
Myelin-Specific microRNA-23a/b Cluster Deletion Inhibits Myelination in the Central Nervous System during Postnatal Growth and Aging

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Genotyping
2.3. Real-Time PCR
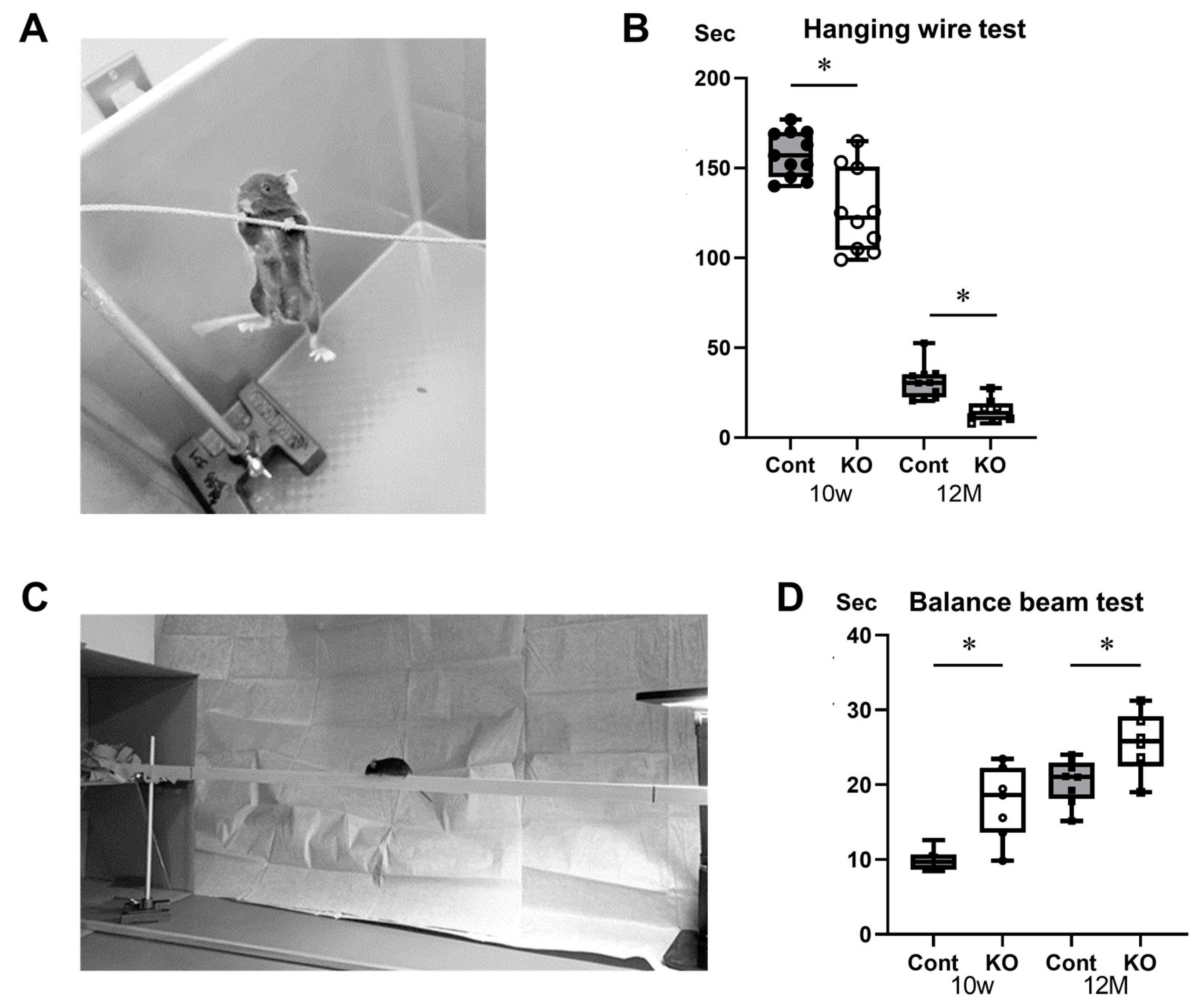
2.4. Behavioral Assessments
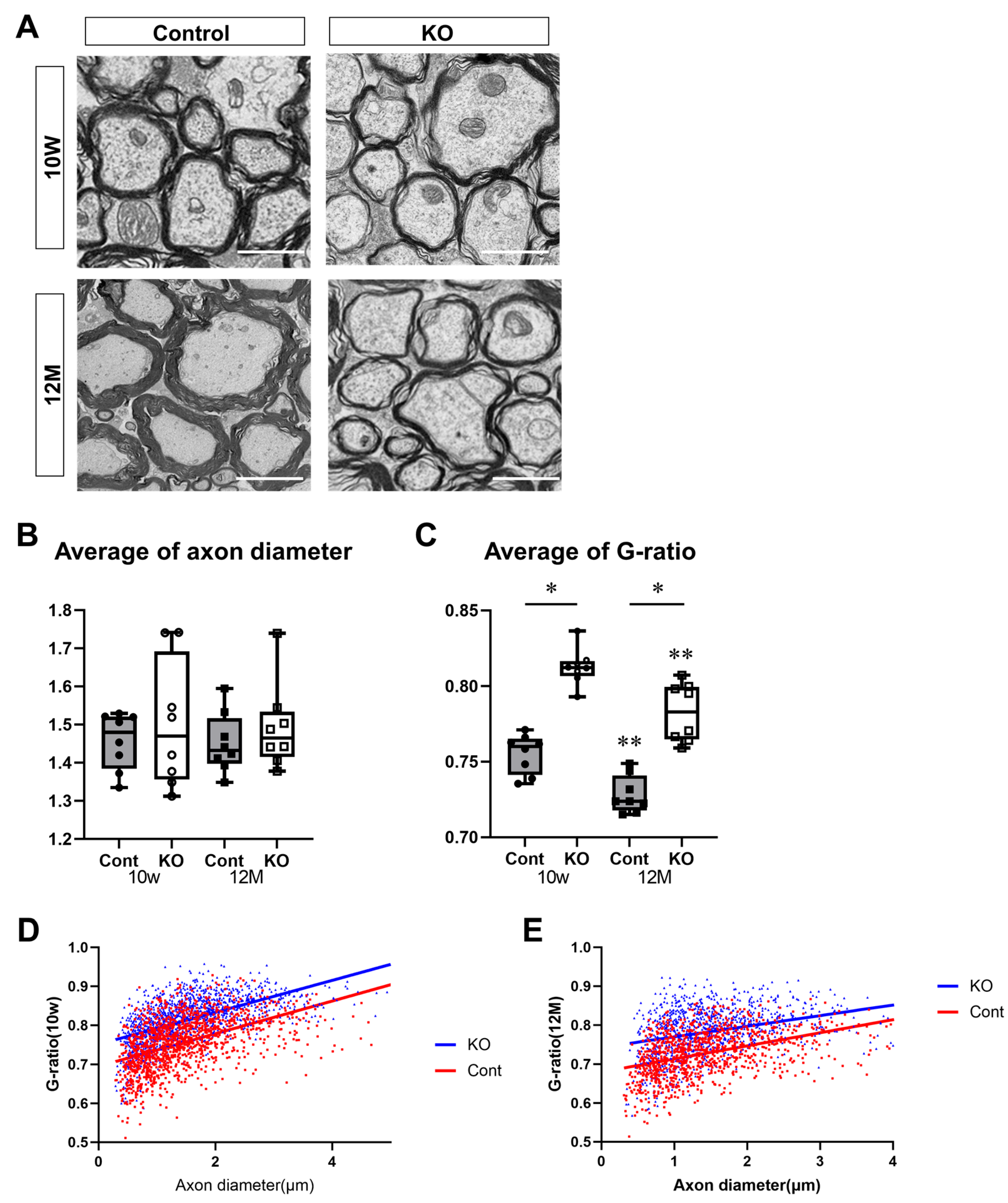
2.5. Electron Microscopy
2.6. Luxol Fast Blue Staining
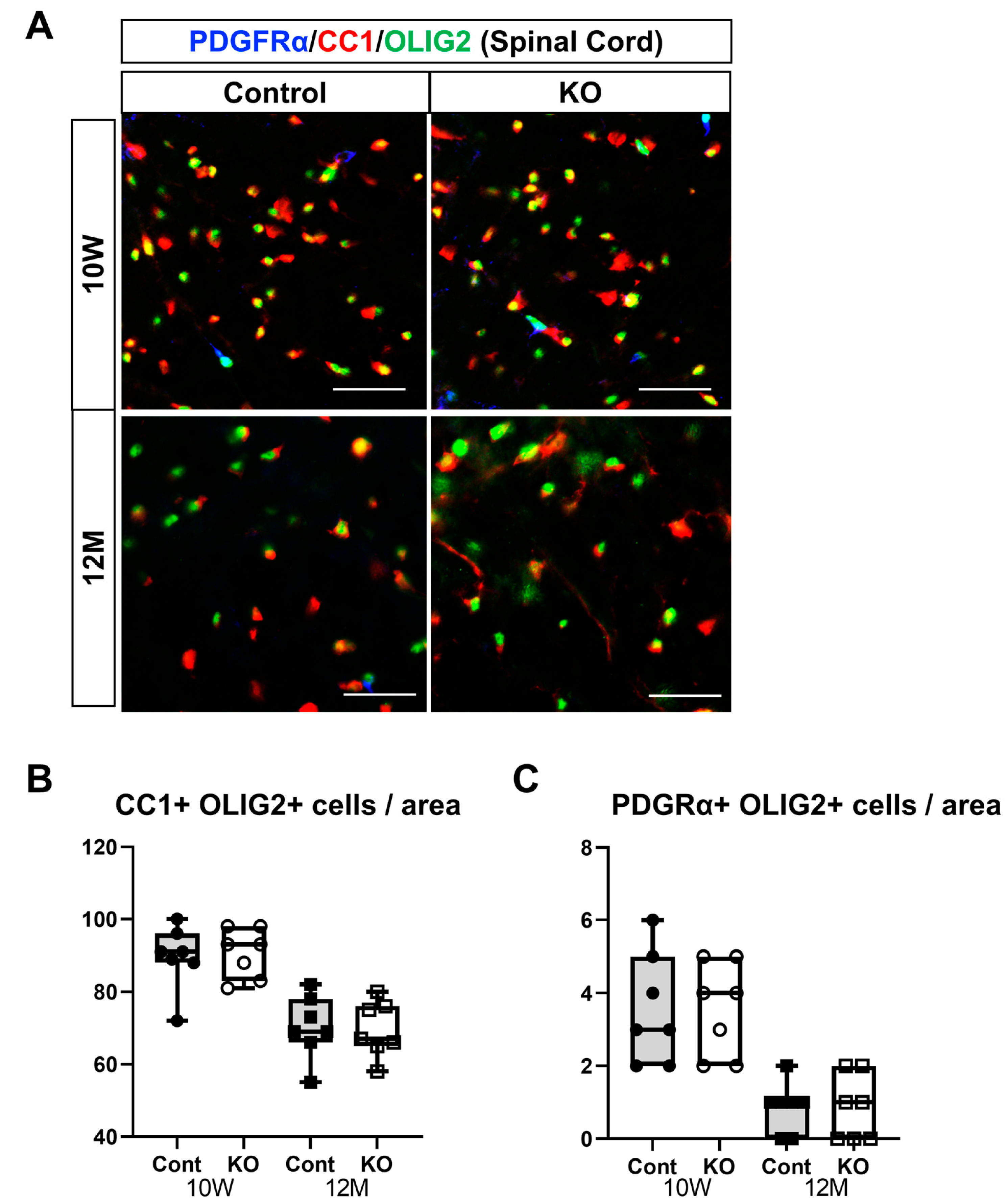
2.7. Immunohistochemistry
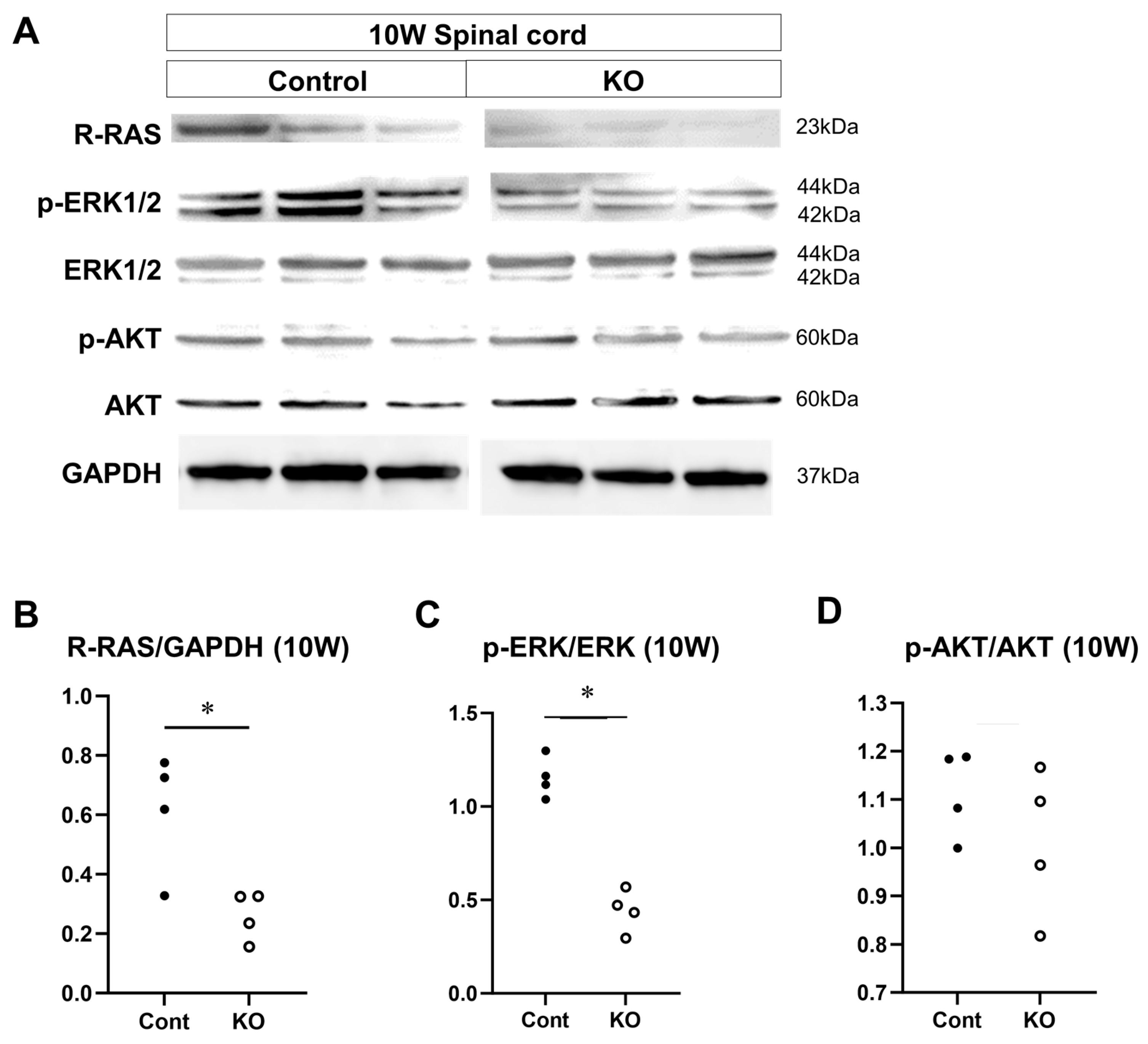
2.8. Western Blotting Analysis
2.9. Statistical Analysis
3. Results
3.1. Body Size and Morphology
3.2. Expression of the miR-23-27-24 Cluster
3.3. Impaired Motor Function in miR-23a/b Cluster-Deficient Mice
3.4. Hypoplasia of CNS Myelin Due to Deficiency of miR-23a/b Clusters
3.5. Effects of miR-23a/b Cluster Deletion on Oligodendrocyte Differentiation and Myelination
3.6. Decreased Expression of MBP and PLP Due to the Deletion of miR-23a/b Clusters
3.7. Candidate Target Molecules for miR-23a/b Clusters
3.8. Signaling Factors That are Altered by miR-23a/b Cluster Deletion
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Franklin, R.J.M.; Ffrench-Constant, C. Regenerating CNS myelin—From mechanisms to experimental medicines. Nat. Rev. Neurosci. 2017, 18, 753–769. [Google Scholar] [CrossRef]
- Micu, I.; Plemel, J.R.; Caprariello, A.V.; Nave, K.A.; Stys, P.K. Axo-myelinic neurotransmission: A novel mode of cell signalling in the central nervous system. Nat. Rev. Neurosci. 2018, 19, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Nave, K.A. Myelination and support of axonal integrity by glia. Nature 2010, 468, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Morrison, B.M.; Li, Y.; Lengacher, S.; Farah, M.H.; Hoffman, P.N.; Liu, Y.; Tsingalia, A.; Jin, L.; Zhang, P.W.; et al. Oligodendroglia metabolically support axons and contribute to neurodegeneration. Nature 2012, 487, 443–448. [Google Scholar] [CrossRef]
- Goldschmidt, T.; Antel, J.; Konig, F.B.; Bruck, W.; Kuhlmann, T. Remyelination capacity of the MS brain decreases with disease chronicity. Neurology 2009, 72, 1914–1921. [Google Scholar] [CrossRef]
- Huang, H.; Zhao, X.F.; Zheng, K.; Qiu, M. Regulation of the timing of oligodendrocyte differentiation: Mechanisms and perspectives. Neurosci. Bull. 2013, 29, 155–164. [Google Scholar] [CrossRef]
- Emery, B.; Lu, Q.R. Transcriptional and Epigenetic Regulation of Oligodendrocyte Development and Myelination in the Central Nervous System. Cold Spring Harb. Perspect. Biol. 2015, 7, a020461. [Google Scholar] [CrossRef] [PubMed]
- Maciak, K.; Dziedzic, A.; Saluk, J. Remyelination in multiple sclerosis from the miRNA perspective. Front. Mol. Neurosci. 2023, 16, 1199313. [Google Scholar] [CrossRef]
- Li, J.S.; Yao, Z.X. MicroRNAs: Novel regulators of oligodendrocyte differentiation and potential therapeutic targets in demyelination-related diseases. Mol. Neurobiol. 2012, 45, 200–212. [Google Scholar] [CrossRef]
- Kornfeld, S.F.; Cummings, S.E.; Fathi, S.; Bonin, S.R.; Kothary, R. MiRNA-145-5p prevents differentiation of oligodendrocyte progenitor cells by regulating expression of myelin gene regulatory factor. J. Cell. Physiol. 2021, 236, 997–1012. [Google Scholar] [CrossRef]
- Wang, H.; Moyano, A.L.; Ma, Z.; Deng, Y.; Lin, Y.; Zhao, C.; Zhang, L.; Jiang, M.; He, X.; Ma, Z.; et al. miR-219 Cooperates with miR-338 in Myelination and Promotes Myelin Repair in the CNS. Dev. Cell. 2017, 40, 566–582 e565. [Google Scholar] [CrossRef]
- Tripathi, A.; Volsko, C.; Garcia, J.P.; Agirre, E.; Allan, K.C.; Tesar, P.J.; Trapp, B.D.; Castelo-Branco, G.; Sim, F.J.; Dutta, R. Oligodendrocyte Intrinsic miR-27a Controls Myelination and Remyelination. Cell Rep. 2019, 29, 904–919 e909. [Google Scholar] [CrossRef]
- Lin, S.T.; Huang, Y.; Zhang, L.; Heng, M.Y.; Ptacek, L.J.; Fu, Y.H. MicroRNA-23a promotes myelination in the central nervous system. Proc. Natl. Acad. Sci. USA 2013, 110, 17468–17473. [Google Scholar] [CrossRef] [PubMed]
- Tsuchikawa, Y.; Kamei, N.; Sanada, Y.; Nakamae, T.; Harada, T.; Imaizumi, K.; Akimoto, T.; Miyaki, S.; Adachi, N. Deficiency of MicroRNA-23-27-24 Clusters Exhibits the Impairment of Myelination in the Central Nervous System. Neural Plast. 2023, 2023, 8938674. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Cambi, F. MicroRNA expression in mouse oligodendrocytes and regulation of proteolipid protein gene expression. J. Neurosci. Res. 2012, 90, 1701–1712. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Luo, J.; Zhang, H.; Lu, J. microRNAs in the Same Clusters Evolve to Coordinately Regulate Functionally Related Genes. Mol. Biol. Evol. 2016, 33, 2232–2247. [Google Scholar] [CrossRef]
- Lee, M.; Wada, S.; Oikawa, S.; Suzuki, K.; Ushida, T.; Akimoto, T. Loss of microRNA-23-27-24 clusters in skeletal muscle is not influential in skeletal muscle development and exercise-induced muscle adaptation. Sci. Rep. 2019, 9, 1092. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Ding, C.; Sanada, Y.; Yimiti, D.; Ishikawa, M.; Nakasa, T.; Kamei, N.; Imaizumi, K.; Lotz, M.K.; Akimoto, T.; et al. miR-23a/b clusters are not essential for the pathogenesis of osteoarthritis in mouse aging and post-traumatic models. Front. Cell Dev. Biol. 2022, 10, 1043259. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, H.; Hasuwa, H.; Inoue, N.; Ikawa, M.; Okabe, M. Lineage-specific cell disruption in living mice by Cre-mediated expression of diphtheria toxin A chain. Biochem. Biophys. Res. Commun. 2004, 321, 275–279. [Google Scholar] [CrossRef]
- Oikawa, S.; Wada, S.; Lee, M.; Maeda, S.; Akimoto, T. Role of endothelial microRNA-23 clusters in angiogenesis in vivo. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H838–H846. [Google Scholar] [CrossRef]
- Doerflinger, N.H.; Macklin, W.B.; Popko, B. Inducible site-specific recombination in myelinating cells. Genesis 2003, 35, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ma, X.; Hu, X.; Wen, J.; Zhang, H.; Xu, J.; He, Y.; Wang, X.; Guo, J. Schwann cell-specific RhoA knockout accelerates peripheral nerve regeneration via promoting Schwann cell dedifferentiation. Glia 2023, 71, 1715–1728. [Google Scholar] [CrossRef] [PubMed]
- Aartsma-Rus, A.; van Putten, M. Assessing functional performance in the mdx mouse model. J. Vis. Exp. 2014, 27, 51303. [Google Scholar]
- Orenduff, M.C.; Rezeli, E.T.; Hursting, S.D.; Pieper, C.F. Psychometrics of the Balance Beam Functional Test in C57BL/6 Mice. Comp. Med. 2021, 71, 302–308. [Google Scholar] [CrossRef]
- Lin, S.T.; Fu, Y.H. miR-23 regulation of lamin B1 is crucial for oligodendrocyte development and myelination. Dis. Model. Mech. 2009, 2, 178–188. [Google Scholar] [CrossRef]
- Baumann, N.; Pham-Dinh, D. Biology of oligodendrocyte and myelin in the mammalian central nervous system. Physiol. Rev. 2001, 81, 871–927. [Google Scholar] [CrossRef]
- Flores, A.I.; Narayanan, S.P.; Morse, E.N.; Shick, H.E.; Yin, X.; Kidd, G.; Avila, R.L.; Kirschner, D.A.; Macklin, W.B. Constitutively active Akt induces enhanced myelination in the CNS. J. Neurosci. 2008, 28, 7174–7183. [Google Scholar] [CrossRef]
- Rivers, L.E.; Young, K.M.; Rizzi, M.; Jamen, F.; Psachoulia, K.; Wade, A.; Kessaris, N.; Richardson, W.D. PDGFRA/NG2 glia generate myelinating oligodendrocytes and piriform projection neurons in adult mice. Nat. Neurosci. 2008, 11, 1392–1401. [Google Scholar] [CrossRef] [PubMed]
- Emery, B. Regulation of oligodendrocyte differentiation and myelination. Science 2010, 330, 779–782. [Google Scholar] [CrossRef] [PubMed]
- Ngo, C.; Kothary, R. MicroRNAs in oligodendrocyte development and remyelination. J. Neurochem. 2022, 162, 310–321. [Google Scholar] [CrossRef]
- Zhou, B.; Zhu, Z.; Ransom, B.R.; Tong, X. Oligodendrocyte lineage cells and depression. Mol. Psychiatry 2021, 26, 103–117. [Google Scholar] [CrossRef]
- Harlow, D.E.; Saul, K.E.; Culp, C.M.; Vesely, E.M.; Macklin, W.B. Expression of proteolipid protein gene in spinal cord stem cells and early oligodendrocyte progenitor cells is dispensable for normal cell migration and myelination. J. Neurosci. 2014, 34, 1333–1343. [Google Scholar] [CrossRef]
- Leone, D.P.; Genoud, S.; Atanasoski, S.; Grausenburger, R.; Berger, P.; Metzger, D.; Macklin, W.B.; Chambon, P.; Suter, U. Tamoxifen-inducible glia-specific Cre mice for somatic mutagenesis in oligodendrocytes and Schwann cells. Mol. Cell. Neurosci. 2003, 22, 430–440. [Google Scholar] [CrossRef]
- Lin, S.T.; Heng, M.Y.; Ptacek, L.J.; Fu, Y.H. Regulation of Myelination in the Central Nervous System by Nuclear Lamin B1 and Non-coding RNAs. Transl. Neurodegener. 2014, 3, 4. [Google Scholar] [CrossRef]
- Alcover-Sanchez, B.; Garcia-Martin, G.; Escudero-Ramirez, J.; Gonzalez-Riano, C.; Lorenzo, P.; Gimenez-Cassina, A.; Formentini, L.; de la Villa-Polo, P.; Pereira, M.P.; Wandosell, F.; et al. Absence of R-Ras1 and R-Ras2 causes mitochondrial alterations that trigger axonal degeneration in a hypomyelinating disease model. Glia 2021, 69, 619–637. [Google Scholar] [CrossRef]
- Sanz-Rodriguez, M.; Gruart, A.; Escudero-Ramirez, J.; de Castro, F.; Delgado-Garcia, J.M.; Wandosell, F.; Cubelos, B. R-Ras1 and R-Ras2 Are Essential for Oligodendrocyte Differentiation and Survival for Correct Myelination in the Central Nervous System. J. Neurosci. 2018, 38, 5096–5110. [Google Scholar] [CrossRef] [PubMed]
- Ishii, A.; Furusho, M.; Macklin, W.; Bansal, R. Independent and cooperative roles of the Mek/ERK1/2-MAPK and PI3K/Akt/mTOR pathways during developmental myelination and in adulthood. Glia 2019, 67, 1277–1295. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; He, W.; Tang, T.; Qiu, M. Immunological Markers for Central Nervous System Glia. Neurosci. Bull. 2023, 39, 379–392. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brain Corpus Callosum | Brain Gray Matter | |||||
|---|---|---|---|---|---|---|
| Control | KO | p Value | Control | KO | p Value | |
| miR-23a | 1.00 ± 0.35 | 0.43 ± 0.20 | <0.01 | 1.00 ± 0.31 | 1.24 ± 0.42 | 0.34 |
| miR-23b | 1.00 ± 0.29 | 0.39 ± 0.20 | <0.01 | 1.00 ± 0.39 | 1.23 ± 0.44 | 0.69 |
| miR-24 | 1.00 ± 0.37 | 0.32 ± 0.10 | <0.01 | 1.00 ± 0.32 | 1.13 ± 0.16 | 0.69 |
| miR-27a | 1.00 ± 0.24 | 0.26 ± 0.14 | <0.01 | 1.00 ± 0.35 | 1.04 ± 0.34 | 0.89 |
| miR-27b | 1.00 ± 0.45 | 0.25 ± 0.18 | <0.01 | 1.00 ± 0.30 | 1.14 ± 0.37 | 0.69 |
| Spinal cord white matter | Spinal cord gray matter | |||||
| Control | KO | p value | Control | KO | p value | |
| miR-23a | 1.00 ± 0.59 | 0.40 ± 0.23 | 0.03 | 1.00 ± 0.24 | 0.91 ± 0.44 | 0.49 |
| miR-23b | 1.00 ± 0.46 | 0.36 ± 0.19 | 0.01 | 1.00 ± 0.43 | 0.71 ± 0.31 | 0.49 |
| miR-24 | 1.00 ± 0.50 | 0.30 ± 0.13 | <0.01 | 1.00 ± 0.32 | 0.90 ± 0.17 | 0.69 |
| miR-27a | 1.00 ± 0.40 | 0.29 ± 0.25 | <0.01 | 1.00 ± 0.07 | 1.09 ± 0.48 | 1.00 |
| miR-27b | 1.00 ± 0.35 | 0.15 ± 0.06 | <0.01 | 1.00 ± 0.36 | 0.75 ± 0.37 | 0.34 |
| Liver | Lung | |||||
| Control | KO | p value | Control | KO | p value | |
| miR-23a | 1.00 ± 0.45 | 0.90 ± 0.29 | 0.89 | 1.00 ± 0.89 | 0.99 ± 0.80 | 0.69 |
| miR-23b | 1.00 ± 0.33 | 0.83 ± 0.25 | 0.34 | 1.00 ± 0.75 | 1.13 ± 0.83 | 0.89 |
| miR-24 | 1.00 ± 0.47 | 0.94 ± 0.23 | 0.89 | 1.00 ± 0.85 | 1.07 ± 0.80 | 0.49 |
| miR-27a | 1.00 ± 0.53 | 0.61 ± 0.23 | 0.34 | 1.00 ± 0.97 | 0.98 ± 0.90 | 0.89 |
| miR-27b | 1.00 ± 0.51 | 0.65 ± 0.14 | 0.34 | 1.00 ± 1.11 | 0.95 ± 0.83 | 0.69 |
| Muscle | ||||||
| Control | KO | p value | ||||
| miR-23a | 1.00 ± 0.33 | 1.09 ± 0.70 | 0.69 | |||
| miR-23b | 1.00 ± 0.29 | 1.02 ± 0.56 | 0.89 | |||
| miR-24 | 1.00 ± 0.29 | 1.01 ± 0.59 | 0.69 | |||
| miR-27a | 1.00 ± 0.56 | 0.94 ± 0.68 | 1.00 | |||
| miR-27b | 1.00 ± 0.41 | 1.00 ± 0.59 | 1.00 | |||
| Brain Corpus Callosum | Brain Gray Matter | |||||
|---|---|---|---|---|---|---|
| Control | KO | p Value | Control | KO | p Value | |
| miR-23a | 1.00 ± 0.68 | 0.12 ± 0.04 | <0.01 | 1.00 ± 0.34 | 0.90 ± 0.31 | 0.69 |
| miR-23b | 1.00 ± 0.66 | 0.11 ± 0.03 | <0.01 | 1.00 ± 0.14 | 0.91 ± 0.39 | 0.69 |
| miR-24 | 1.00 ± 0.67 | 0.13 ± 0.06 | <0.01 | 1.00 ± 0.09 | 1.10 ± 0.48 | 1.00 |
| miR-27a | 1.00 ± 0.83 | 0.07 ± 0.03 | <0.01 | 1.00 ± 0.27 | 0.96 ± 0.35 | 0.89 |
| miR-27b | 1.00 ± 0.98 | 0.06 ± 0.02 | <0.01 | 1.00 ± 0.23 | 1.01 ± 0.40 | 1.00 |
| Spinal cord white matter | Spinal cord gray matter | |||||
| Control | KO | p value | Control | KO | p value | |
| miR-23a | 1.00 ± 0.49 | 0.32 ± 0.12 | 0.03 | 1.00 ± 0.34 | 0.95 ± 0.33 | 1.00 |
| miR-23b | 1.00 ± 0.35 | 0.23 ± 0.11 | <0.01 | 1.00 ± 0.49 | 0.77 ± 0.43 | 0.89 |
| miR-24 | 1.00 ± 0.40 | 0.39 ± 0.20 | 0.02 | 1.00 ± 0.39 | 1.11 ± 0.33 | 0.89 |
| miR-27a | 1.00 ± 0.39 | 0.38 ± 0.19 | <0.01 | 1.00 ± 0.38 | 1.08 ± 0.19 | 0.89 |
| miR-27b | 1.00 ± 0.76 | 0.35 ± 0.32 | 0.03 | 1.00 ± 0.56 | 0.62 ± 0.42 | 0.49 |
| Liver | Lung | |||||
| Control | KO | p value | Control | KO | p value | |
| miR-23a | 1.00 ± 0.13 | 1.06 ± 0.23 | 0.69 | 1.00 ± 0.17 | 1.03 ± 0.22 | 0.89 |
| miR-23b | 1.00 ± 0.41 | 0.93 ± 0.20 | 0.89 | 1.00 ± 0.61 | 1.64 ± 0.83 | 0.69 |
| miR-24 | 1.00 ± 0.41 | 1.13 ± 0.36 | 0.34 | 1.00 ± 0.38 | 1.04 ± 0.29 | 0.89 |
| miR-27a | 1.00 ± 0.44 | 0.91 ± 0.30 | 1.00 | 1.00 ± 0.42 | 1.12 ± 0.49 | 0.89 |
| miR-27b | 1.00 ± 0.23 | 0.85 ± 0.22 | 0.69 | 1.00 ± 0.43 | 0.95 ± 0.24 | 1.00 |
| Muscle | ||||||
| Control | KO | p value | ||||
| miR-23a | 1.00 ± 0.32 | 1.27 ± 0.34 | 0.34 | |||
| miR-23b | 1.00 ± 0.42 | 1.23 ± 0.30 | 0.49 | |||
| miR-24 | 1.00 ± 0.40 | 1.34 ± 0.27 | 0.34 | |||
| miR-27a | 1.00 ± 0.41 | 1.04 ± 0.38 | 1.00 | |||
| miR-27b | 1.00 ± 0.43 | 0.95 ± 0.69 | 0.69 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishibashi, S.; Kamei, N.; Tsuchikawa, Y.; Nakamae, T.; Akimoto, T.; Miyaki, S.; Adachi, N. Myelin-Specific microRNA-23a/b Cluster Deletion Inhibits Myelination in the Central Nervous System during Postnatal Growth and Aging. Genes 2024, 15, 402. https://doi.org/10.3390/genes15040402
Ishibashi S, Kamei N, Tsuchikawa Y, Nakamae T, Akimoto T, Miyaki S, Adachi N. Myelin-Specific microRNA-23a/b Cluster Deletion Inhibits Myelination in the Central Nervous System during Postnatal Growth and Aging. Genes. 2024; 15(4):402. https://doi.org/10.3390/genes15040402
Chicago/Turabian StyleIshibashi, Shigeki, Naosuke Kamei, Yuji Tsuchikawa, Toshio Nakamae, Takayuki Akimoto, Shigeru Miyaki, and Nobuo Adachi. 2024. "Myelin-Specific microRNA-23a/b Cluster Deletion Inhibits Myelination in the Central Nervous System during Postnatal Growth and Aging" Genes 15, no. 4: 402. https://doi.org/10.3390/genes15040402