
Duplicated Copy Number Variant of the Maize 9-Lipoxygenase ZmLOX5 Improves 9,10-KODA-Mediated Resistance to Fall Armyworms

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
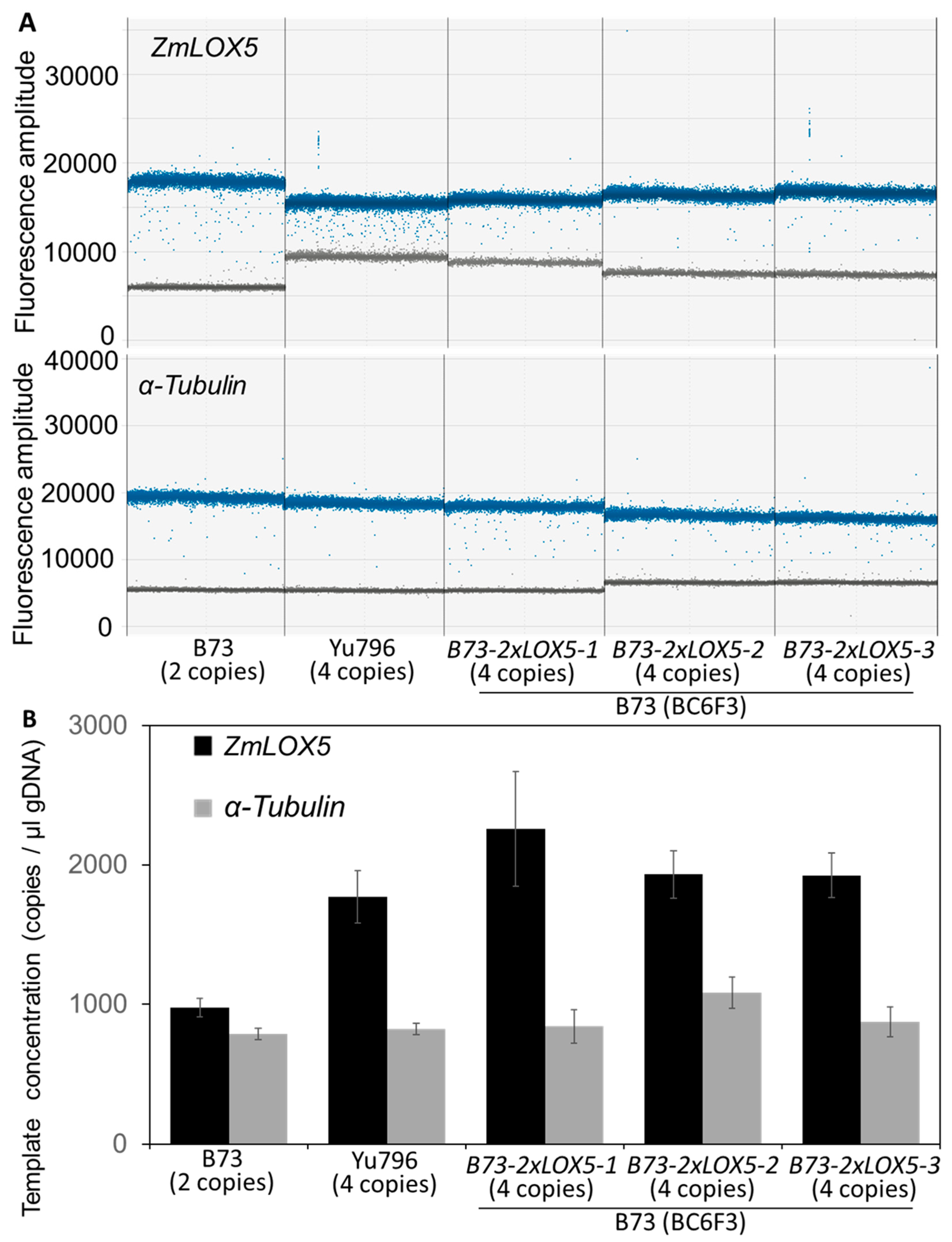
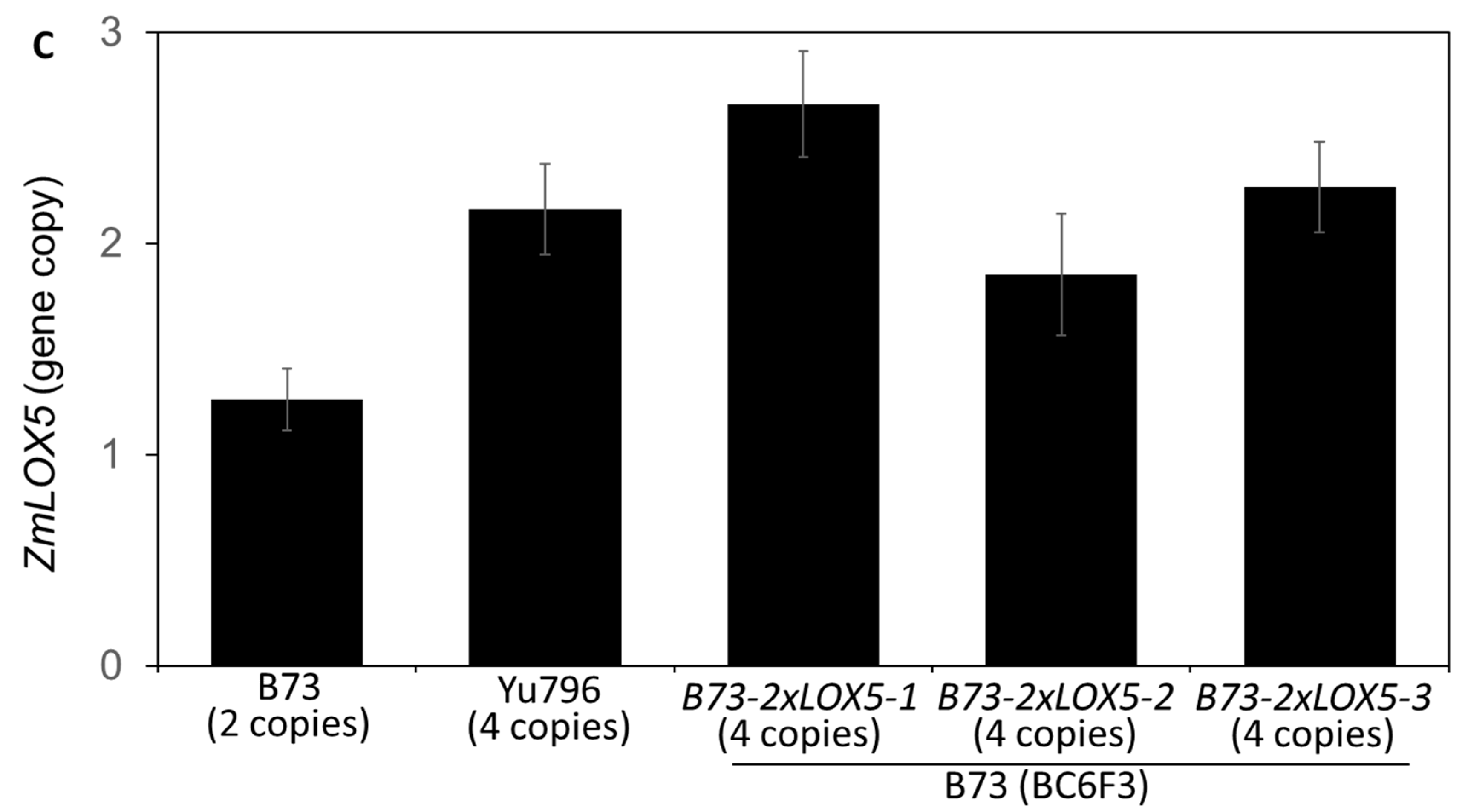
2.2. Genomic DNA Extraction and Droplet Digital PCR Analyses
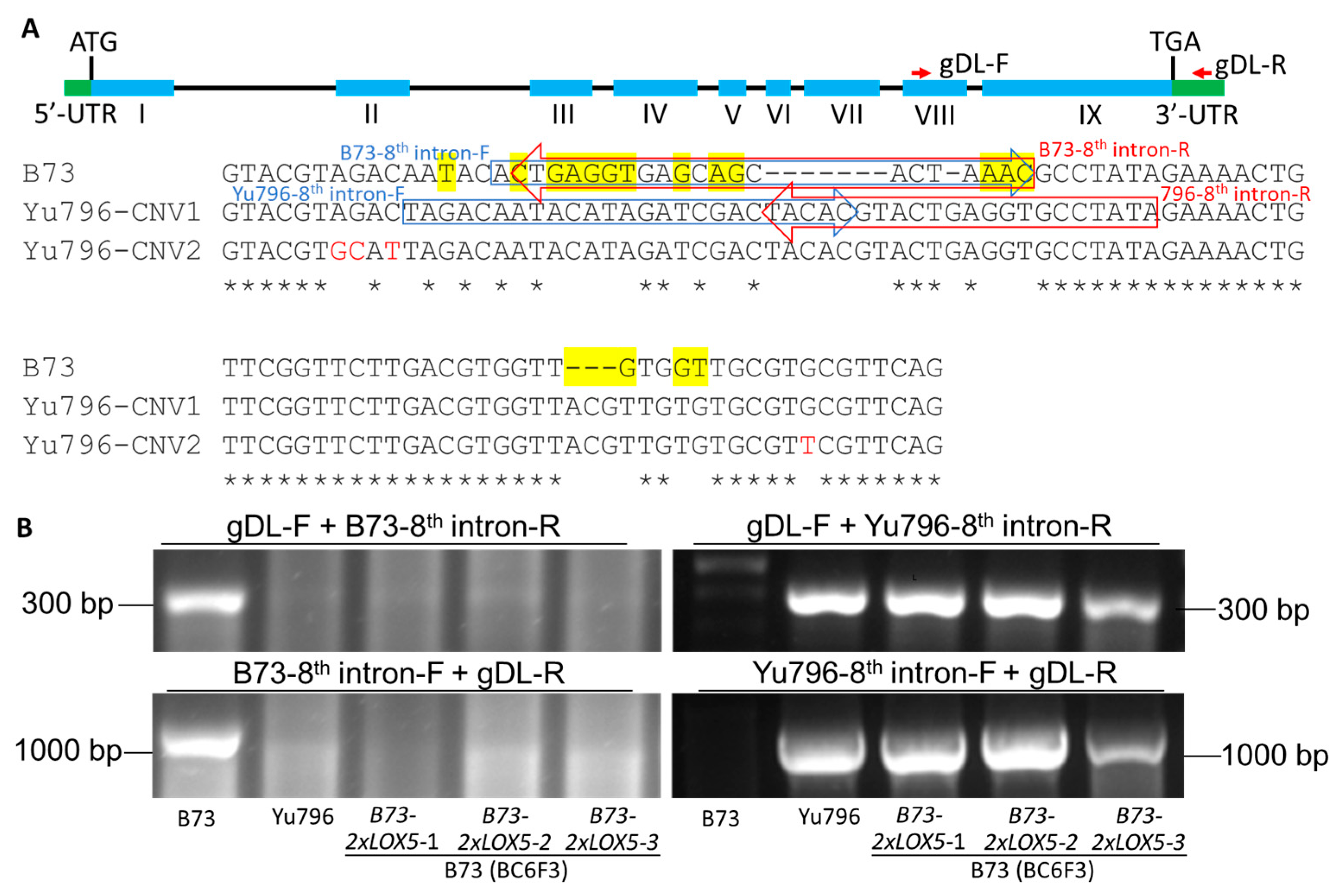
2.3. Sequence Analysis of 2×LOX5 CNVs
2.4. Mechanical Wounding of Leaves
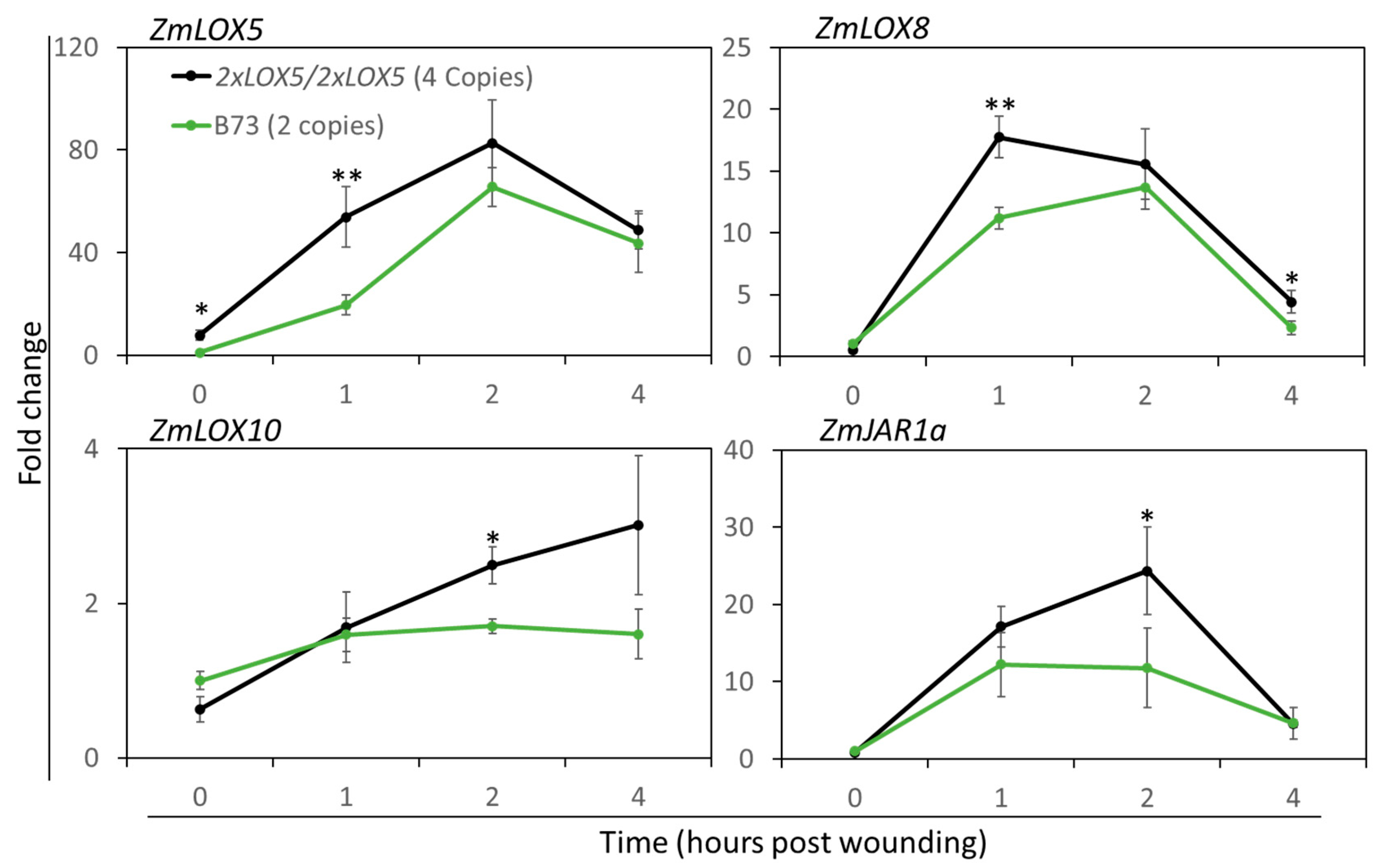
2.5. RNA Extraction and Expression Analyses Using qRT-PCR
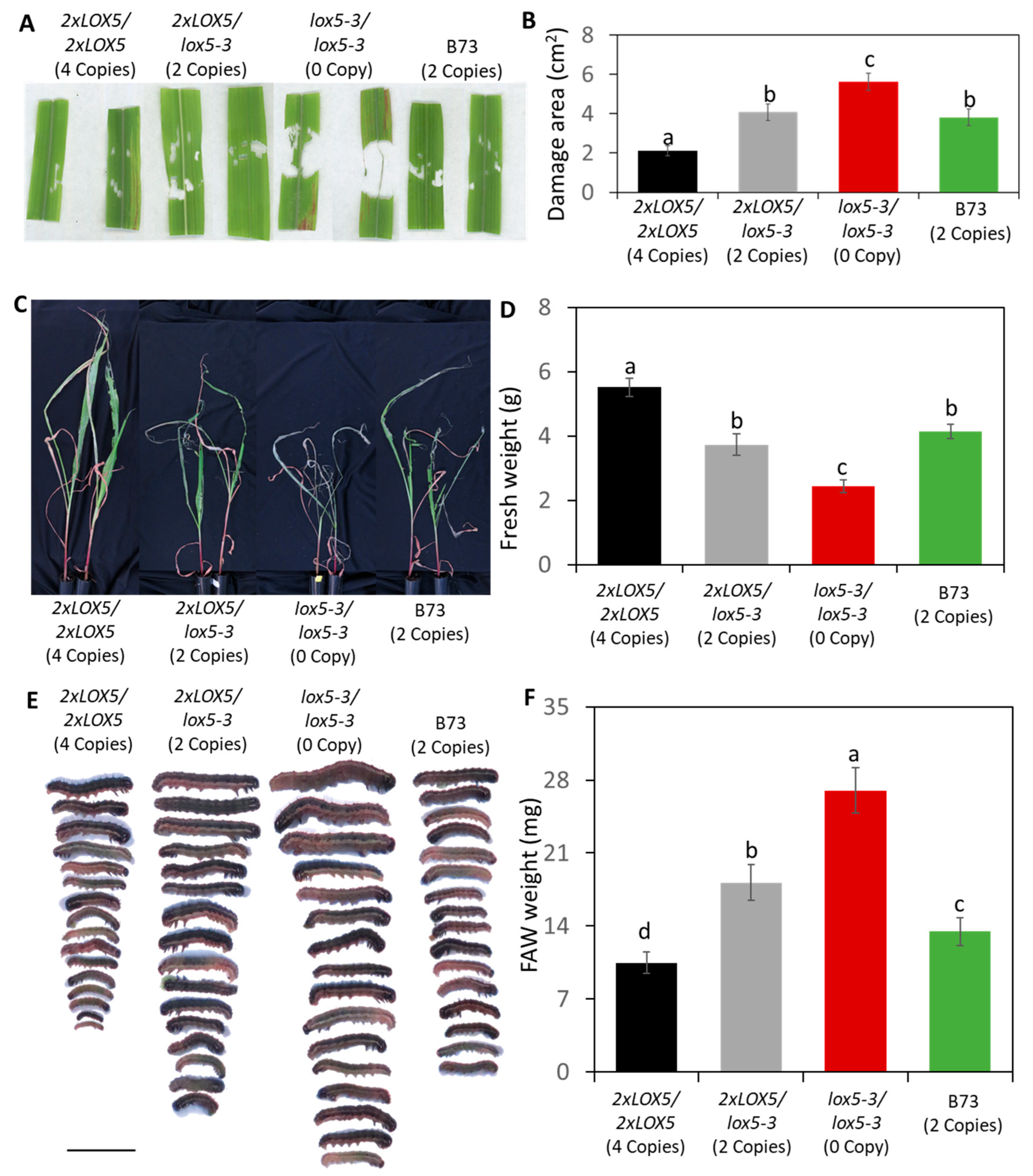
2.6. Fall Armyworm Resistance Assay
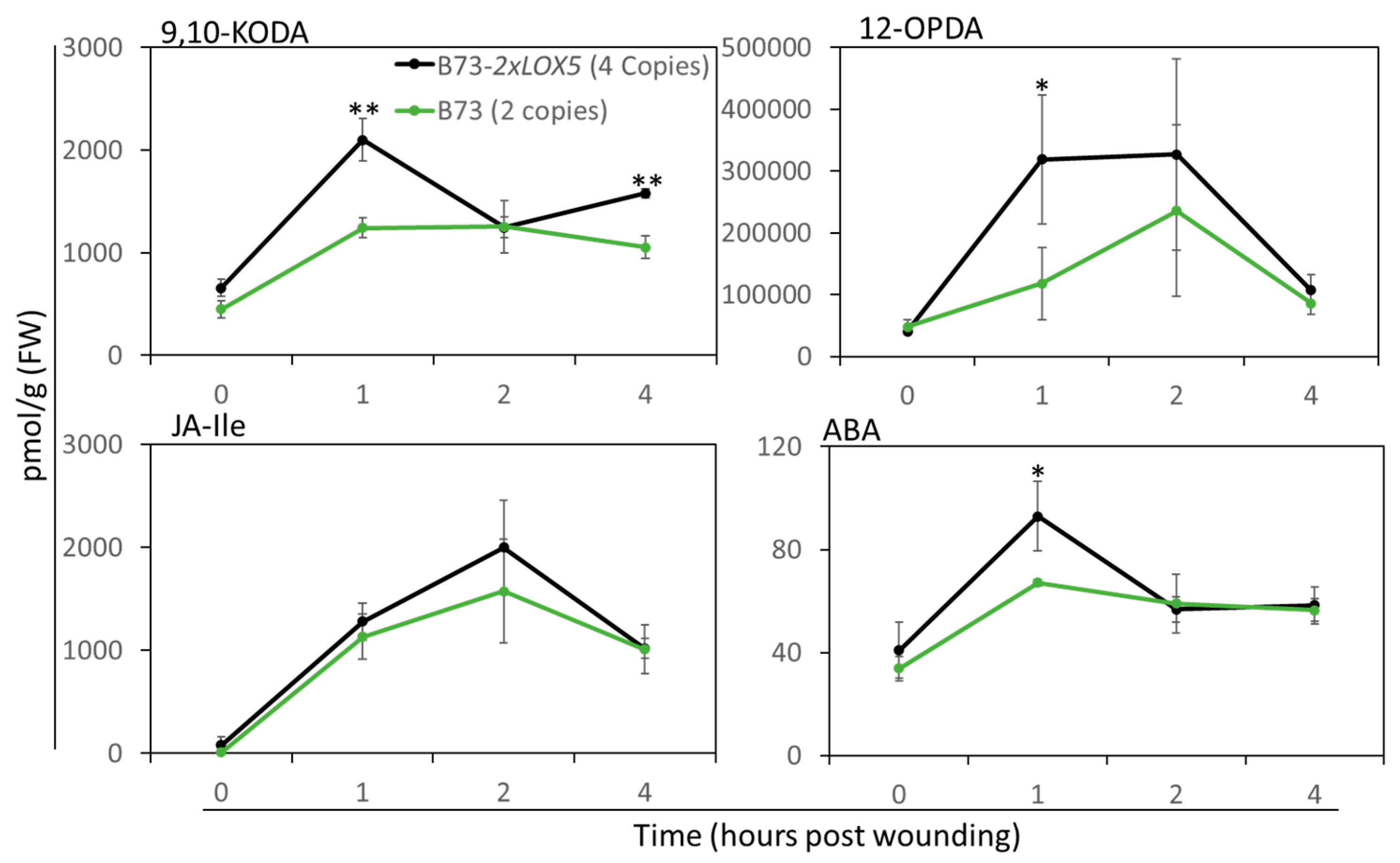
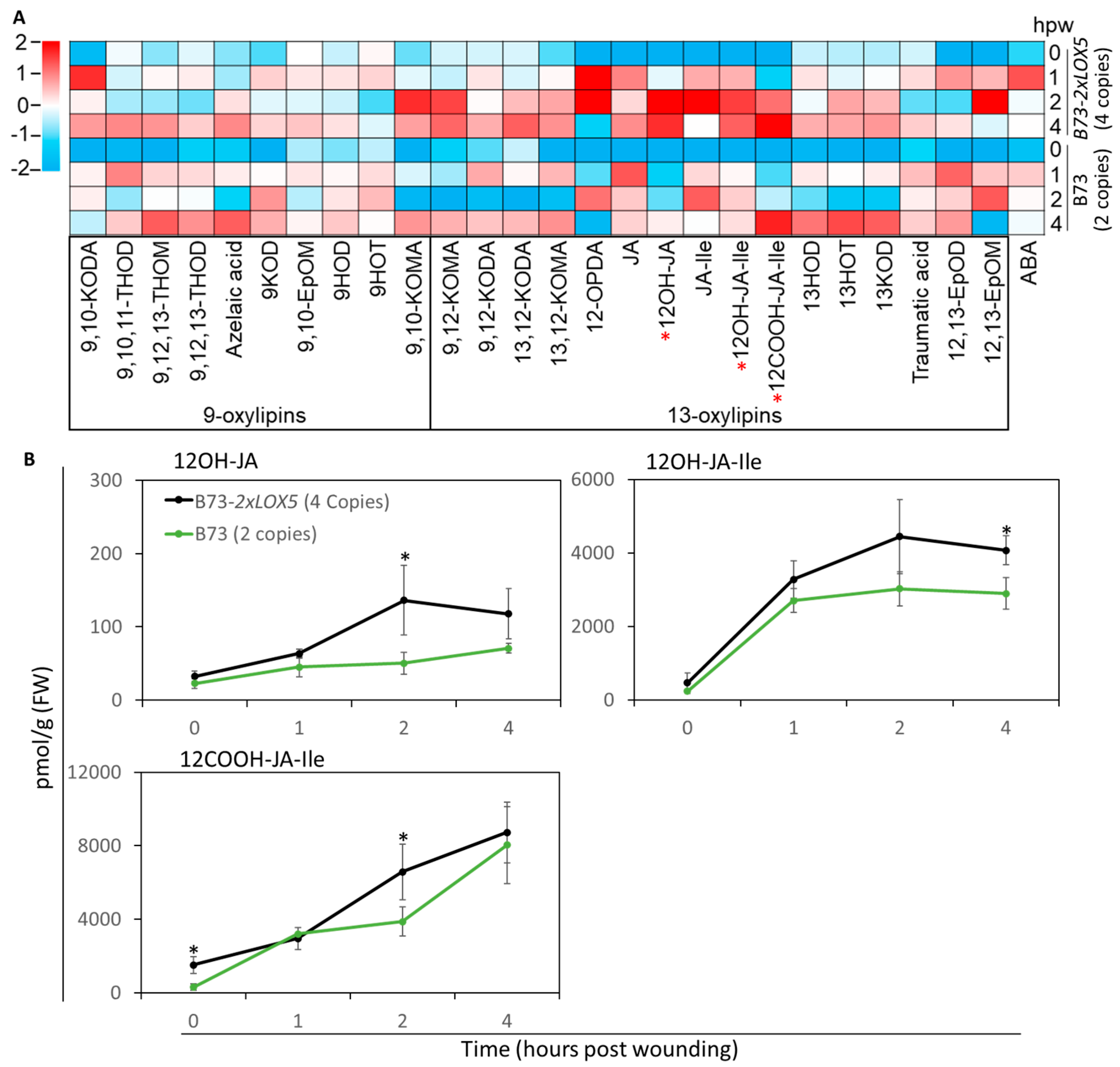
2.7. Quantification of Metabolites
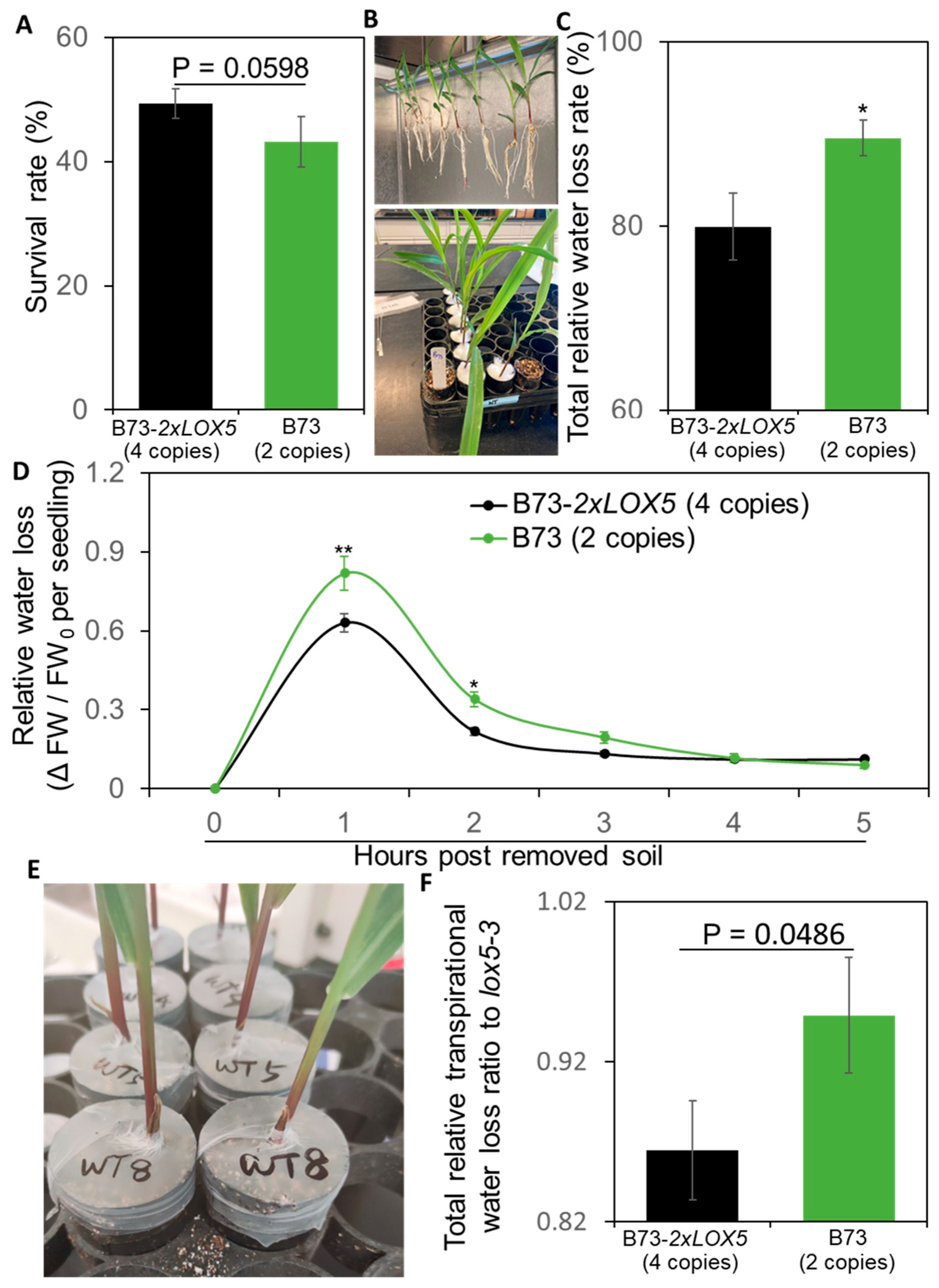
2.8. Drought Stress Test
2.9. Statistical Analysis
3. Results
3.1. Introgression of Duplicated Copy Variants of ZmLOX5 from Yu796 into B73
3.2. CNVs of Yu796-ZmLOX5 Are Tandemly Duplicated and Contain Multiple SNPs and Several InDel When Compared to the B73-ZmLOX5 Locus
3.3. Duplication of ZmLOX5 Leads to the Increased Expression of ZmLOX5 and Other Wound-Inducible Oxylipin Biosynthesis Genes
3.4. Duplication of ZmLOX5 Conferred Enhanced Resistance against FAW
3.5. Duplication of ZmLOX5-Promoted Wound-Induced Oxylipin and ABA Production
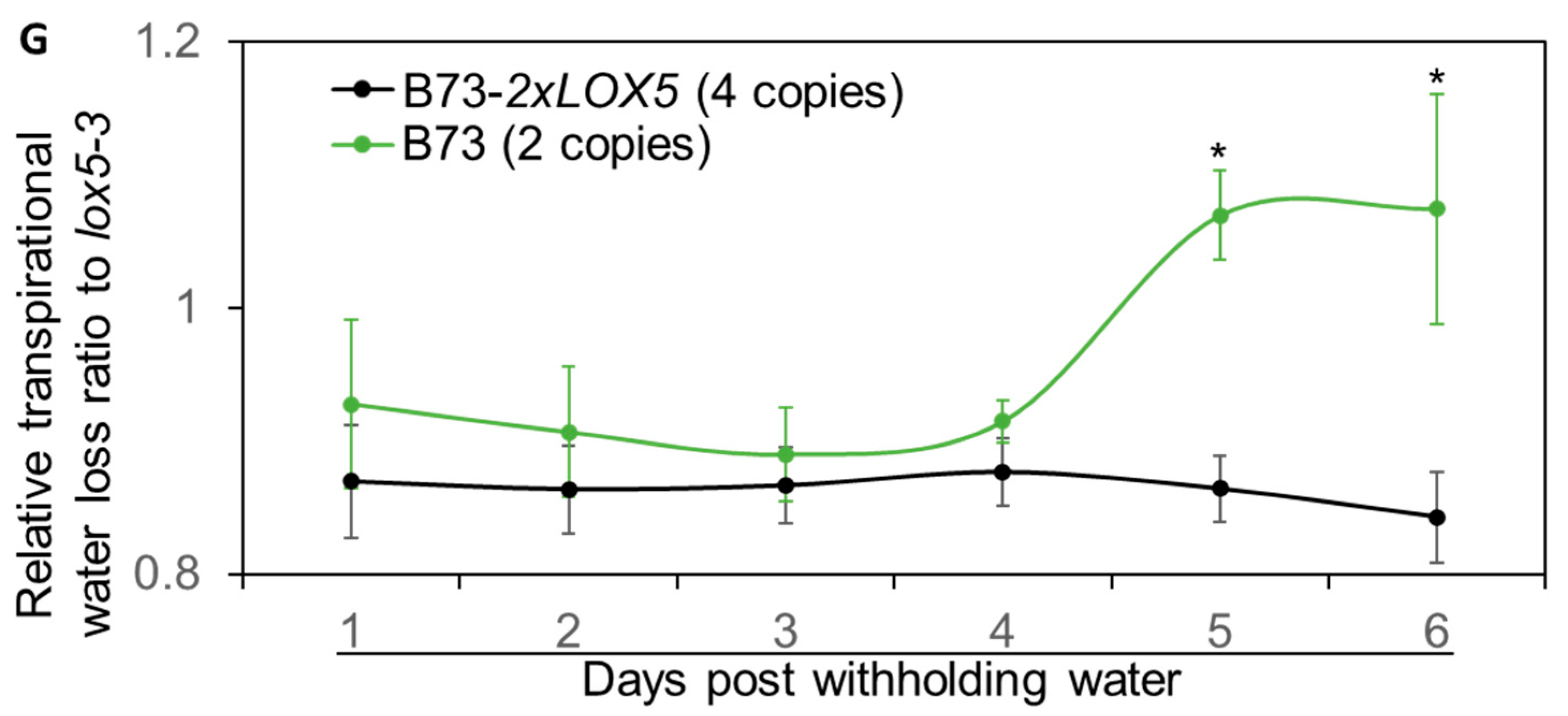
3.6. Duplication of ZmLOX5-Promoted Drought Tolerance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feldmann, F.; Rieckmann, U.; Winter, S. The spread of the fall armyworm Spodoptera frugiperda in Africa—What should be done next? J. Plant Dis. Prot. 2019, 126, 97–101. [Google Scholar] [CrossRef]
- Gassmann, A.J. Resistance to Bt Maize by Western Corn Rootworm: Effects of Pest Biology, the Pest-Crop Interaction and the Agricultural Landscape on Resistance. Insects 2021, 12, 136. [Google Scholar] [CrossRef] [PubMed]
- Csorba, A.B.; Fora, C.G.; Bálint, J.; Felföldi, T.; Szabó, A.; Máthé, I.; Loxdale, H.D.; Kentelky, E.; Nyárádi, I.-I.; Balog, A. Endosymbiotic Bacterial Diversity of Corn Leaf Aphid, Rhopalosiphum maidis Fitch (Hemiptera: Aphididae) Associated with Maize Management Systems. Microorganisms 2022, 10, 939. [Google Scholar] [CrossRef] [PubMed]
- Reay-Jones, F.P.F. Pest Status and Management of Corn Earworm (Lepidoptera: Noctuidae) in Field Corn in the United States. J. Integr. Pest Manag. 2019, 10, 19. [Google Scholar] [CrossRef]
- Tambo, J.A.; Day, R.K.; Lamontagne-Godwin, J.; Silvestri, S.; Beseh, P.K.; Oppong-Mensah, B.; Phiri, N.A.; Matimelo, M. Tackling fall armyworm (Spodoptera frugiperda) outbreak in Africa: An analysis of farmers’ control actions. Int. J. Pest Manag. 2020, 66, 298–310. [Google Scholar] [CrossRef]
- Overton, K.; Maino, J.L.; Day, R.; Umina, P.A.; Bett, B.; Carnovale, D.; Ekesi, S.; Meagher, R.; Reynolds, O.L. Global crop impacts, yield losses and action thresholds for fall armyworm (Spodoptera frugiperda): A review. Crop Prot. 2021, 145, 105641. [Google Scholar] [CrossRef]
- Clarkson, J.; Borah, J.R.; Baudron, F.; Sunderland, T.C.H. Forest Proximity Positively Affects Natural Enemy Mediated Control of Fall Armyworm in Southern Africa. Front. For. Glob. Chang. 2022, 5, 781574. [Google Scholar] [CrossRef]
- Kassie, M.; Wossen, T.; De Groote, H.; Tefera, T.; Sevgan, S.; Balew, S. Economic impacts of fall armyworm and its management strategies: Evidence from southern Ethiopia. Eur. Rev. Agric. Econ. 2020, 47, 1473–1501. [Google Scholar] [CrossRef]
- Makgoba, M.C.; Tshikhudo, P.P.; Nnzeru, L.R.; Makhado, R.A. Impact of fall armyworm (Spodoptera frugiperda) (J.E. Smith) on small-scale maize farmers and its control strategies in the Limpopo province, South Africa. Jàmbá J. Disaster Risk Stud. 2021, 13, 1016. [Google Scholar] [CrossRef]
- Kushwaha, U.K.S. A cost-efficient and alternative technique of managing fall armyworm Spodoptera frugiperda (J.E. Smith) larvae in maize crop. Sci. Rep. 2022, 12, 6741. [Google Scholar] [CrossRef]
- De Groote, H.; Kimenju, S.C.; Munyua, B.; Palmas, S.; Kassie, M.; Bruce, A. Spread and impact of fall armyworm (Spodoptera frugiperda J.E. Smith) in maize production areas of Kenya. Agric. Ecosyst. Environ. 2020, 292, 106804. [Google Scholar] [CrossRef] [PubMed]
- Paudel Timilsena, B.; Niassy, S.; Kimathi, E.; Abdel-Rahman, E.M.; Seidl-Adams, I.; Wamalwa, M.; Tonnang, H.E.Z.; Ekesi, S.; Hughes, D.P.; Rajotte, E.G.; et al. Potential distribution of fall armyworm in Africa and beyond, considering climate change and irrigation patterns. Sci. Rep. 2022, 12, 539. [Google Scholar] [CrossRef] [PubMed]
- Kansiime, M.K.; Rwomushana, I.; Mugambi, I. Fall armyworm invasion in Sub-Saharan Africa and impacts on community sustainability in the wake of Coronavirus Disease 2019: Reviewing the evidence. Curr. Opin. Environ. Sustain. 2023, 62, 101279. [Google Scholar] [CrossRef]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall Armyworm: Impacts and Implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Abbas, M.S.T. Genetically engineered (modified) crops (Bacillus thuringiensis crops) and the world controversy on their safety. Egypt. J. Biol. Pest Control 2018, 28, 52. [Google Scholar] [CrossRef]
- Raman, R. The impact of Genetically Modified (GM) crops in modern agriculture: A review. GM Crops Food 2017, 8, 195–208. [Google Scholar] [CrossRef] [PubMed]
- McConn, M.; Creelman, R.A.; Bell, E.; Mullet, J.E.; Browse, J. Jasmonate is essential for insect defense in Arabidopsis. Proc. Natl. Acad. Sci. USA 1997, 94, 5473–5477. [Google Scholar] [CrossRef]
- Koo, A.J.; Howe, G.A. The wound hormone jasmonate. Phytochemistry 2009, 70, 1571–1580. [Google Scholar] [CrossRef]
- Turner, J.G.; Ellis, C.; Devoto, A. The Jasmonate Signal Pathway. Plant Cell 2002, 14, s153–s164. [Google Scholar] [CrossRef]
- Staswick, P.E.; Su, W.; Howell, S.H. Methyl jasmonate inhibition of root growth and induction of a leaf protein are decreased in an Arabidopsis thaliana mutant. Proc. Natl. Acad. Sci. USA 1992, 89, 6837–6840. [Google Scholar] [CrossRef]
- Yuan, P.; Du, L.; Poovaiah, B. Ca2+/Calmodulin-Dependent AtSR1/CAMTA3 Plays Critical Roles in Balancing Plant Growth and Immunity. Int. J. Mol. Sci. 2018, 19, 1764. [Google Scholar] [CrossRef] [PubMed]
- Yuan, P.; Borrego, E.; Park, Y.-S.; Gorman, Z.; Huang, P.-C.; Tolley, J.; Christensen, S.A.; Blanford, J.; Kilaru, A.; Meeley, R.; et al. 9,10-KODA, an α-ketol produced by the tonoplast-localized 9-lipoxygenase ZmLOX5, plays a signaling role in maize defense against insect herbivory. Mol. Plant 2023, 16, 1283–1303. [Google Scholar] [CrossRef]
- Berg-Falloure, K.M.; Kolomiets, M.V. Ketols Emerge as Potent Oxylipin Signals Regulating Diverse Physiological Processes in Plants. Plants 2023, 12, 2088. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Aleman, G.H.; Jander, G. Maize defense against insect herbivory: A novel role for 9-LOX-derived oxylipins. Mol. Plant 2023, 16, 1484–1486. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-S.; Kunze, S.; Ni, X.; Feussner, I.; Kolomiets, M.V. Comparative molecular and biochemical characterization of segmentally duplicated 9-lipoxygenase genes ZmLOX4 and ZmLOX5 of maize. Planta 2010, 231, 1425–1437. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Borrego, E.J.; Gorman, Z.; Huang, P.C.; Kolomiets, M.V. Relative contribution of LOX10, green leaf volatiles and JA to wound-induced local and systemic oxylipin and hormone signature in Zea mays (maize). Phytochemistry 2020, 174, 112334. [Google Scholar] [CrossRef] [PubMed]
- Springer, N.M.; Ying, K.; Fu, Y.; Ji, T.; Yeh, C.-T.; Jia, Y.; Wu, W.; Richmond, T.; Kitzman, J.; Rosenbaum, H.; et al. Maize Inbreds Exhibit High Levels of Copy Number Variation (CNV) and Presence/Absence Variation (PAV) in Genome Content. PLOS Genet. 2009, 5, e1000734. [Google Scholar] [CrossRef] [PubMed]
- Dolatabadian, A.; Yuan, Y.; Bayer, P.E.; Petereit, J.; Severn-Ellis, A.; Tirnaz, S.; Patel, D.; Edwards, D.; Batley, J. Copy Number Variation among Resistance Genes Analogues in Brassica napus. Genes 2022, 13, 2037. [Google Scholar] [CrossRef]
- Żmieńko, A.; Samelak, A.; Kozłowski, P.; Figlerowicz, M. Copy number polymorphism in plant genomes. Theor. Appl. Genet. 2014, 127, 1–18. [Google Scholar] [CrossRef]
- De La Fuente, G.N.; Murray, S.C.; Isakeit, T.; Park, Y.-S.; Yan, Y.; Warburton, M.L.; Kolomiets, M.V. Characterization of Genetic Diversity and Linkage Disequilibrium of ZmLOX4 and ZmLOX5 Loci in Maize. PLoS ONE 2013, 8, e53973. [Google Scholar] [CrossRef]
- Cai, Y.M.; Dudley, Q.M.; Patron, N.J. Measurement of Transgene Copy Number in Plants Using Droplet Digital PCR. Bio-Protocol 2021, 11, e4075. [Google Scholar] [CrossRef]
- Bell, A.D.; Usher, C.L.; McCarroll, S.A. Analyzing Copy Number Variation with Droplet Digital PCR. Methods Mol. Biol. 2018, 1768, 143–160. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Joyce, P.A. Application of droplet digital PCR to determine copy number of endogenous genes and transgenes in sugarcane. Plant Cell Rep. 2017, 36, 1775–1783. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.-D.; Borrego, E.J.; Kenerley, C.M.; Kolomiets, M.V. Oxylipins Other Than Jasmonic Acid Are Xylem-Resident Signals Regulating Systemic Resistance Induced by Trichoderma virens in Maize. Plant Cell 2020, 32, 166–185. [Google Scholar] [CrossRef] [PubMed]
- Iwobi, A.; Gerdes, L.; Busch, U.; Pecoraro, S. Droplet digital PCR for routine analysis of genetically modified foods (GMO)—A comparison with real-time quantitative PCR. Food Control 2016, 69, 205–213. [Google Scholar] [CrossRef]
- Pires, A.M.; Branco, J.A. A Statistical Model to Explain the Mendel–Fisher Controversy. Stat. Sci. 2010, 25, 545–565. [Google Scholar] [CrossRef]
- Lye, Z.N.; Purugganan, M.D. Copy Number Variation in Domestication. Trends Plant Sci. 2019, 24, 352–365. [Google Scholar] [CrossRef]
- Lunde, C.; Kimberlin, A.; Leiboff, S.; Koo, A.J.; Hake, S. Tasselseed5 overexpresses a wound-inducible enzyme, ZmCYP94B1, that affects jasmonate catabolism, sex determination, and plant architecture in maize. Commun. Biol. 2019, 2, 114. [Google Scholar] [CrossRef]
- Huang, P.-C.; Grunseich, J.M.; Berg-Falloure, K.M.; Tolley, J.P.; Koiwa, H.; Bernal, J.S.; Kolomiets, M.V. Maize OPR2 and LOX10 Mediate Defense against Fall Armyworm and Western Corn Rootworm by Tissue-Specific Regulation of Jasmonic Acid and Ketol Metabolism. Genes 2023, 14, 1732. [Google Scholar] [CrossRef]
- Fujita, Y.; Fujita, M.; Satoh, R.; Maruyama, K.; Parvez, M.M.; Seki, M.; Hiratsu, K.; Ohme-Takagi, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1 Is a Transcription Activator of Novel ABRE-Dependent ABA Signaling That Enhances Drought Stress Tolerance in Arabidopsis. Plant Cell 2005, 17, 3470–3488. [Google Scholar] [CrossRef]
- Li, X.; Li, J.; Wei, S.; Gao, Y.; Pei, H.; Geng, R.; Lu, Z.; Wang, P.; Zhou, W. Maize GOLDEN2-LIKE proteins enhance drought tolerance in rice by promoting stomatal closure. Plant Physiol. 2023, 194, 774–786. [Google Scholar] [CrossRef] [PubMed]
- Leckie, C.P.; McAinsh, M.R.; Allen, G.J.; Sanders, D.; Hetherington, A.M. Abscisic acid-induced stomatal closure mediated by cyclic ADP-ribose. Proc. Natl. Acad. Sci. USA 1998, 95, 15837–15842. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Shi, M.; Sun, Y.; Cheng, H.; Ou, X.; Zhao, Y.; Zhang, X.; Day, B.; Miao, C.; Jiang, K. Light-induced stomatal opening in Arabidopsis is negatively regulated by chloroplast-originated OPDA signaling. Curr. Biol. 2023, 33, 1071–1081.e1075. [Google Scholar] [CrossRef] [PubMed]
- Kidd, J.M.; Cooper, G.M.; Donahue, W.F.; Hayden, H.S.; Sampas, N.; Graves, T.; Hansen, N.; Teague, B.; Alkan, C.; Antonacci, F.; et al. Mapping and sequencing of structural variation from eight human genomes. Nature 2008, 453, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.E.; Lee, T.G.; Guo, X.; Melito, S.; Wang, K.; Bayless, A.M.; Wang, J.; Hughes, T.J.; Willis, D.K.; Clemente, T.E.; et al. Copy Number Variation of Multiple Genes at Rhg1 Mediates Nematode Resistance in Soybean. Science 2012, 338, 1206–1209. [Google Scholar] [CrossRef]
- Yuan, P.; Poovaiah, B.W. Interplay between Ca2+/Calmodulin-Mediated Signaling and AtSR1/CAMTA3 during Increased Temperature Resulting in Compromised Immune Response in Plants. Int. J. Mol. Sci. 2022, 23, 2175. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Simeoni, F.; Skirycz, A.; Simoni, L.; Castorina, G.; de Souza, L.P.; Fernie, A.R.; Alseekh, S.; Giavalisco, P.; Conti, L.; Tonelli, C.; et al. The AtMYB60 transcription factor regulates stomatal opening by modulating oxylipin synthesis in guard cells. Sci. Rep. 2022, 12, 533. [Google Scholar] [CrossRef]
- Savchenko, T.; Kolla, V.A.; Wang, C.-Q.; Nasafi, Z.; Hicks, D.R.; Phadungchob, B.; Chehab, W.E.; Brandizzi, F.; Froehlich, J.; Dehesh, K. Functional Convergence of Oxylipin and Abscisic Acid Pathways Controls Stomatal Closure in Response to Drought. Plant Physiol. 2014, 164, 1151–1160. [Google Scholar] [CrossRef]
- Haque, E.; Osmani, A.A.; Ahmadi, S.H.; Ogawa, S.; Takagi, K.; Yokoyama, M.; Ban, T. KODA, an α-ketol derivative of linolenic acid provides wide recovery ability of wheat against various abiotic stresses. Biocatal. Agric. Biotechnol. 2016, 7, 67–75. [Google Scholar] [CrossRef]
- Kondrashov, F.A. Gene duplication as a mechanism of genomic adaptation to a changing environment. Proc. Biol. Sci. 2012, 279, 5048–5057. [Google Scholar] [CrossRef] [PubMed]
- Wingen, L.U.; Münster, T.; Faigl, W.; Deleu, W.; Sommer, H.; Saedler, H.; Theißen, G. Molecular genetic basis of pod corn (Tunicate maize). Proc. Natl. Acad. Sci. USA 2012, 109, 7115–7120. [Google Scholar] [CrossRef] [PubMed]
- Zuo, T.; Zhang, J.; Lithio, A.; Dash, S.; Weber, D.F.; Wise, R.; Nettleton, D.; Peterson, T. Genes and Small RNA Transcripts Exhibit Dosage-Dependent Expression Pattern in Maize Copy-Number Alterations. Genetics 2016, 203, 1133–1147. [Google Scholar] [CrossRef] [PubMed]
- Maron, L.G.; Guimarães, C.T.; Kirst, M.; Albert, P.S.; Birchler, J.A.; Bradbury, P.J.; Buckler, E.S.; Coluccio, A.E.; Danilova, T.V.; Kudrna, D.; et al. Aluminum tolerance in maize is associated with higher MATE1 gene copy number. Proc. Natl. Acad. Sci. USA 2013, 110, 5241–5246. [Google Scholar] [CrossRef]
- Gaudin, A.C.M.; McClymont, S.A.; Soliman, S.S.M.; Raizada, M.N. The effect of altered dosage of a mutant allele of Teosinte branched 1 (tb1-ref) on the root system of modern maize. BMC Genet. 2014, 15, 23. [Google Scholar] [CrossRef]
- Liu, H.; Shi, J.; Sun, C.; Gong, H.; Fan, X.; Qiu, F.; Huang, X.; Feng, Q.; Zheng, X.; Yuan, N.; et al. Gene duplication confers enhanced expression of 27-kDa γ-zein for endosperm modification in quality protein maize. Proc. Natl. Acad. Sci. USA 2016, 113, 4964–4969. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Ren, J.; Peng, Z.; Umana, A.A.; Le, H.; Danilova, T.; Fu, J.; Wang, H.; Robertson, A.; Hulbert, S.H.; et al. Analysis of Extreme Phenotype Bulk Copy Number Variation (XP-CNV) Identified the Association of rp1 with Resistance to Goss’s Wilt of Maize. Front. Plant Sci. 2018, 9, 110. [Google Scholar] [CrossRef] [PubMed]
- Hindson, B.J.; Ness, K.D.; Masquelier, D.A.; Belgrader, P.; Heredia, N.J.; Makarewicz, A.J.; Bright, I.J.; Lucero, M.Y.; Hiddessen, A.L.; Legler, T.C.; et al. High-Throughput Droplet Digital PCR System for Absolute Quantitation of DNA Copy Number. Anal. Chem. 2011, 83, 8604–8610. [Google Scholar] [CrossRef]
- Miotke, L.; Lau, B.T.; Rumma, R.T.; Ji, H.P. High Sensitivity Detection and Quantitation of DNA Copy Number and Single Nucleotide Variants with Single Color Droplet Digital PCR. Anal. Chem. 2014, 86, 2618–2624. [Google Scholar] [CrossRef]
- Qi, X.; Zhang, C.; Zhu, J.; Liu, C.; Huang, C.; Li, X.; Xie, C. Genome Editing Enables Next-Generation Hybrid Seed Production Technology. Mol. Plant 2020, 13, 1262–1269. [Google Scholar] [CrossRef]
- Wang, H.; Qi, X.; Zhu, J.; Liu, C.; Fan, H.; Zhang, X.; Li, X.; Yang, Q.; Xie, C. Pollen self-elimination CRISPR-Cas genome editing prevents transgenic pollen dispersal in maize. Plant Commun. 2023, 4, 100637. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2×LOX5/ 2×LOX5a | 2×LOX5/ lox5-3a | lox5-3/ lox5-3a | Actual Ratio | Chi-Square Value (χ2 ) for Tandem CNV | Fit Tandem CNV Ratio (χ2 < 5.991, DF = 2) b | Chi-Square Value (χ2) for Interspersed CNV | Fit Interspersed CNV Ratio (χ2 < 5.991, DF = 2) | |
|---|---|---|---|---|---|---|---|---|
| Tandem CNV | 1 | 2 | 1 | (1):(2):(1) | ||||
| Interspersed CNV | 1 | 14 | 1 | (1):(14):(1) | ||||
| BC4F2 c | 31 | 55 | 24 | (1.13):(2):(0.87) | 0.89 | Yes | 144.99 | No |
| BC5F2 c | 22 | 63 | 31 | (0.76):(2.17):(1.07) | 2.26 | Yes | 122.41 | No |
| BC6F2 c | 28 | 61 | 24 | (0.99):(2.16):(0.85) | 1.00 | Yes | 117.20 | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, P.; Huang, P.-C.; Martin, T.K.; Chappell, T.M.; Kolomiets, M.V. Duplicated Copy Number Variant of the Maize 9-Lipoxygenase ZmLOX5 Improves 9,10-KODA-Mediated Resistance to Fall Armyworms. Genes 2024, 15, 401. https://doi.org/10.3390/genes15040401
Yuan P, Huang P-C, Martin TK, Chappell TM, Kolomiets MV. Duplicated Copy Number Variant of the Maize 9-Lipoxygenase ZmLOX5 Improves 9,10-KODA-Mediated Resistance to Fall Armyworms. Genes. 2024; 15(4):401. https://doi.org/10.3390/genes15040401
Chicago/Turabian StyleYuan, Peiguo, Pei-Cheng Huang, Timothy K. Martin, Thomas M. Chappell, and Michael V. Kolomiets. 2024. "Duplicated Copy Number Variant of the Maize 9-Lipoxygenase ZmLOX5 Improves 9,10-KODA-Mediated Resistance to Fall Armyworms" Genes 15, no. 4: 401. https://doi.org/10.3390/genes15040401