Loess Plateau. Ecol. Evol. 2020, 10, 12211–12223. [Google Scholar] [CrossRef] [PubMed]
- Oduor, C.O.; Karanja, N.K.; Onwonga, R.N.; Mureithi, S.M.; Pelster, D.; Nyberg, G. Enhancing soil organic carbon, particulate organic carbon and microbial biomass in semi-arid rangeland using pasture enclosures. BMC Ecol. 2018, 18, 45. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Types | Site | Altitude (m) | Dominant Species [5] | Vegetation Coverage | Soil Types |
|---|---|---|---|---|---|
| Forestgrass land (FG) | Mountain top | 2350–2400 | Trees and shrubs, including Pinus tabuliformis Carr., Salix pseudotangii C. Wang et C. Y. Yu, Hippophae rhamnoides Linn., Populus tomentosa Carr., Rhododendron simsii Planch., etc. | 60%–70% | Yellow soil |
| Dwarf grasses, including Festuca rubra Linn., Carex lehmanii Drej., Medicago lupulina Linn., etc. | 20%–30% | ||||
| Shrub grassland (SG) | Mountain middle | 2100–2200 | Trees and shrubs, including S. pseudotangii C. Wang et C. Y. Yu, Betula albo-sinensis Burk., Populus davidiana Dode, etc. | 30%–40% | Yellow soil |
| Dwarf grasses with coverage, including F. rubra Linn., Leontopodium leontopodiodes (Willd.) Beauv., C. lehmanii Drej., Elymus daburicus Turcz., etc. | 40%–50% | ||||
| Herb grassland (HG) | Mountain base | 2050–2100 | Trees and shrubs Dwarf grass, including Duchesnea indica (Andr.) Fock., Ranunculus japonicas Thunb., C. lehmanii Drej., Cyperus sp., Anemone rivularis Buch.-Ham., etc. | <1% 70% | Yellow soil |
| Slope Aspect | Vegetation Types | SOC (g kg−1) | TN (g kg−1) | AN (mg kg−1) | TP (g kg−1) | AP (mg kg−1) | TK (g kg−1) | AK (mg kg−1) | pH | SM |
|---|---|---|---|---|---|---|---|---|---|---|
| East Slope | Forest grassland (FG) | 81.48 ± 1.29 a | 3.83 ± 0.02 a | 236.37 ± 4.72a | 36.09 ± 0.27 a | 18.33 ± 0.42 a | 9.90 ± 0.08 a | 173.44 ± 1.66 a | 6.43 ± 0.01 c | 0.25 ± 0.01 a |
| Shrub grassland (SG) | 77.19 ± 0.63 b | 3.30 ± 0.05 b | 200.20 ± 2.43 b | 25.13 ± 0.32 b | 9.79 ± 0.18 b | 8.47 ± 0.04 b | 163.33 ± 1.53 b | 6.52 ± 0.01 b | 0.22 ± 0.01 ab | |
| Herb grassland (HG) | 71.84 ± 0.99 c | 2.24 ± 0.03 c | 169.87 ± 2.33 c | 21.75 ± 0.30 c | 4.08 ± 0.08 c | 8.40 ± 0.08 b | 132.89 ± 1.24 c | 6.61 ± 0.02 a | 0.20 ± 0.01 b | |
| West Slope | Forest grassland (FG) | 61.60 ± 0.40 a | 2.75 ± 0.06 a | 204.87 ± 3.56 a | 25.27 ± 0.37 a | 17.21 ± 0.53 a | 8.38 ± 0.06 a | 139.44 ± 1.64 a | 6.47 ± 0.01 c | 0.24 ± 0.01 a |
| Shrub grassland (SG) | 58.55 ± 2.73 a | 2.04 ± 0.04 b | 194.14 ± 2.70 a | 19.49 ± 0.14 b | 8.30 ± 0.10 b | 8.06 ± 0.04 b | 146.78 ± 3.82 a | 6.54 ± 0.02 b | 0.23 ± 0.01 a | |
| Herb grassland (HG) | 41.73 ± 0.70 b | 1.60 ± 0.03 c | 164.42 ± 3.04 b | 16.61 ± 1.04 c | 3.95 ± 0.14 c | 7.96 ± 0.02 b | 117.22 ± 3.19 b | 6.68 ± 0.02 a | 0.23 ± 0.00 a |
| Source of Variation | df | Mean Squares | ||||
|---|---|---|---|---|---|---|
| Cmic | Nmic | Pmic | BR | qCO2 | ||
| Vegetation types Slope aspect | 2 | 1,300,408.50 *** | 38,366.48 *** | 12,414.70 *** | 6956.16 *** | 0.07 *** |
| 1 | 454,039.52 *** | 3332.16 *** | 1251.85 *** | 4933.85 *** | 0.01 ns | |
| Soil depth | 2 | 245,950.88 *** | 7663.83 *** | 1175.46 *** | 95,075.23 *** | 0.67 *** |
| Vegetation types × slope aspect | 2 | 68,045.06 *** | 99.40 * | 145.95 *** | 916.38 *** | 0.00 * |
| Vegetation types × soil depth | 4 | 33,478.86 *** | 711.99 *** | 58.10 ** | 505.65 *** | 0.01 ** |
| Slope aspect × soil depth | 2 | 38,048.38 *** | 27.46 ns | 6.02 ns | 179.20 * | 0.00 ns |
| Vegetation types × slope aspect × soil depth | 4 | 7160.85 *** | 31.12 ns | 5.32 ns | 99.80 ns | 0.01 * |
| Source of Variation | df | Mean Squares | ||||||
|---|---|---|---|---|---|---|---|---|
| Urease Activity | Sucrase Activity | Catalase Activity | Dehydrogenase Activity | Alkaline Phosphatase Activity | Protease Activity | Cellulase Activity | ||
| Vegetation types | 2 | 1.18 *** | 0.3 5*** | 0.33 *** | 82.64 *** | 958.66 *** | 6151.91 *** | 3876.34 *** |
| Slope aspect | 1 | 0.26 *** | 1.85 *** | 0.17 *** | 171.36 *** | 1393.64 *** | 3112.98 *** | 1467.33 *** |
| Soil depth | 2 | 1.69 *** | 2.37 *** | 0.20 *** | 271.41 *** | 1232.96 *** | 12,354.36 *** | 10,984.66 *** |
| Vegetation types × slope aspect | 2 | 0.20 *** | 0.02 *** | 0.00 *** | 2.66 *** | 308.95 *** | 403.73 *** | 359.24 **** |
| Vegetation types × soil depth | 4 | 0.04 *** | 0.01 *** | 0.00 *** | 2.29 *** | 40.50 *** | 74.92 ** | 533.99 *** |
| Slope aspect × soil depth | 2 | 0.00 ns | 0.04 *** | 0.01 *** | 0.11 ns | 17.91 * | 48.95 ns | 13.68 ns |
| Vegetation types × slope aspect × soil depth | 4 | 0.01 ns | 0.0 ns | 0.00 * | 0.49 ** | 38.68 *** | 34.82 ns | 24.86 ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Yang, H.; Xing, Z.; Zou, Y.; Cui, Z. Vegetation Degradation of Guanshan Grassland Suppresses the Microbial Biomass and Activity of Soil. Land 2021, 10, 203. https://doi.org/10.3390/land10020203
Liu Y, Yang H, Xing Z, Zou Y, Cui Z. Vegetation Degradation of Guanshan Grassland Suppresses the Microbial Biomass and Activity of Soil. Land. 2021; 10(2):203. https://doi.org/10.3390/land10020203
Chicago/Turabian StyleLiu, Yanmei, Hangyu Yang, Zisheng Xing, Yali Zou, and Zheming Cui. 2021. "Vegetation Degradation of Guanshan Grassland Suppresses the Microbial Biomass and Activity of Soil" Land 10, no. 2: 203. https://doi.org/10.3390/land10020203