Investigation of Pathogenic Bacterial Transport by Waterbirds: A Case Study of Flooded and Non-Flooded Rice Systems in Mississippi

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Soil Samplingy
2.3. Water Sampling
2.4. Quantifying Bird Fecal Matter
2.5. Pathogens
2.5.1. Soil
2.5.2. Water
3. Statistical Analysis
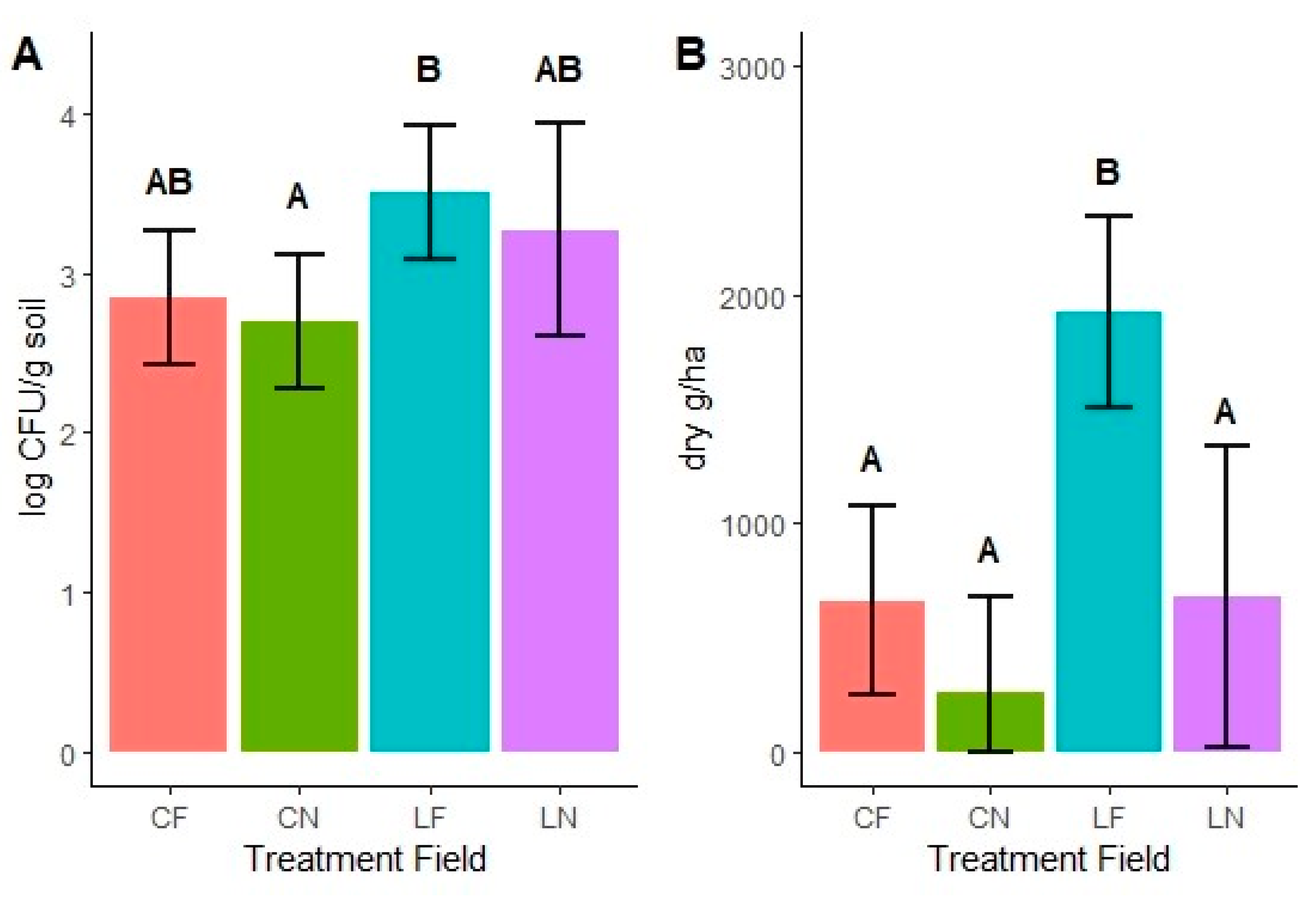
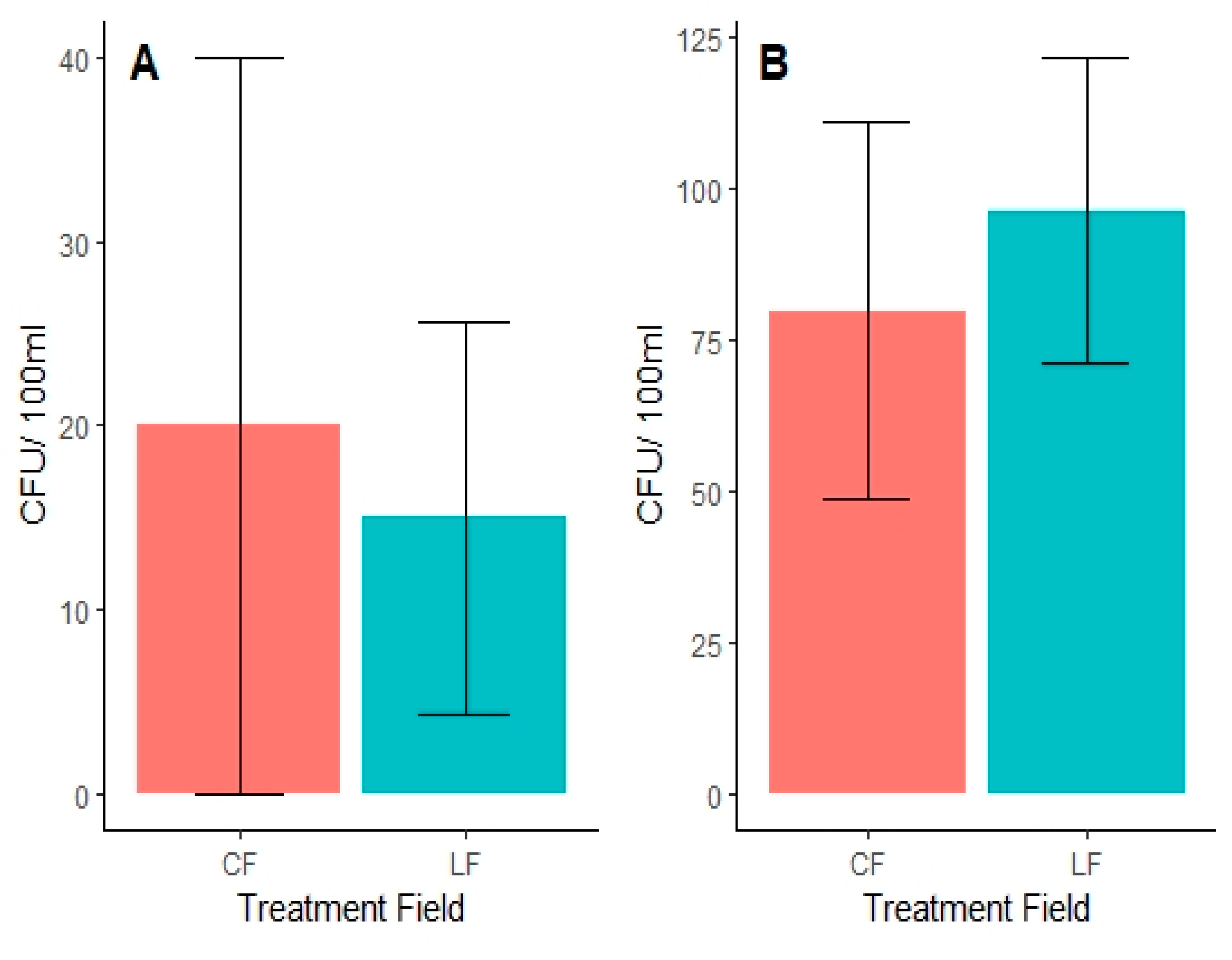
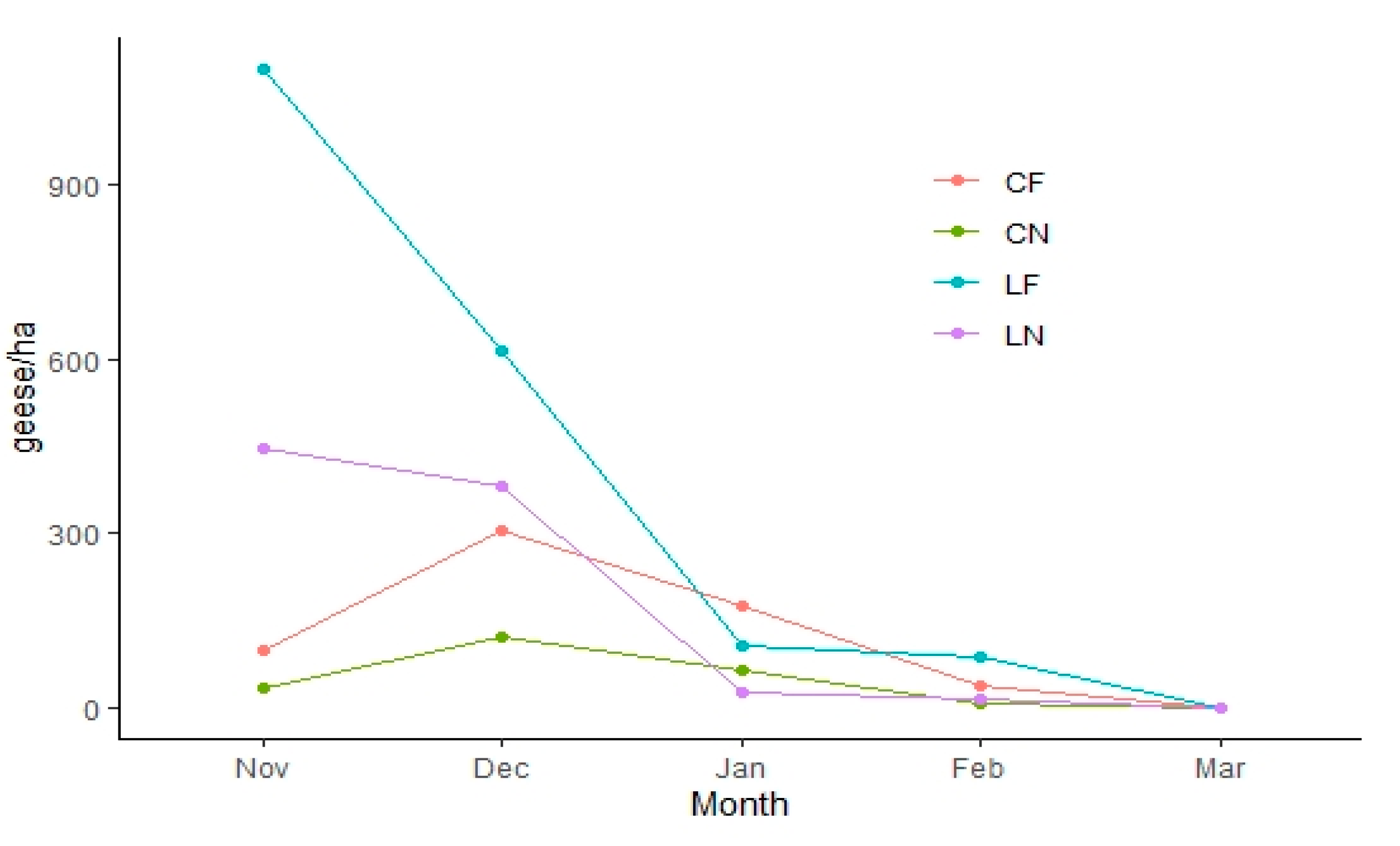
4. Results
5. Discussion
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Elphick, C.S. Functional Equivalency between Rice Fields and Seminatural Wetland Habitats. Conserv. Biol. 2000, 14, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Elphick, C.S.; Oring, L.W. Conservation implications of flooding rice fields on winter waterbird communities. Agric. Ecosyst. Environ. 2003, 94, 17–29. [Google Scholar] [CrossRef]
- Tourenq, C.; Bennetts, R.E.; Kowalski, H.; Vialet, E.; Lucchesi, J.-L.; Kayser, Y.; Isenmann, P. Are ricefields a good alternative to natural marshes for waterbird communities in the Camargue, southern France? Biol. Conserv. 2001, 100, 335–343. [Google Scholar] [CrossRef]
- Petrie, M.; Brasher, M.; James, D. Estimating the Biological and Economic Contributions that Rice Habitats Make in Support of North American Waterfowl Populations; The Rice Foundation: Stuttgart, AR, USA, 2014. [Google Scholar]
- Williams, C.K.; Dugger, B.D.; Brasher, M.G.; Coluccy, J.M.; Cramer, D.M.; Eadie, J.M.; Gray, M.J.; Hagy, H.M.; Livolsi, M.; McWilliams, S.R.; et al. Estimating habitat carrying capacity for migrating and wintering waterfowl: Considerations, pitfalls and improvements. Wildfowl 2014, 4, 407–435. [Google Scholar]
- National Agriculture Statistics Service. Crop Production 2018 Summary; National Agriculture Statistics Service: Washington, DC, USA, 2019.
- Kross, J.P.; Kaminski, R.M.; Reinecke, K.J.; Pearse, A.T. Conserving waste rice for wintering waterfowl in the mississippi alluvial valley. J. Wildl. Manag. 2008, 72, 1383–1387. [Google Scholar] [CrossRef]
- Abraham, K.F.; Jefferies, R.L.; Alisauskas, R.T. The dynamics of landscape change and snow geese in mid-continent North America. Glob. Chang. Biol. 2005, 11, 841–855. [Google Scholar] [CrossRef] [Green Version]
- Bird, J.A.; Pettygrove, G.S.; Eadie, J.M. The impact of waterfowl foraging on the decomposition of rice straw: Mutual benefits for rice growers and waterfowl. J. Appl. Ecol. 2000, 37, 728–741. [Google Scholar] [CrossRef]
- Manley, S.W. Ecological and Agricultural Values of Winter-Flooded Ricefields in Mississippi; Mississippi State University Library: Starkville, MS, USA, 1999. [Google Scholar]
- Firth, A.G.; Baker, B.H.; Brooks, J.P.; Smith, R.; Iglay, R.B.; Davis, J.B. Low external input sustainable agriculture: Winter flooding in rice fields increases bird use, fecal matter and soil health, reducing fertilizer requirements. Agric. Ecosyst. Environ. 2020, 300, 106962. [Google Scholar] [CrossRef]
- Fallacara, D.M.; Monahan, C.M.; Morishita, T.Y.; Wack, R.F. Fecal shedding and antimicrobial susceptibility of selected bacterial pathogens and a survey of intestinal parasites in free-living waterfowl. Avian Dis. 2001, 45, 128–135. [Google Scholar] [CrossRef]
- Hussong, D.; Damaré, J.M.; Limpert, R.J.; Sladen, W.J.; Weiner, R.M.; Colwell, R.R. Microbial impact of Canada geese (Branta canadensis) and whistling swans (Cygnus columbianus columbianus) on aquatic ecosystems. Appl. Environ. Microbiol. 1979, 37, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Chrząstek, K.; Kuczkowski, M.; Wieliczko, A.K.; Bednarek, K.J.; Wieliczko, A.; Bossa, L.L. Molecular Epidemiologic Investigation of Polish Avian Pasteurella multocida Strains Isolated from Fowl Cholera Outbreaks Showing Restricted Geographical and Host-Specific Distribution. Avian Dis. 2012, 56, 529–536. [Google Scholar] [CrossRef]
- Waldenström, J.; Broman, T.; Carlsson, I.; Hasselquist, D.; Achterberg, R.P.; Wagenaar, J.A.; Olsen, B. Prevalence of Campylobacter jejuni, Campylobacter lari, and Campylobacter coli in different ecological guilds and taxa of migrating birds. Appl. Environ. Microbiol. 2002, 68, 5911–5917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubálek, Z.; Hubá, Z. An Annotated Checklist of Pathogenic Microorganims Associated with Migratory Birds. J. Wildl. Dis. 2004, 40, 639–659. [Google Scholar]
- Keawcharoen, J.; Van Riel, D.; Van Amerongen, G.; Bestebroer, T.; Beyer, W.E.; Van Lavieren, R.; Osterhaus, A.D.M.; Fouchier, R.A.M.; Juiken, T. Wild ducks as vectors of HPAIV (H5N1). Emerg. Infect. Dis. 2008, 14, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Dessborn, L.; Hessel, R.; Elmberg, J. Geese as vectors of nitrogen and phosphorous to freshwater systems. Inland Waters 2016, 6, 111–122. [Google Scholar] [CrossRef]
- Navedo, J.G.; Hahn, S.; Parejo, M.; Abad-Gómez, J.M.; Gutiérrez, J.S.; Villegas, A.; Sánchez-Guzmán, J.M.; Masero, J.A. Unravelling trophic subsidies of agroecosystems for biodiversity conservation: Food consumption and nutrient recycling by waterbirds in Mediterranean rice fields. Sci. Total Environ. 2015, 511, 288–297. [Google Scholar] [CrossRef]
- Wobeser, G.A. Diseases of Wild Waterfowl; Springer: New York, NY, USA, 1997; ISBN 9781461559511. [Google Scholar]
- Sixl, W.; Karpísková, R.; Hubálek, Z.; Halouzka, J.; Mikulásková, M.; Salava, J. Campylobacter spp. and Salmonella spp. in black-headed gulls (Larus ridibundus). Cent. Eur. J. Public Health 1997, 5, 24–26. [Google Scholar]
- Hudson, C.R.; Quist, C.; Lee, M.D.; Keyes, K.; Dodson, S.V.; Morales, C.; Sanchez, S.; White, D.G.; Maurer, J.J. Genetic relatedness of Salmonella isolates from nondomestic birds in Southeastern United States. J. Clin. Microbiol. 2000, 38, 1860–1865. [Google Scholar] [CrossRef] [Green Version]
- Reed, K.D.; Meece, J.K.; Henkel, J.S.; Shukla, S.K. Birds, migration and emerging zoonoses: West nile virus, lyme disease, influenza A and enteropathogens. Clin. Med. Res. 2003, 1, 5–12. [Google Scholar] [CrossRef] [Green Version]
- EPA. National Water Quality Inventory: Report to Congress; United States Environmental Protection Agency: Washington, DC, USA, 2017.
- EPA. Purposes of Monitoring for Pathogens and Indicators; United States Environmental Protection Agency: Washington, DC, USA, 2013.
- Patil, G.P. Composite Sampling. In Encyclopedia of Environmetrics; John Wiley & Sons, Ltd: Chichester, UK, 2013; ISBN 9780470057339. [Google Scholar]
- Brooks, J.P.; McLaughlin, M.R.; Scheffler, B.; Miles, D.M. Microbial and antibiotic resistant constituents associated with biological aerosols and poultry litter within a commercial poultry house. Sci. Total Environ. 2010, 408, 4770–4777. [Google Scholar] [CrossRef]
- EPA. EPA Recreational Water Quality Criteria; United States Environmental Protection Agency: Washington, DC, USA, 2012.
- Marty, J.R. Estimates of Waste Rice, Natural Seeds, and Wetland Birds in Gulf Coast Prairie Ricelands during Fall-Winter; Mississippi State University: Starkville, MS, USA, 2017. [Google Scholar]
- McNeil, R.; Drapeau, P.; Goss-Custard, J.D. The occurrence and adaptive significance of nocturnal habitats in waterfowl. Biol. Rev. 1992, 67, 381–419. [Google Scholar] [CrossRef]
- Gersberg, R.M.; Lyon, S.R.; Brenner, R.; Elkins, B.V. Fate of viruses in artificial wetlands. Appl. Environ. Microbiol. 1987, 53, 731–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, M.B.; Griffin, P.; Seabridge, J.K.; Dhobie, D. Removal of bacteria in subsurface flow wetlands. Water Sci. Technol. 1997, 35, 109–116. [Google Scholar] [CrossRef]
- Karim, M.R.; Manshadi, F.D.; Karpiscak, M.M.; Gerba, C.P. The persistence and removal of enteric pathogens in constructed wetlands. Water Res. 2004, 38, 1831–1837. [Google Scholar] [CrossRef]
- Russo, S.A.; Hunn, J.; Characklis, G.W. Considering bacteria-sediment associations in microbial fate and transport modeling. J. Environ. Eng. 2011, 137, 697–706. [Google Scholar] [CrossRef]
- Garzio-Hadzick, A.; Shelton, D.R.; Hill, R.L.; Pachepsky, Y.A.; Guber, A.K.; Rowland, R. Survival of manure-borne E. coli in streambed sediment: Effects of temperature and sediment properties. Water Res. 2010, 44, 2753–2762. [Google Scholar] [CrossRef]
- Pernthaler, J. Predation on prokaryotes in the water column and its ecological implications. Nat. Rev. Microbiol. 2005, 3, 537–546. [Google Scholar] [CrossRef]
- Thingstad, T.; Lignell, R. Theoretical models for the control of bacterial growth rate, abundance, diversity and carbon demand. Aquat. Microb. Ecol. 1997, 13, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Gasol, J.M.; Pedrós-Alió, C.; Vaqué, D. Regulation of bacterial assemblages in oligotrophic plankton systems: Results from experimental and emperical approaches. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2002, 81, 435–452. [Google Scholar] [CrossRef]
- Howell, J.M.; Coyne, M.S.; Cornelius, P.L.; Howell, J.M. Effect of sediment particle size and temperature on fecal bacteria mortality rates and the fecal coliform/fecal streptococci ratio repository citation. J. Environ. Qual. 1996, 25, 1216–1220. [Google Scholar] [CrossRef] [Green Version]
- Sherer, B.M.; Miner, J.R.; Moore, J.A.; Buckhouse, J.C. Indicator Bacterial Survival in Stream Sediments. J. Environ. Qual. 1992, 21, 591–595. [Google Scholar] [CrossRef] [Green Version]
- Davies, C.M.; Bavor, H.J. The fate of stormwater-associated bacteria in constructed wetland and water pollution control pond systems. J. Appl. Microbiol. 2000, 89, 349–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decamp, O.; Warren, A. Investigation of Escherichia coli removal in various designs of subsurface flow wetlands used for wastewater treatment. Ecol. Eng. 2000, 14, 293–299. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef]
- Shikuma, N.J.; Hadfield, M.G. Marine bioflims on submerged surfaces are a reservoir for Escherichia coli and Vibrio cholerae. Biofouling 2010, 26, 39–46. [Google Scholar] [CrossRef]
- Mallin, M.A.; McIver, M.R.; Robuck, A.R.; Dickens, A.K. Industrial swine and poultry production causes chronic nutrient and fecal microbial stream pollution. Water Air Soil Pollut. 2015, 226. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Firth, A.G.; Baker, B.H.; Brooks, J.P.; Smith, R.; Iglay, R.B.; Davis, J.B. Investigation of Pathogenic Bacterial Transport by Waterbirds: A Case Study of Flooded and Non-Flooded Rice Systems in Mississippi. Water 2020, 12, 1833. https://doi.org/10.3390/w12061833
Firth AG, Baker BH, Brooks JP, Smith R, Iglay RB, Davis JB. Investigation of Pathogenic Bacterial Transport by Waterbirds: A Case Study of Flooded and Non-Flooded Rice Systems in Mississippi. Water. 2020; 12(6):1833. https://doi.org/10.3390/w12061833
Chicago/Turabian StyleFirth, Alexandra G., Beth H. Baker, John P. Brooks, Renotta Smith, Raymond B. Iglay, and J. Brian Davis. 2020. "Investigation of Pathogenic Bacterial Transport by Waterbirds: A Case Study of Flooded and Non-Flooded Rice Systems in Mississippi" Water 12, no. 6: 1833. https://doi.org/10.3390/w12061833