
Dynamics of Microbial Biomass, Nitrogen Mineralization and Crop Uptake in Response to Placement of Maize Residue Returned to Chinese Mollisols over the Maize Growing Season

Abstract
:1. Introduction
2. Materials and Methods
2.1. Soils and Maize Residue Preparation
2.2. Experiment Design
2.3. Soil Collection and Analysis
2.4. Data Analysis
3. Results
3.1. Soil Microbial Biomass
3.2. Inorganic N Dynamics
3.3. The Relationship between N Uptake and Maize Growth
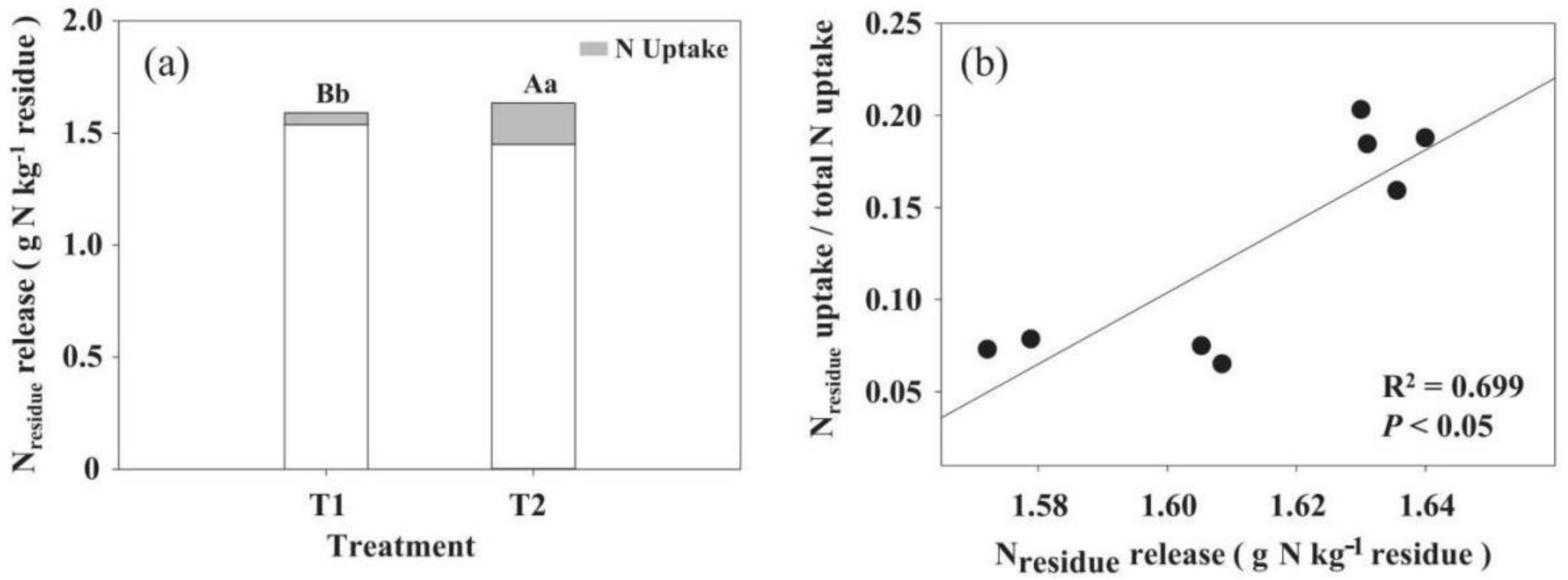
3.4. The Distribution of Residue-Derived N
4. Discussion
4.1. Microbial Biomass
4.2. N Dynamics Dominated by Microbes in the Growing Period
4.3. Maize N Uptake and Growth
4.4. The Distribution of Residue-Derived N in the Soil
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, X.; Xin, X.; Yang, W.; Zhu, A.; Ding, S. Short-term decomposition turnover and retention of residue-derived carbon are influenced by the fertility level in a sandy loam soil. Geoderma 2019, 349, 68–78. [Google Scholar] [CrossRef]
- Liang, A.Z.; McLaughlin, N.B.; Ma, B.L.; Gregorich, E.G.; Morrison, M.J.; Burtt, S.D.; Evenson, L. Changes in mouldboard plough draught and tractor fuel consumption on continuous corn after 18 years of organic and inorganic N amendments. Energy 2013, 52, 89–95. [Google Scholar] [CrossRef]
- Chen, J.M. Carbon neutrality: Towards a sustainable future. Innovation 2021, 2, 100127. [Google Scholar]
- Zhang, J.; Zhang, J.; Shen, G.; Wang, R.; Gao, L.; Li, Z.; Dai, Y.; Meng, G.; Xiang, B.; Zhang, Z. Effects of different types of straw and stalk returning on carbon and nitrogen mineralization in tobacco soil. Tob. Sci. Technol. 2016, 49, 1–6. [Google Scholar]
- Li, Z.; Zhao, B.; Olk, D.C.; Jia, Z.; Mao, J.; Cai, Y.; Zhang, J. Contributions of residue-C and -N to plant growth and soil organic matter pools under planted and unplanted conditions. Soil Biol. Biochem. 2018, 120, 91–104. [Google Scholar] [CrossRef]
- Burgess, M.S.; Mehuys, G.R.; Madramootoo, C.A. Nitrogen dynamics of decomposing corn residue components under three tillage systems. SSSA J. 2002, 66, 1350–1358. [Google Scholar] [CrossRef] [Green Version]
- Hu, G.; Liu, X.; He, H.; Zhang, W.; Xie, H.; Wu, Y. Multi-seasonal nitrogen recoveries from crop residue in soil and crop in a temperate agro-ecosystem. PLoS ONE 2015, 10, e0133437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.H.; Yan, K.; Tang, L.S.; Jia, Z.J.; Li, Y. Change in deep soil microbial communities due to long-term fertilization. Soil Biol. Biochem. 2014, 75, 264–272. [Google Scholar] [CrossRef]
- Cayuela, M.L.; Sinicco, T.; Mondini, C. Mineralization dynamics and biochemical properties during initial decomposition of plant and animal residues in soil. Appl. Soil Ecol. 2009, 41, 118–127. [Google Scholar] [CrossRef]
- Blanco-Canqui, H.; Lal, R.; Trigiano, R.N.; Gray, D.J. Crop residue removal impacts on soil productivity and environmental quality. Crit. Rev. Plant Sci. 2009, 28, 139–163. [Google Scholar] [CrossRef]
- Govaerts, B.; Sayre, K.D.; Ceballos-Ramirez, J.M.; Luna-Guido, M.L.; Limon-Ortega, A.; Deckers, L.; Dendooven, L. Conventionally tilled and permanent raised beds with different crop residue management: Effects on soil C and N dynamics. Plant Soil 2006, 280, 143–155. [Google Scholar] [CrossRef]
- Aulakh, M.A.; Doran, J.W.; Walters, D.T.; Mosier, A.R.; Francis, D.D. Crop residue type and placement effects on denitrification and mineralization. SSSA J. 1991, 55, 1020–1025. [Google Scholar] [CrossRef] [Green Version]
- Jat, R.L.; Jha, P.; Dotaniya, M.L.; Lakaria, B.L.; Rashmi, I.; Meena, B.P.; Meena, A.L. Carbon and nitrogen mineralization in Vertisol as mediated by type and placement method of residue. Environ. Monit. Assess. 2018, 190, 439. [Google Scholar] [CrossRef]
- Curtin, D.; Selles, F.; Wang, H.; Campbell, C.A.; Biederbeck, V.O. Carbon dioxide emissions and transformation of soil carbon and nitrogen during wheat straw decomposition. SSSA J. 1998, 62, 1035–1041. [Google Scholar] [CrossRef]
- Coppens, F.; Garnier, P.; Merckx, R.; Recous, S. Soil moisture carbon and nitrogen dynamics following incorporation versus surface application of labelled residues in soil columns. Eur. J. Soil Sci. 2006, 57, 894–905. [Google Scholar] [CrossRef]
- Urakawa, R.; Shibata, H.; Kuroiwa, M.; Inagaki, Y.; Tateno, R.; Hishi, T.; Suwa, Y. Effects of freeze–thaw cycles resulting from winter climate change on soil nitrogen cycling in ten temperate forest ecosystems throughout the Japanese archipelago. Soil Biol. Biochem. 2014, 74, 82–94. [Google Scholar] [CrossRef]
- Li, X.X.; Hu, C.S.; Delgado, J.A.; Zhang, Y.M.; Ouyang, Z.Y. Increased nitrogen use efficiencies as a key mitigation alternative to reduce nitrate leaching in north China plain. Agric. Water Manag. 2007, 89, 137–147. [Google Scholar] [CrossRef]
- Zhu, D.W.; Huang, Y.; Jin, Z.Q.; Zhang, W.; Jiang, J. Nitrogen management evaluated by models combined with GIS—A case study of Jiangsu croplands China in 2000. Agric. Sci. China 2008, 7, 999–1009. (In Chinese) [Google Scholar] [CrossRef]
- Bowles, T.M.; Atallah, S.S.; Campbell, E.E.; Gaudin, A.C.M.; Wieder, W.R.; Grandy, A.S. Addressing agricultural nitrogen losses in a changing climate. Nat. Sustain. 2018, 1, 399–408. [Google Scholar] [CrossRef]
- Naresh, R.K.; Timsina, J.; Bhaskar, V.; Gupta, R.K.; Singh, A.K.; Dhaliwa, S.S.; Rathore, R.S.; Kumar, V.; Singh, P.; Singh, S.P. Effects of tillage residue and nutrient management on soil organic carbon dynamics and its fractions soil aggregate stability and soil carbon sequestration: A review. EC Nutr. 2017, 12, 53–80. [Google Scholar]
- Sugihara, S.; Funakawa, S.; Kilasara, M.; Kosaki, T. Effect of land management on soil microbial N supply to crop N uptake in a dry tropical cropland in Tanzania. Agric. Ecosyst. Environ. 2012, 146, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Bird, J.A.; Horwath, W.R.; Eagle, A.J. Immobilization of fertilizer nitrogen in rice. SSSA J. 2001, 65, 1143–1152. [Google Scholar] [CrossRef]
- Sakala, W.D.; Cadisch, G.; Giller, K.E. Interactions between residues of maize and pigeon pea and mineral N fertilizers during decomposition and N mineralization. Soil Biol. Biochem. 2000, 32, 679–688. [Google Scholar] [CrossRef]
- Pan, F.; Yu, W.; Ma, Q.; Zhou, H.; Jiang, C.; Xu, Y. Do organic amendments improve the synchronism between soil N supply and wheat demand? Appl. Soil Ecol. 2018, 125, 184–191. [Google Scholar] [CrossRef]
- Brookes, P.C.; Kragt, J.F.; Powlson, D.S.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: The effects of fumigation time and temperature. Soil Biol. Biochem. 1985, 17, 831–835. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Joergensen, R.G.; Mueller, T. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the ken value. Soil Biol. Biochem. 1996, 28, 33–37. [Google Scholar] [CrossRef]
- Chen, Y.; Tang, X.; Yang, S.; Wu, C.; Wang, J. Contributions of different N sources to crop N nutrition in a Chinese rice field. Pedosphere 2010, 20, 198–208. [Google Scholar] [CrossRef]
- Zhao, S.; Li, K.; Zhou, W.; Qiu, S.; Huang, S.; He, P. Changes in soil microbial community enzyme activities and organic matter fractions under long–term straw return in north–central China. Agric. Ecosyst. Environ. 2016, 216, 82–88. [Google Scholar] [CrossRef]
- Heijboer, A.; ten Berge, H.F.; de Ruiter, P.C.; Jørgensen, H.B.; Kowalchuk, G.A.; Bloem, J. Plant biomass soil microbial community structure and nitrogen cycling under different organic amendment regimes: A 15N tracer–based approach. Appl. Soil Ecol. 2016, 107, 251–260. [Google Scholar] [CrossRef]
- Lou, Y.; Liang, W.; Xu, M.; He, X.; Wang, Y.; Zhao, K. Straw coverage alleviates seasonal variability of the topsoil microbial biomass and activity. Catena 2011, 86, 117–120. [Google Scholar] [CrossRef]
- Sun, B.J.; Jia, S.X.; Zhang, S.X.; McLaughlin, N.B.; Zhang, X.; Liang, A.Z.; Liu, S.Y. Tillage seasonal and depths effects on soil microbial properties in black soil of Northeast China. Soil Till. Res. 2016, 155, 421–428. [Google Scholar] [CrossRef]
- Liang, A.Z.; Zhang, Y.; Zhang, X.P.; Yang, X.M.; McLaughlin, N.B.; Chen, X.W.; Tang, J. Investigations of relationships among aggregate pore structure microbial biomass and soil organic carbon in a Mollisol using combined non-destructive measurements and phospholipid fatty acid analysis. Soil Till. Res. 2019, 185, 94–101. [Google Scholar] [CrossRef]
- Fontaine, S.; Barot, S.; Barré, P.; Bdioui, N.; Mary, B.; Rumpel, C. Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature 2007, 450, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Mooshammer, M.; Wanek, W.; Hammerle, I.; Fuchslueger, L.; Hofhansl, F.; Knoltsch, A.; Schnecker, J.; Takriti, M.; Watzka, M.; Wild, B.; et al. Adjustment of microbial nitrogen use efficiency to carbon:nitrogen imbalances regulates soil nitrogen cycling. Nat. Commun. 2014, 5, 3694. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, Y.Y.; Yang, S.; Wang, Z.R.; Feng, X.; Liu, H.Y.; Jiang, Y. Variations in soil bacterial taxonomic profiles and putative functions in response to straw incorporation combined with N fertilization during the maize growing season. Agric. Ecosyst. Environ. 2019, 283, 106578. [Google Scholar] [CrossRef]
- Rumpel, C.; Kogel-Knabner, I. Deep soil organic matter-a key but poorly understood component of terrestrial C cycle. Plant Soil 2011, 338, 143–158. [Google Scholar] [CrossRef]
- Jobbagy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Recous, S.; Robin, D.; Darwis, D.; Mary, B. Soil inorganic N availability: Effect on maize residue decomposition. Soil Biol. Biochem. 1995, 27, 1529–1538. [Google Scholar] [CrossRef]
- Corbeels, M.; Hofman, G.; Cleemput, O.V. Nitrogen cycling associated with the decomposition of sunflower stalks and wheat straw in a Vertisol. Plant Soil 2000, 218, 71–82. [Google Scholar] [CrossRef]
- Said-Pullicino, D.; Cucu, M.A.; Sodano, M.; Birk, J.J.; Glaser, B.; Celi, L. Nitrogen immobilization in paddy soils as affected by redox conditions and rice straw incorporation. Geoderma 2014, 228–229, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Palm, C.A.; Gachengo, C.; Delve, R.; Cadisch, G.; Giller, K.E. Organic inputs for soil fertility management in tropical agroecosystems: Application of an organic resource database. Agric. Ecosyst. Environ. 2001, 83, 27–42. [Google Scholar] [CrossRef]
- Geisseler, D.; Horwath, W.R.; Joergensen, R.G.; Ludwig, B. Pathways of nitrogen utilization by soil microorganisms—A review. Soil Biol. Biochem. 2010, 42, 2058–2067. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Xu, X. Tansley Review: Competition between roots and microorganisms for N: Mechanisms and ecological relevance. New Phytol. 2013, 198, 656–669. [Google Scholar] [CrossRef]
- Liu, C.; Lu, M.; Cui, J.; Li, B.; Fang, C. Effects of straw carbon input on carbon dynamics in agricultural soils: A meta-analysis. GCB. Bioenergy 2014, 20, 1366–1381. [Google Scholar] [CrossRef]
- Wang, M.; Pendall, E.; Fang, C.; Li, B.; Nie, M. A global perspective on agroecosystem nitrogen cycles after returning crop residue. Agric. Ecosyst. Environ. 2018, 266, 49–54. [Google Scholar] [CrossRef]
- Soon, Y.K.; Lupwayi, N.Z. Straw management in a cold semi-arid region: Impact on soil quality and crop productivity. Field Crops Res. 2012, 139, 39–46. [Google Scholar] [CrossRef]
- Freeman, K.W.; Girma, K.; Arnall, D.B.; Mullen, R.W.; Martin, K.L.; Teal, R.K. By-plant prediction of corn forage biomass and nitrogen uptake at various growth stages using remote sensing and plant height. Agron. J. 2007, 99, 530–536. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Niu, J.; Peng, Z.; Zhang, F.; Li, C. Shoot growth potential drives N uptake in maize plants and correlates with root growth in the soil. Field Crop. Res. 2010, 115, 85–93. [Google Scholar] [CrossRef]
- Ding, W.; Li, S.; He, P.; Huang, S. Contribution and fate of maize residue-15N and urea-15N as affected by N fertilization regime. PLoS ONE 2019, 14, e0210176. [Google Scholar] [CrossRef]
- Shan, H.X.; Lu, C.A.; Zhang, J.T.; Wang, J.Z.; Xu, M.G. Effect of maize straw applied with N fertilizer on nitrogen adsorption of winter wheat under different soil fertility. J. Plant Nutr. Fert. 2012, 18, 35–41. (In Chinese) [Google Scholar]
- Cotrufo, W.F.; Wallenstein, M.D.; Boot, C.M.; Paul, E. The Microbial Efficiency-Matrix Stabilization framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Chang. Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SOC | TN | pH | SWC | NO3− | NH4+ | MBN | |
|---|---|---|---|---|---|---|---|
| g kg−1 | g kg−1 | -- | % | mg N kg−1 | mg N kg−1 | mg N kg−1 | |
| Value | 11.64 (0.06) | 1.10 (0.02) | 7.01 (0.03) | 22.30 (0.87) | 13.10 (0.27) | 9.22 (0.09) | 40.86 (2.00) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Liang, A.; Zhang, Y.; McLaughlin, N.; Zhang, S.; Chen, X.; Zheng, H.; Fan, R. Dynamics of Microbial Biomass, Nitrogen Mineralization and Crop Uptake in Response to Placement of Maize Residue Returned to Chinese Mollisols over the Maize Growing Season. Atmosphere 2021, 12, 1166. https://doi.org/10.3390/atmos12091166
Gao Y, Liang A, Zhang Y, McLaughlin N, Zhang S, Chen X, Zheng H, Fan R. Dynamics of Microbial Biomass, Nitrogen Mineralization and Crop Uptake in Response to Placement of Maize Residue Returned to Chinese Mollisols over the Maize Growing Season. Atmosphere. 2021; 12(9):1166. https://doi.org/10.3390/atmos12091166
Chicago/Turabian StyleGao, Yan, Aizhen Liang, Yan Zhang, Neil McLaughlin, Shixiu Zhang, Xuewen Chen, Haifeng Zheng, and Ruqin Fan. 2021. "Dynamics of Microbial Biomass, Nitrogen Mineralization and Crop Uptake in Response to Placement of Maize Residue Returned to Chinese Mollisols over the Maize Growing Season" Atmosphere 12, no. 9: 1166. https://doi.org/10.3390/atmos12091166