
Complete Chloroplast Genome of Krascheninnikovia ewersmanniana: Comparative and Phylogenetic Analysis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Genomic DNA Extraction and Sequencing
2.3. Chloroplast Genome Assembly and Annotation
2.4. Analysis of Chloroplast Genome Repeats and Codon Usage Preferences
2.5. Comparison of Chloroplast Genomes
2.6. Phylogenetic Analysis
3. Results
3.1. Genomic Features of K. ewersmanniana
3.2. Analysis of the Codon Usage Profiles
3.3. Repeat Analysis
3.4. Comparison of the Chloroplast Genomes of Seven Amaranthaceae Plants
3.5. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Editorial Committee of Chinese Journal of Plant of Chinese Academy of Sciences. Flora China Tomus; Science Press: Beijing, China, 1996; Volume 25, pp. 26–27. [Google Scholar]
- Ke, M.; Hou, Y.R.; Wei, P.; Lan, J.Y.; Kang, S.; Li, C. Physiological responses of Krascheninnikovia ewersmannia under drought stress. Pratacultural. Sci. 2023, 40, 1349–1357. [Google Scholar]
- Cai, D.H.; Li, C.J.; Wei, Y. Germination behavior of Cerateoides latens seed in Junggar desert. Acta Agrestia Sin. 2016, 24, 384–388. [Google Scholar]
- Khan, M.A.; Gul, B.; Weber, D.J. Action of plant growth regulators and salinity on seed germination of Ceratoides lanata. Anim. Husb. Feed Sci. 2012, 82, 37. [Google Scholar] [CrossRef]
- Mao, Z.M. Flora Xinjiang Gensis Tomus; Xinjiang Science & Technology & Hygiene Publishing House: Urumqi, China, 1994; p. 2 (I). [Google Scholar]
- Wang, M.R.; Wei, Y. Germination behavior of Ceratoides ewersmanniana seeds in northern Xinjiang. Pratacultural. Sci. 2018, 35, 1836–1842. [Google Scholar]
- Chu, G.L.; Sergei, L.M.; Steven, E.C. Chenopodiaceae. Flora of China; Science Press: Beijing, China, 2003; Volume 5, pp. 351–414. [Google Scholar]
- Jia, M. Ceratoides latens resources and utilization indesert steppe. Anim. Husb. Feed Sci. 2012, 33, 123–124. [Google Scholar]
- Tao, Y.; Liu, T.; Jia, Y.M.; Cui, Y.H.; Luo, C.; Wei, P.; Jiangbo, X. Fractal characteristics of spatial distribution pattern of Ceratoides ewersmanniana and Haloxylon ammodendron population in the southern Gurbantunggut desert, Xinjiang. Arid Land Geogr 2008, 31, 365–372. [Google Scholar]
- Liu, T.; Li, Z.; An, S.Z.; Xu, G.Y. A study on root system of Ceratoides ewersmanniana. Grassl. Turf 2008, 1, 13–17. [Google Scholar]
- Jia, Y.M. Research on the Bioecological Characteristics of Spatial Distribution Pattern of Ceratoides ewersmanniana in the Southern Edge of Mosuowan Desert; Shihezi University: Shihezi, China, 2007. [Google Scholar]
- Li, L.; Jia, N.T.; Liu, M. Study on the physiological and biochemical index of drought resistance for Ceratoides Ewersmanniana ‘Yili’. Grass Feed Livest 2013, 1, 41–45. [Google Scholar]
- Zheng, X.H.; Li, Z.; Fu, A.L.; Yang, G.; Sha, W.L. Study in regenerated plants induced from embryo of Ceratoides ewersmanniana. China J. Grassl. 2007, 4, 50–55. [Google Scholar]
- Li, D.L.; Wang, J.H.; Li, A.D.; Liu, H.J.; An, F.B. Salt tolerance of 3 Ceratoides pps varieties in germination period. J. Desert Res. 2006, 26, 1009–1014. [Google Scholar]
- Mohajery, A.; Rasti, M. Germination temperature for Bromus tomentellus Boiss and Eurotia ceratoides (L.) C.A.M. Seed. Sci. Technol. 1995, 23, 241–243. [Google Scholar]
- Aranda-Sicilia, M.N.; Aboukila, A.; Armbruster, U.; Cagnac, O.; Schumann, T.; Kunz, H.H.; Jahns, P.; Rodríguez-Rosales, M.P.; Sze, H.; Venema, K. Envelope K+/H+ antiporters AtKEA1 and AtKEA2 function in plastid development. Plant Physiol. 2016, 172, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Freyer, R.; Hoch, B.; Neckermann, K.; Maier, R.M.; Kössel, H. RNA editing in maize chloroplasts is a processing step independent of splicing and cleavage to monocistronic mRNAs. Plant J. 1993, 4, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, H.E.; Emes, M.J. Nonphotosynthetic metabolism in plastids. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 111–140. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Li, D.Z. Advances in phylogenomics based on complete chloroplast genomes. Plant Divers Res. 2011, 33, 365–375. [Google Scholar] [CrossRef]
- Chen, C.H.; Zheng, Y.J.; Liu, S.A.; Zhong, Y.; Wu, Y.; Li, J.; Xu, L.A.; Xu, M. The complete chloroplast genome of Cinnamomum camphora and its comparison with related Lauraceae species. Peer J. 2017, 5, e3820. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H. Chloroplast Genome Sequences of Vitis: Structural Organization and Phylogenetic Analyses; Huazhong Agricultural University: Wuhan, China, 2018. [Google Scholar]
- Michał, M.; Alžběta, N.; Minasiewicz, J.; Selosse, M.A.; Jakalski, M. The complete chloroplast genome sequence of Dactylorhiza majalis (Rchb.) P.F. Hunt et Summerh. (Orchidaceae). Mitochondrial DNA 2019, 4, 2821–2823. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; He, N.; Li, Y.; Fang, Y.; Zhang, F. Complete chloroplast genome sequence of Chinese lacquer Tree (Toxicodendron vernicifluum, Anacardiaceae) and its phylogenetic significance. BioMed Res. Int. 2020, 2020, 9014873. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Wang, X.M.; Yin, J.; Zhao, M.L.; Yang, Y.Y. Genetic diversity of Ceratoides germplasm by RAPD analysis. Acta Agrestia Sin. 2003, 2, 154–158. [Google Scholar] [CrossRef]
- Liu, Y.; Zheng, C.; Su, X.; Chen, J.; Li, X.; Sun, C.; Nizamani, M.M. Comparative analysis and characterization of the chloroplast genome of Krascheninnikovia ceratoides (Amarathaceae): A xerophytic semi-shrub exhibiting drought resistance and high-quality traits. BMC Genom. Data 2024, 25, 10. [Google Scholar] [CrossRef] [PubMed]
- Bremer, B.; Bremer, K.; Chase, M.W.; Fay, M.F.; Reveal, J.L.; Soltis, D.E.; Soltis, P.S.; Stevens, P.F.; Anderberg, A.A.; Moore, M.J.; et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar] [CrossRef]
- Liang, C.Z.; Zhu, Z.Y.; Tian, Y.; Wang, W. The correction for scientific name of Ceratoides and Prinsepia in the second edition of (Flora Intramongolica). J. Inn. Mong. Univ. (Nat. Sci. Ed.) 2003, 5, 503–505. [Google Scholar]
- Schubert, M.; Lindgreen, S.; Orlando, L. AdapterRemoval v2: Rapid adapter trimming, identification, and read merging. BMC Res. Notes 2016, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.J.; Yu, W.B.; Yang, J.B.; Song, Y.; dePamphilis, C.W.; Yi, T.S.; Li, D.Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.J.; Moore, M.J.; Li, D.Z.; Yi, T.S. PGA: A software package for rapid, accurate, and flexible batch annotation of plastomes. Plant Methods 2019, 15, 50. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Greiner, S.; Lehwark, P.; OrganellarGenomeDRAW, B.R. (OGDRAW) version 1.3. 1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef] [PubMed]
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Mega, K.S. Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Mazumdar, P.; Binti, O.R.; Mebus, K.; Ramakrishnan, N.; Ann, H.J. Codon usage and codon pair patterns in non-grass monocot genomes. Ann. Bot. 2017, 120, 893–909. [Google Scholar] [CrossRef] [PubMed]
- Amiryousefi, A.; Hyvönen, J.; Poczai, P. IRscope: An online program to visualize the junction sites of chloroplast genomes. Bioinformatics 2018, 34, 3030–3031. [Google Scholar] [CrossRef] [PubMed]
- Mayor, C.; Brudno, M.; Schwartz, J.R.; Poliakov, A.; Rubin, E.M.; Frazer, K.A.; Pachter, L.S.; Dubchak, I. VISTA: Visualizing global DNA sequence alignments of arbitrary length. Bioinformatics 2020, 16, 1046–1047. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhao, X.J.; Jia, Y.M.; Cui, Y.H.; Luo, C.; Wei, P.; Zhang, Y.H.; Lin, H.R. Spatial pattern of population recruitment of C. ewersmanniana in South of Gurbantunggut Desert. J. Desert Res. 2008, 28, 258–265. [Google Scholar]
- Shawulie, S.B.H.; An, S.Z.; Yang, G.; Nuerjialili, N.; Zhang, H.H.; Fu, A.L.; Zheng, X.H. Study on the Botanical-biological and ecological characteristics of Ceratoides ewersmanniana cv. YiLi. Xinjiang Agric. Sci. 2010, 47, 1842–1846. [Google Scholar]
- Deng, Y.B.; Jiang, Y.C.; Liu, J. The xeromorphic and saline morphic structure of leaves and assimilating branches in ten Chenopodiaceae species in Xinjiang. Chin. J. Plant Ecol. 1998, 22, 69–75. [Google Scholar]
- Park, J.; Min, J.; Kim, Y.; Chung, Y. The comparative analyses of six complete chloroplast genomes of morphologically diverse Chenopodium album L. (Amaranthaceae) collected in Korea. Int. J. Genom. 2021, 2021, 6643444. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Chen, F.; Ding, H.; Liu, W.; Yang, B.; Geng, J.; Chen, S.; Guo, S. Comparative analysis of six chloroplast genomes in Chenopodium and its related genera (Amaranthaceae): New insights into phylogenetic relationships and the development of species-specific molecular markers. Genes 2023, 14, 2183. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zuo, Z.; He, Y.; Miao, J.; Yu, J.; Qu, C. The complete chloroplast genome of a halophyte glasswort Salicornia europaea. Mitochondrial DNA B Resour. 2023, 8, 1165–1168. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Zhang, C.; Wang, Y.; Zhang, Y. The complete chloroplast genome of Caroxylon passerinum (Chenopodiaceae), an annual desert plant. Mitochondrial DNA B Resour. 2022, 7, 426–427. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.Z.; Dong, Y.H.; Valcárcel, V.; Ren, Z.M.; Li, Y.L. Complete chloroplast genome of the grain Chenopodium quinoa Willed., an important economical and dietary plant. Mitochondrial DNA B Resour. 2021, 6, 40–42. [Google Scholar] [CrossRef]
- Wariss, H.M.; Qu, X.J. The complete chloroplast genome of Chenopodium acuminatum Willd. (Amaranthaceae). Mitochondrial DNA B Resour. 2021, 6, 174–175. [Google Scholar] [CrossRef]
- Li, F.Z.; Zhang, X.L.; Zhu, L.L.; Lu, H.F.; Song, D.L. Characterization of the complete chloroplast genome of an annual herb, Chenopodium album (Amaranthaceae). Mitochondrial DNA B Resour. 2021, 6, 2107–2108. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zheng, Y. Dynamic evolution and phylogenomic analysis of the chloroplast genome in Schisandraceae. Sci. Rep. 2018, 8, 9285. [Google Scholar] [CrossRef] [PubMed]
- Bao, G.Y.; Wang, Y.Q.; Li, W.X. Comparative analysis of chloroplast genome repeats of betavulgaris and its related groups. Seed 2023, 42, 36–44. [Google Scholar]
- Yang, T.; Aishan, S.; Zhu, J.; Qin, Y.; Liu, J.; Liu, H.; Tie, J.; Wang, J.; Qin, R. Chloroplast genomes and phylogenetic analysis of three Carthamus (Asteraceae) species. Int. J. Mol. Sci. 2023, 24, 15634. [Google Scholar] [CrossRef]
- Saha, D.; Rana, R.S.; Das, S.; Datta, S.; Mitra, J.; Cloutier, S.J.; You, F.M. Genome-wide regulatory gene-derived SSRs reveal genetic differentiation and population structure in fiber flax genotypes. J. Appl. Genet. 2019, 60, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; de Waard, J.R. Biological identifications through DNA barcodes. Proc. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Hajibabaei, M.; Singer, G.A.; Hebert, P.D.; Hickey, D.A. DNA barcoding: How it complements taxonomy, molecular phylogenetics and population genetics. Trends Genet. 2007, 23, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Song, J.Y.; Ma, X.Y.; Liu, C.; Li, Y.; Xu, H.X.; Han, J.P.; Duan, L.S.; Chen, S.L. Identification of Dendrobium species by a candidate DNA barcode sequence: The chloroplast psbA-trnH intergenic region. Planta Med. 2009, 75, 667–669. [Google Scholar] [CrossRef]
- Fuentes-Bazan, S.; Uotila, P.; Borsch, T. A novel phylogeny-based generic classification for Chenopodium sensu lato, and a tribal rearrangement of Chenopodioideae (Chenopodiaceae). Willdenowia 2012, 42, 5–24. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
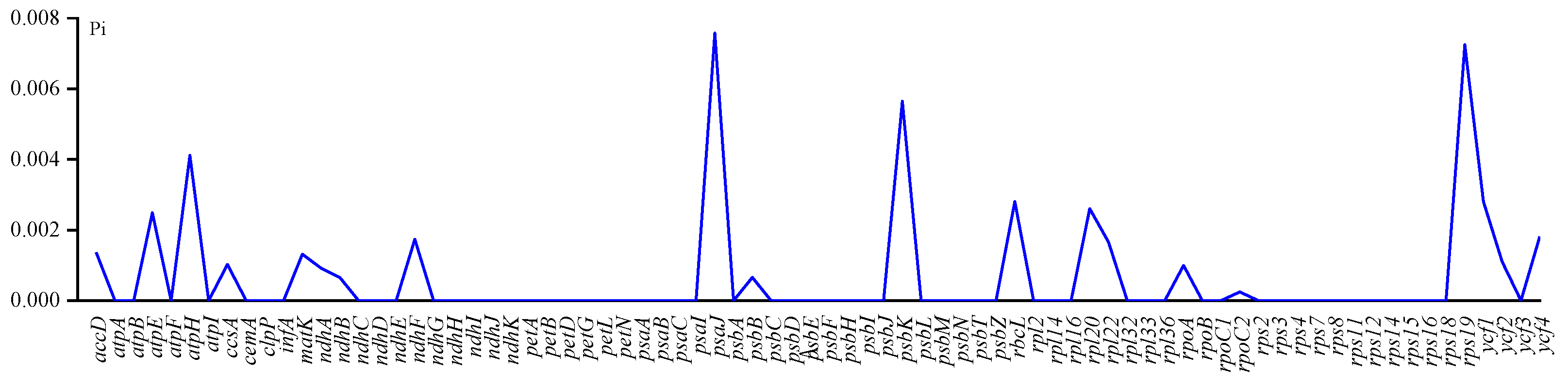{kind=link}
{kind=link}
| Gene Category | Gene Group | Gene Name |
|---|---|---|
| Photosynthesis | Photosystem II | psbZ, psbA, psbT, psbB, psbN, psbC, psbM, psbD, psbL, psbE, psbJ, psbK, psbF, psbI, psbH |
| Photosystem I | psaJ, psaB, psaI, psaC, psaA | |
| Rubisco | rbcL | |
| Cytochrome b/f complex | petD *, petB *, petL, petA, petN, petG | |
| ATP synthase | atpF *, atpI, atpA, atpE, atpB, atpH | |
| NADH-dehydrogenase | ndhB *(2), ndhA *, ndhI, ndhD, ndhC, ndhE, ndhG, ndhF ndhH, ndhK, ndhJ | |
| Self-replication | rRNA genes | rrn4.5S(2), rrn23S(2), rrn5S(2), rrn16S(2) |
| Large subunit of ribosome | rpl16 *, rpl14, rpl2(2), rpl22, rpl36, rpl20, rpl33, rpl32 | |
| DNA-dependent RNA polymerase | rpoC1 *, rpoB, rpoC2, rpoA | |
| tRNA genes | trnA-UGC *(2), trnI-GAU *(2), trnV-UAC *, trnL-UAA *, trnR-UCU *, trnK-UUU *, trnI-CAU(2), trnfM-CAU, trnL-CAA(2), trnW-CCA, trnN-GUU(2), trnT-GGU, trnS-GGA, trnR-ACG(2), trnS-UGA, trnV-GAC(2), trnC-GCA, trnS-GCU, trnD-GUC, trnQ-UUG, trnY-GUA, trnT-UGU, trnM-CAU, trnP-UGG, trnL-UAG, trnF-GAA, trnG-UCC, trnG-GCC, trnE-UUC | |
| Small subunit of ribosome | rps12 *(2), rps16 *, rps8, rps4, rps19, rps14, rps7(2), rps15, rps11, rps2, rps18, rps3, rps12 | |
| Other genes | Translation initiation factor | infA |
| C-type cytochrome synthesis gene | ccsA | |
| Protease | clpP ** | |
| Acetyl-CoA carboxylase | accD | |
| Maturase | matK | |
| Envelope membrane protein | cemA | |
| Unknown | Conserved open reading frames | ycf3 **, ycf2(2), ycf4, ycf1 |
| Gene | Strand | Start | End | Exon I (bp) | Intron I (bp) | Exon II (bp) | Intron II (bp) | Exon III (bp) |
|---|---|---|---|---|---|---|---|---|
| ycf3 | − | 41,858 | 43,885 | 124 | 779 | 230 | 805 | 90 |
| clpP | − | 69,732 | 71,733 | 71 | 811 | 292 | 603 | 225 |
| trnK-UUU | − | 1470 | 4034 | 37 | 2493 | 35 | ||
| rpl16 | − | 80,996 | 82,458 | 9 | 1055 | 399 | ||
| ndhA | + | 118,491 | 120,536 | 552 | 954 | 540 | ||
| petB | + | 74,655 | 76,137 | 6 | 835 | 642 | ||
| rps12 | − | 139,808 | 140,608 | 232 | 543 | 26 | ||
| ndhB | + | 92,771 | 94,970 | 777 | 667 | 756 | ||
| rps12 | + | 95,778 | 96,578 | 232 | 543 | 26 | ||
| trnA-UGC | − | 134,254 | 135,153 | 37 | 827 | 36+ | ||
| ndhB | − | 141,416 | 143,615 | 777 | 667 | 756 | ||
| petD | + | 76,359 | 77,560 | 8 | 719 | 475 | ||
| trnI-GAU | − | 135,224 | 136,227 | 37 | 932 | 35 | ||
| rps16 | − | 4713 | 5913 | 100 | 907 | 194 | ||
| trnV-UAC | − | 51,060 | 51,751 | 38 | 616 | 38 | ||
| trnI-GAU | + | 100,159 | 101,162 | 37 | 932 | 35 | ||
| atpF | − | 11,200 | 12,593 | 145 | 839 | 410 | ||
| trnL-UAA | + | 46,709 | 47,436 | 39 | 639 | 50 | ||
| trnA-UGC | + | 101,233 | 102,132 | 38 | 827 | 35 |
| Codon | Count | Codon | Count | Codon | Count | Codon | Count |
|---|---|---|---|---|---|---|---|
| UAA | 40 | GGC | 168 | AUG | 501 | AGU | 353 |
| UAG | 19 | GGG | 280 | AAC | 253 | UCA | 340 |
| UGA | 19 | GGU | 492 | AAU | 796 | UCC | 224 |
| GCA | 363 | CAC | 132 | CCA | 265 | UCG | 157 |
| GCC | 211 | CAU | 392 | CCC | 169 | UCU | 472 |
| GCG | 160 | AUA | 626 | CCG | 140 | ACA | 381 |
| GCU | 544 | AUC | 320 | CCU | 374 | ACC | 205 |
| UGC | 63 | AUU | 982 | CAA | 655 | ACG | 124 |
| UGU | 190 | AAA | 978 | CAG | 162 | ACU | 476 |
| GAC | 181 | AAG | 280 | AGA | 338 | GUA | 462 |
| GAU | 736 | CUA | 344 | AGG | 141 | GUC | 140 |
| GAA | 1033 | CUC | 144 | CGA | 331 | GUG | 160 |
| GAG | 260 | CUG | 128 | CGC | 88 | GUU | 486 |
| UUC | 409 | CUU | 490 | CGG | 99 | UGG | 381 |
| UUU | 933 | UUA | 827 | CGU | 300 | UAC | 155 |
| GGA | 598 | UUG | 462 | AGC | 104 | UAU | 660 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, P.; Li, Y.; Ke, M.; Hou, Y.; Aikebaier, A.; Wu, Z. Complete Chloroplast Genome of Krascheninnikovia ewersmanniana: Comparative and Phylogenetic Analysis. Genes 2024, 15, 546. https://doi.org/10.3390/genes15050546
Wei P, Li Y, Ke M, Hou Y, Aikebaier A, Wu Z. Complete Chloroplast Genome of Krascheninnikovia ewersmanniana: Comparative and Phylogenetic Analysis. Genes. 2024; 15(5):546. https://doi.org/10.3390/genes15050546
Chicago/Turabian StyleWei, Peng, Youzheng Li, Mei Ke, Yurong Hou, Abudureyimu Aikebaier, and Zinian Wu. 2024. "Complete Chloroplast Genome of Krascheninnikovia ewersmanniana: Comparative and Phylogenetic Analysis" Genes 15, no. 5: 546. https://doi.org/10.3390/genes15050546