Seasonal Distributions of Phytoplankton and Environmental Factors Generate Algal Blooms in the Taehwa River, South Korea

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Physical Environment
3.2. Water Environment
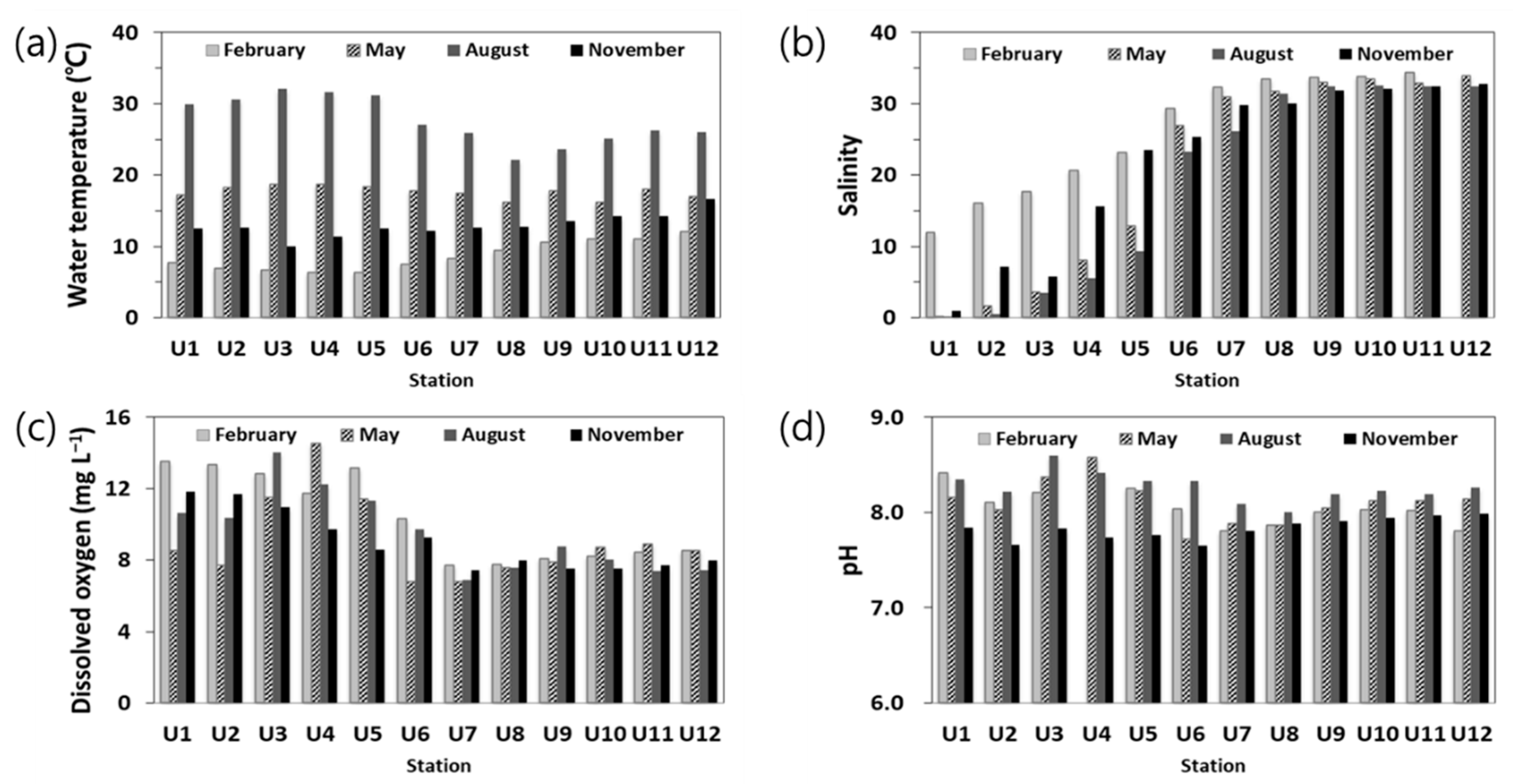
3.2.1. Temperature, Salinity, Dissolved Oxygen (DO), and pH
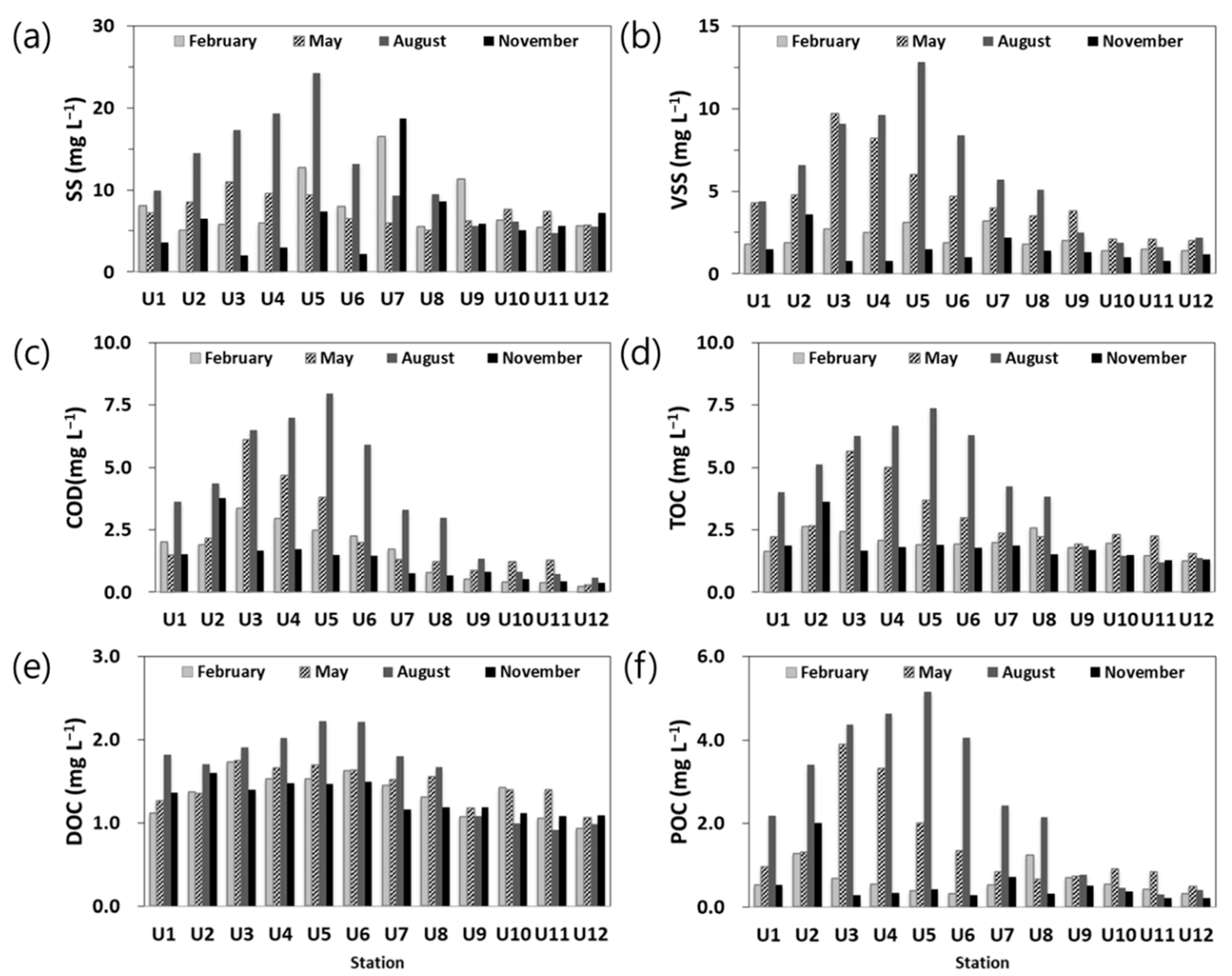
3.2.2. Organic Contents
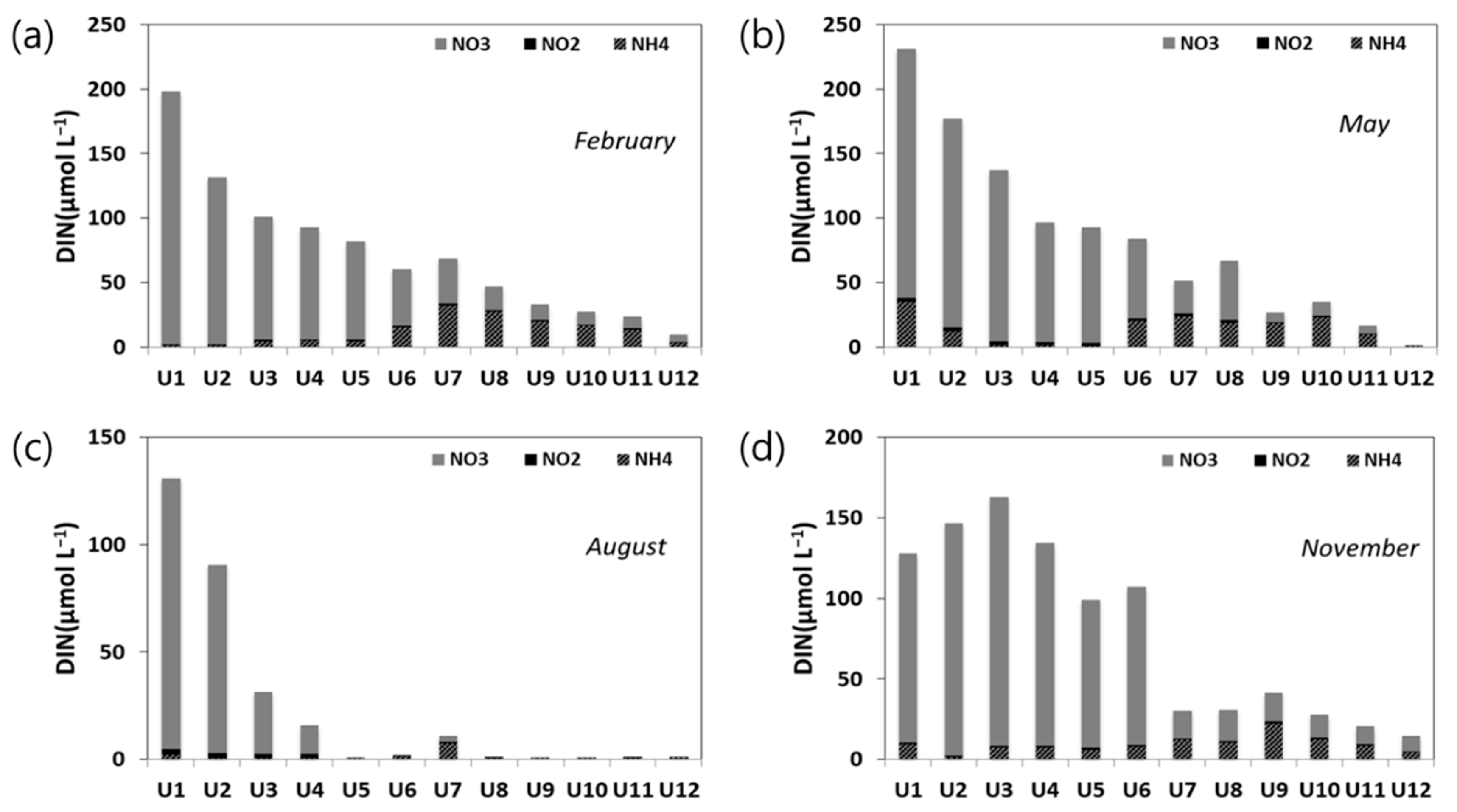
3.2.3. Nutrients
3.2.4. Chl.a
3.3. Phytoplankton
3.3.1. Species Composition
3.3.2. Phytoplankton Population and Dominant Species
4. Discussion
4.1. Seasonal Environmental Factors According to Water Flow
4.2. Water Retention Time and Algal Blooms
4.3. Water Quality Assessment
4.4. Chl.a and Biomass
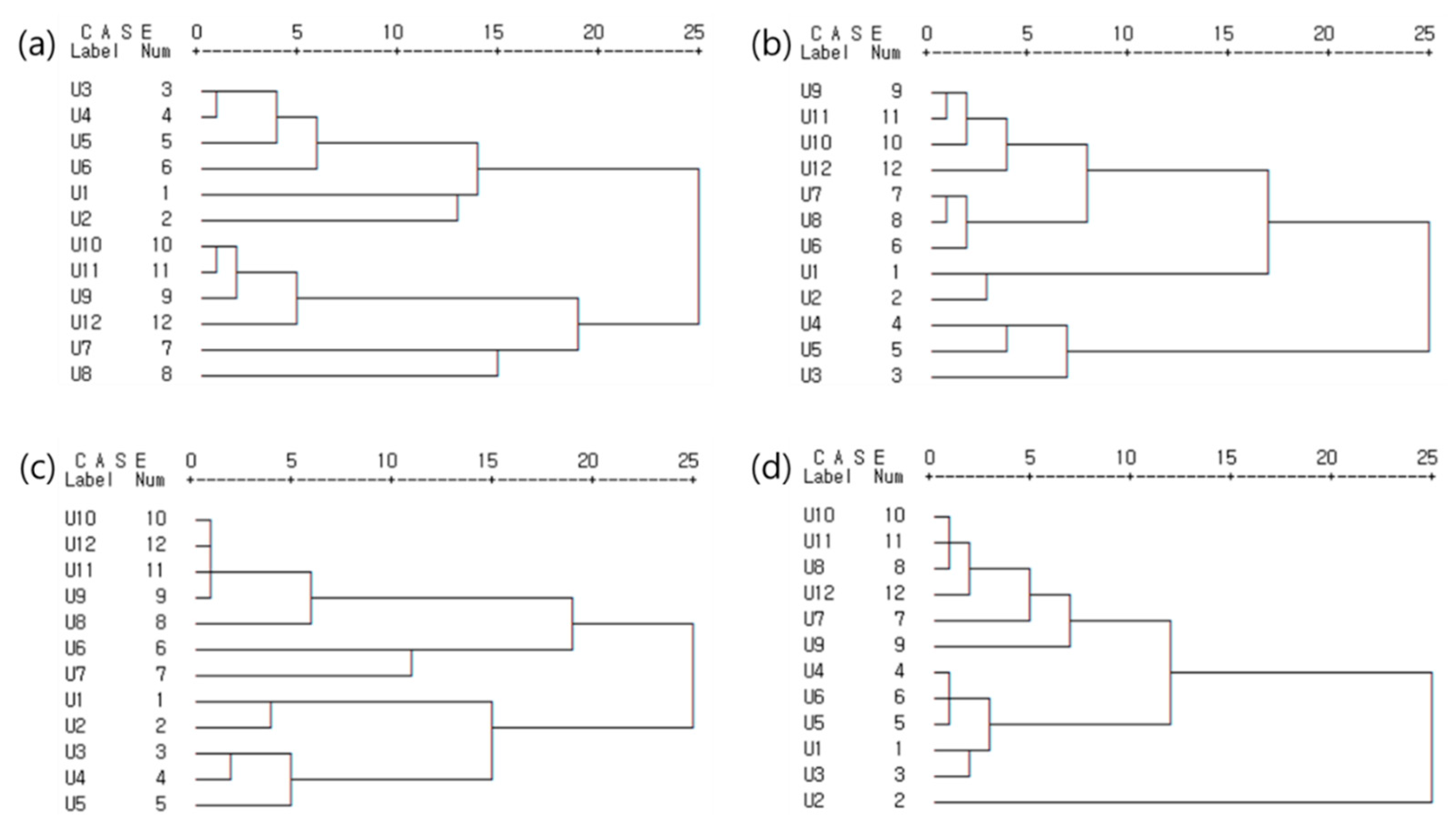
4.5. Estuary Classification
4.6. Causes of Seasonal Algal Blooms
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hallegraeff, G.M. Harmful algal blooms in the Australian region. Mar. Pollut. Bull. 1992, 25, 186–790. [Google Scholar] [CrossRef]
- Tang, D.; Kester, D.R.; Ni, I.-H.; Qi, Y.; Kawamura, H. In situ and satellite observations of a harmful algal bloom and water condition at the Pearl River estuary in late autumn 1998. Harmful Algae 2003, 2, 89–99. [Google Scholar] [CrossRef]
- Anderson, D.M.; Burkholder, J.M.; Cochlan, W.P.; Gilbert, P.M.; Gobler, C.J.; Heil, C.A.; Kudela, R.; Parsons, M.L.; Rensel, J.E.J.; Townsend, D.W.; et al. Harmful algal blooms and eutrophications: Examining linkages from selected coastal regions of the United States. Harmful Algae 2008, 8, 39–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paerl, H.W. Assessing and managing nutrient-enhanced eutrophication in estuarine and coastal waters: Interactive effects of human and climatic perturbations. Ecol. Eng. 2006, 26, 40–54. [Google Scholar] [CrossRef]
- Howarth, R.W.; Bilen, G.; Swaney, D.; Townsend, A.; Jaworski, N.; Lajtha, K.; Oowning, J.A.; Elmgren, R.; Caraco, N.; Jordan, T.; et al. Regional nitrogen budgets and riverine N and P fluxes for the drainages to the North Atlantic Ocean: Natural and human influences. Biogeochemistry 1996, 35, 75–79. [Google Scholar] [CrossRef]
- D’Elia, C.F. Too much of a good thing: Nutrient enrichment of the Chesapeake Bay. Environment 1987, 29, 2. [Google Scholar] [CrossRef]
- National Research Council (NRC). Clean Coastal Waters: Understanding and Reducing the Effects of Nutrient Pollution; National Academy Press: Washington, DC, USA, 2000. [Google Scholar]
- Clesceri, E.J. Sources and Depositional Patterns of Particulate Organic Matter in the Neuse River Estuary, North Carolina (USA). Ph.D. Thesis, University of North Carolina at Chapel Hill, Chapel Hill, NC, USA, February 2004. [Google Scholar]
- Moore, S.K.; Trainer, V.L.; Mantua, N.J.; Parker, M.S.; Laws, E.A.; Backer, L.C.; Fleming, L.E. Impacts of climate variability and future climate change on harmful algal blooms and human health. Environ. Health 2008, 7, S4. [Google Scholar] [CrossRef] [Green Version]
- Rabalais, N.N.; Turner, R.E.; Diaz, R.J.; Justic, D. Global change and eutrophication of coastal waters. ICES J. Mar. Sci. 2009, 66, 1528–1537. [Google Scholar] [CrossRef]
- Cloern, J.E.; Schraga, T.S.; Lopez, C.B.; Knowles, N. Climate anomalies generate an exceptional dinoflagellate bloom in San Francisco Bay. Geophys. Res. Lett. 2005, 32, L14608. [Google Scholar] [CrossRef] [Green Version]
- Jahan, R.; Choi, J.K. Climate regime shift and phytoplankton phenology in a macrotidal estuary: Long-term surveys in Gyeonggi Bay, Korea. Estuaries Coasts 2014, 37, 1169–1187. [Google Scholar] [CrossRef]
- Revstock, G.A.; Kang, Y.S. A comparison of three marine ecosystems surrounding the Korean peninsula: Responses to climate change. Prog. Oceanogr. 2003, 59, 357–379. [Google Scholar] [CrossRef]
- Kim, S.; Zhang, C.-I.; Kim, J.-Y.; Oh, J.-H.; Kang, S.; Lee, J.B. Climate variability and its effects on major fisheries in Korea. Ocean Sci. J. 2007, 42, 179–192. [Google Scholar] [CrossRef]
- Lee, M.O.; Kim, J.K. Characteristics of algal blooms in the southern coastal waters of Korea. Mar. Environ. Res. 2008, 65, 128–147. [Google Scholar] [CrossRef]
- Lee, M.O.; Kim, B.K.; Kwon, Y.; Kim, J.K. Characteristics of the marine environment and algal blooms in Gamak Bay. Fish. Sci. 2009, 75, 410–411. [Google Scholar] [CrossRef]
- Shim, J.M.; Hwang, J.D.; Jeong, C.S.; Lee, Y.H.; Jeon, K.A.; Kwong, K.Y. The influence of oceanic conditions on the occurrence of Cochlodinium polykrikoides blooms in the East Sea. J. Environ. Sci. 2010, 19, 1385–1395. [Google Scholar] [CrossRef]
- Lee, J.; Han, M.-S. Change of blooming pattern and population dynamics of phytoplankton in Masan Bay, Korea. J. Korean Soc. Oceanogr. 2007, 12, 147–158. [Google Scholar]
- Kim, H.G.; Jung, C.S.; Lim, W.A.; Lee, C.K.; Kim, S.Y.; Youn, S.H.; Cho, Y.C.; Lee, S.G. The spatio-temporal progress of Cochlodinium polykrikoides blooms in the coastal waters of Korea. Korean Soc. Fish. Aquat. Sci. 2001, 34, 691–696. [Google Scholar]
- Khim, J.S.; Lee, K.T.; Kannan, K.; Villeneuve, D.L.; Giesy, J.P.; Koh, C.H. Trace organic contaminants in sediment and water from Ulsan Bay and its vicinity, Korea. Arch. Environ. Contam. Toxicol. 2001, 40, 141–150. [Google Scholar]
- Koh, C.H.; Kim, G.B.; Maruya, K.A.; Anderson, J.W.; Jones, J.M.; Kang, S.G. Induction of the P450 reporter gene system bioassay by polycyclic aromatic hydrocarbons in Ulsan Bay (South Korea). Environ. Pollut. 2001, 111, 437–445. [Google Scholar] [CrossRef]
- Ra, K.; Kim, J.K.; Hong, S.H.; Yim, U.H.; Shim, W.J.; Lee, S.Y.; Kim, T.O.; Lim, J.; Kim, E.S.; Kim, K.T. Assessment of pollution and ecological risk of heavy metals in the surface sediments of Ulsan Bay, Korea. Ocean Sci. J. 2014, 49, 279–289. [Google Scholar] [CrossRef]
- Ulsan City. Ulsan Environment Report; Ulsan City Hall: Ulsan, Korea, 2008; Available online: http://www.ulsan.go.kr/rep/ghpaper4 (accessed on 6 April 2020).
- Cho, H.-J.; Yoon, Y.-B.; Kang, H.-S.; Yoon, S.-K. Characteristics of red tide blooms in the lower reaches of Taehwa River. J. Korean Soc. Water Wastewater 2011, 25, 453–462. [Google Scholar] [CrossRef]
- Sohn, E.R.; Park, J.I.; Lee, B.R.; Lee, J.W.; Kim, J.S. Seasonal variation of physic-chemical factors and size-fractionated phytoplankton biomass at Ulsan seaport of East Sea in Korea. Korean J. Microbiol. 2013, 49, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Shin, M.K. Final Report: Research on Algal Bloom in the Taewha River; Ulsan Regional Environmental Technology Development Center: Ulsan, Korea, 2007. [Google Scholar]
- Sim, B.-R.; Kim, H.C.; Kim, C.-S.; Hwang, D.-W.; Park, J.-H.; Cho, Y.-S.; Hong, S.; Lee, W.-C. Spatio-temporal changes of sediment environment in the Taehwa River estuary, Ulsan of Korea. J. Coast. Res. 2018, 85, 41–45. [Google Scholar] [CrossRef]
- MLTMA (Ministry of Land, Transport and Maritime Affairs). Processes of the Korean Standard Methods for Marine Environment; MLTMA: Gwacheon-si, Korea, 2010; p. 493. (In Korean) [Google Scholar]
- Jung, S.W.; Lim, D.I.; Shin, H.H.; Jeong, D.H.; Roh, Y.H. Relationship between physic-chemical factors and chlorophyll-a concentration in surface water of Masan Bay: Bi-daily monitoring data. Korean J. Environ. Biol. 2011, 29, 98–106. [Google Scholar]
- Hecky, R.E.; Kilham, P. Nutrient limitation of phytoplankton in freshwater and marine environments: A review of recent evidence on the effect of enrichment. Limnol. Oceanogr 1998, 33, 796–822. [Google Scholar] [CrossRef] [Green Version]
- Del-Amo, Y.; Pape, O.L.; Treguer, P.; Quequuiner, B.; Menesquen, A.; Aminit, A. Impacts of high nitrate freshwater inputs on macrotidal ecosystems. I. Seasonal evolution of nutrient limitation for the diatom dominated phytoplankton of Bay of Brest (France). Mar. Ecol. Prog. Ser. 1997, 161, 213–224. [Google Scholar] [CrossRef]
- Twomey, L.; Thompson, P. Nutrient limitation of phytoplankton in seasonally open bar-built estuary: Wilson inlet, Western Australia. J. Phycol. 2001, 37, 16–29. [Google Scholar] [CrossRef]
- Hashimoto, T.; Nakano, S. Effect of nutrient limitation on abundance and growth of phytoplankton in a Japanese Pearl farm. Mar. Ecol. Prog. Ser. 2003, 258, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.L.; Liu, Q. Nutrients in the Changjiang River. Environ. Monit. Assess. 2009, 153, 27–44. [Google Scholar] [CrossRef]
- Liu, S.M.; Hong, G.H.; Zhang, J.; Ye, X.W.; Jiang, X.L. Nutrient budgets for large Chinese estuaries. Biogeosciences 2009, 6, 2245–2263. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.K.; Ryt, J.; Yi, Y.; Lee, W.C.; Kang, D.; Lee, C.H.; Hong, S.; Nam, J.; Khim, J.S. Improved water quality in response to pollution control measures at Masan Bay, Korea. Mar. Pollut. Bull. 2012, 64, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Royer, T.V.; Tank, J.L.; David, M.B. Transport and fate of nitrate in headwater agricultural streams in Illinois. J. Environ. Qual. 2004, 33, 1296–1304. [Google Scholar] [CrossRef] [PubMed]
- Rendell, A.R.; Horrobin, T.M.; Jickells, T.D.; Edmunds, H.M.; Brown, J.; Malcolm, S.J. Nutrient cycling in the Great Ouse estuary and its impact on nutrient fluxes to the Wash, England. Estuar. Coast. Shelf Sci. 1997, 45, 653–668. [Google Scholar] [CrossRef]
- Hu, J.; Li, S. Modeling the mass fluxes and transformations of nutrients in the Pearl River Delta, China. J. Mar. Syst. 2009, 78, 146–167. [Google Scholar] [CrossRef]
- Kwon, K.Y. Behavior of Basic Ecological Components Along the Salinity Gradients in Low Turbidity Estuarine Water. Ph.D. Thesis, Pukyong National University, Busan, Korea, 2002. [Google Scholar]
- Lee, J.Y. Consideration of Environmental Factors on Phytoplankton and Dinoflagellate Cyst Distribution of Ulsan Bay in Summer. Master’s Thesis, Pukyong National University, Busan, Korea, 2007. [Google Scholar]
- Moon, C.H.; Choi, H.J. Studies on the environmental characteristics and phytoplankton community in the Nakdong River estuary. J. Korean Soc. Oceanogr. 1991, 26, 144–154. [Google Scholar]
- Yoon, Y.H.; Jeong, D.S. Spatio-temporal distributions of phytoplankton community and it’s variation characteristics in the Ulsan coastal waters. Southern east sea of Korea. J. Korean Soc. Mar. Environ. Energy 2019, 22, 159–171. [Google Scholar] [CrossRef]
- Ministry of Oceans and Fisheries (MOF). Marine Environment Standards; MOF: Sejong, Korea, 2018. [Google Scholar]
- Park, M.-O.; Lee, Y.-W.; Park, J.-K.; Kang, C.-S.; Kim, S.-G.; Kim, S.-S.; Lee, S.M. Evaluation of the seawater quality in the coastal area of Korea in 2013–2017. J. Korean Soc. Mar. Environ. Energy 2019, 22, 47–56. [Google Scholar] [CrossRef]
- Honjo, T.; Shimouse, T.; Hanaika, T.A. Red tide occurred at the Hakozaki fishing port, Hakada Bay, in 1973-The growth process and the chlorophyll content. Bull. Plankton Soc. Jpn. 1978, 25, 7–21. [Google Scholar]
- Curl, H.J.; McLeod, G.C. The physiological ecology of a marine diatom, Skeletonema costutum (Grev.) Cleve. J. Mar. Res. 1961, 19, 70–88. [Google Scholar]
- Türkoglu, M. Temporal variations of surface phytoplankton, nutrients and chlorophyll a in the Dardanelles (Turkish Straits system): A coastal station sample in weekly time intervals. Turk. J. Biol. 2010, 34, 319–333. [Google Scholar]
- Yoon, Y.H.; Rho, H.G.; Kim, Y.K. Seasonal succession of phytoplankton population in the Hamdok port, Northern Cheju Island. Bull. Mar. Cheju Natl. Univ. 1992, 16, 27–42. [Google Scholar]
- Maita, Y.; Odate, T. Seasonal change in size-fractionated primary production and nutrient concentrations in the temperate neritic water of Funka Bay, Japan. J. Oceanogr. Soc. Jpn. 1988, 44, 268–279. [Google Scholar] [CrossRef]
- Yoon, Y.H. Marine Environment and Phytoplankton Community in the Southwestern Sea of Korea; Choi, J.K., Ed.; The Plankton Ecology of Korean Coastal Waters, Donghwa Tech. Publ. Co.: Seoul, Korea, 2011; pp. 68–93. [Google Scholar]
- Guo, C.; Liu, H.; Zheng, L.; Song, S.; Chen, B.; Huang, B. Seasonal and spatial patterns of picophytoplankton growth, grazing and distribution in the East China Sea. Biogeosciences 2014, 11, 1847–1862. [Google Scholar] [CrossRef] [Green Version]
- Iriate, A.; Purdie, D.A. Size distribution of chlorophyll-a biomass and primary production in a temperate estuary (Southampton water)—The contribution of photosynthetic picoplankton. Mar. Ecol. Prog. Ser. 1994, 115, 283–297. [Google Scholar] [CrossRef]
- Lee, M.-J.; Kim, D.S.; Kim, Y.O.; Sohn, M.H.; Moon, C.H.; Baek, S.H. Seasonal phytoplankton growth and distribution pattern by environmental factor changes in inner and outer bay of Ulsan, Korea. J. Korean Soc. Oceanogr. 2016, 21, 24–35. [Google Scholar] [CrossRef]
- Dortch, Q.; Whitledge, T.E. Does nitogen or silcon limit phytoplankton production in the Mississippi River plume and nearby regions. Cont. Shelf Res. 1992, 12, 1293–1309. [Google Scholar] [CrossRef]
- Kim, J.T. Taxonomic and Floristic Accounts of the Green Euglenoids (Euglenophyta) in Korean Fresh Waters. Ph.D. Thesis, Chungnam National University, Daejon, Korea, 1998. [Google Scholar]
- Kim, J.T.; Boo, S.M. The relationships between green Euglenoids and environmental variables in the urban drainage Jeonjucheon in Korea. Koean J. Limnol. 2001, 34, 81–89. [Google Scholar]
- Palmer, C.M. A composite rating of algae tolerating organic pollution. J. Phycol. 1969, 5, 78–82. [Google Scholar] [CrossRef]
- Kim, J.T.; Boo, S.M. Morphological variation and density of Euglena viridis (Euglenophyceae) related to environmental factors in the urban drainages. Korean J. Limnol. 2001, 34, 185–191. [Google Scholar]
- Wang, H.; Hyang, B.; Hong, H. Size-fractionated productivity and nutrient dynamics of phytoplankton in subtropical coastal environments. Hydtobiologia 1997, 352, 97–106. [Google Scholar] [CrossRef]
- Lee, C.K.; Kim, H.C.; Lee, S.-G.; Jung, C.S.; Kim, H.G.; Lim, W.A. Abundance of harmful algae, Cochlodinium catenatum in the coastal area of South sea of Korea and their effects of temperature, salinity, irradiance and nutrient on the growth in culture. J. Korean Fish. Soc. 2001, 34, 536–544. [Google Scholar]
- Adolf, J.E.; Bachvaroff, T.; Place-Allen, R. Can cryptophyte abundance trigger toxic Karlodinium veneficum blooms in eutrophic estuaries. Harmful Algae 2008, 8, 119–128. [Google Scholar] [CrossRef]
- Weng, H.-X.; Qin, Y.-C.; Sun, X.-W.; Dong, H.; Chen, X.-H. Iron and phosphorous effects on the growth of Cryptomonas sp. (Cryptophycea) and their availability in sediments from the Pearl River Estuary, China. Estuar. Coast. Shelf Sci. 2007, 73, 501–509. [Google Scholar] [CrossRef]
- Lee, C.S. Investigation of water quality and hydrological characteristic when red tide develop in the mouth of Hyeongsan River. J. Environ. Sci. 2009, 18, 1155–1162. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| February | May | August | November | |
|---|---|---|---|---|
| Avg. water level (m) | 0.96 | 1.09 | 1.25 | 1.1 |
| Avg. water discharge (m3·s−1) | 6.1 | 11.33 | 12.5 | 6.61 |
| Total precipitation (mm) | 12 | 38.1 | 220.4 | 65.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sim, B.-R.; Kim, H.-C.; Kim, C.-S.; Kim, J.-H.; Park, K.-W.; Lim, W.-A.; Lee, W.-C. Seasonal Distributions of Phytoplankton and Environmental Factors Generate Algal Blooms in the Taehwa River, South Korea. Water 2020, 12, 3329. https://doi.org/10.3390/w12123329
Sim B-R, Kim H-C, Kim C-S, Kim J-H, Park K-W, Lim W-A, Lee W-C. Seasonal Distributions of Phytoplankton and Environmental Factors Generate Algal Blooms in the Taehwa River, South Korea. Water. 2020; 12(12):3329. https://doi.org/10.3390/w12123329
Chicago/Turabian StyleSim, Bo-Ram, Hyung-Chul Kim, Chung-Sook Kim, Jin-Ho Kim, Kyung-Woo Park, Weol-Ae Lim, and Won-Chan Lee. 2020. "Seasonal Distributions of Phytoplankton and Environmental Factors Generate Algal Blooms in the Taehwa River, South Korea" Water 12, no. 12: 3329. https://doi.org/10.3390/w12123329