
Spatiotemporal Expression Characterization of KRTAP6 Family Genes and Its Effect on Wool Traits
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Samples
2.2. In Situ Hybridization Analysis
2.3. RT-qPCR Analysis
2.4. Measurements and Statistical Analysis
3. Results
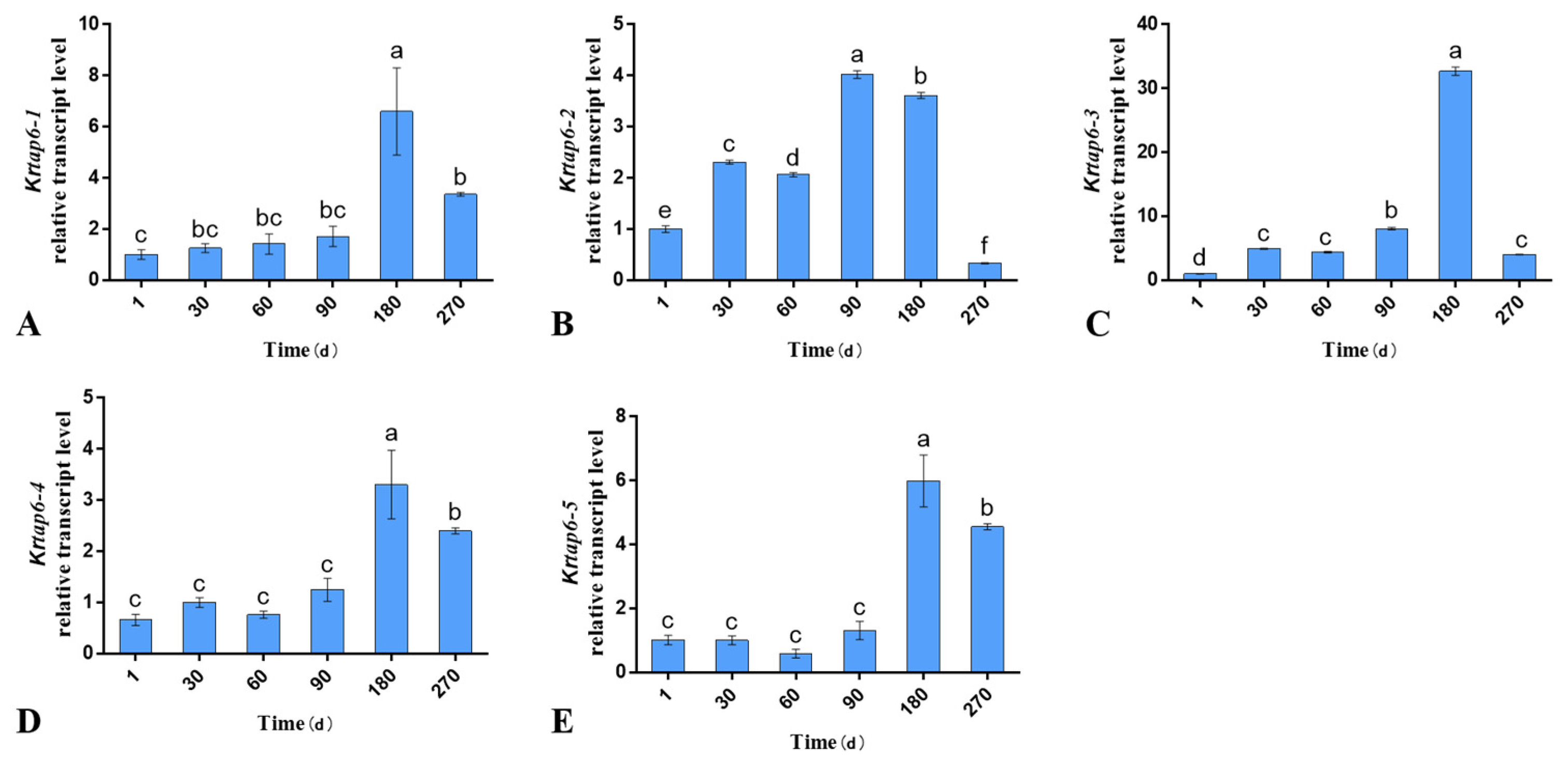
3.1. Expression of KRTAP6 Family Genes in Different Stages of Follicle Cycle Development of Skin
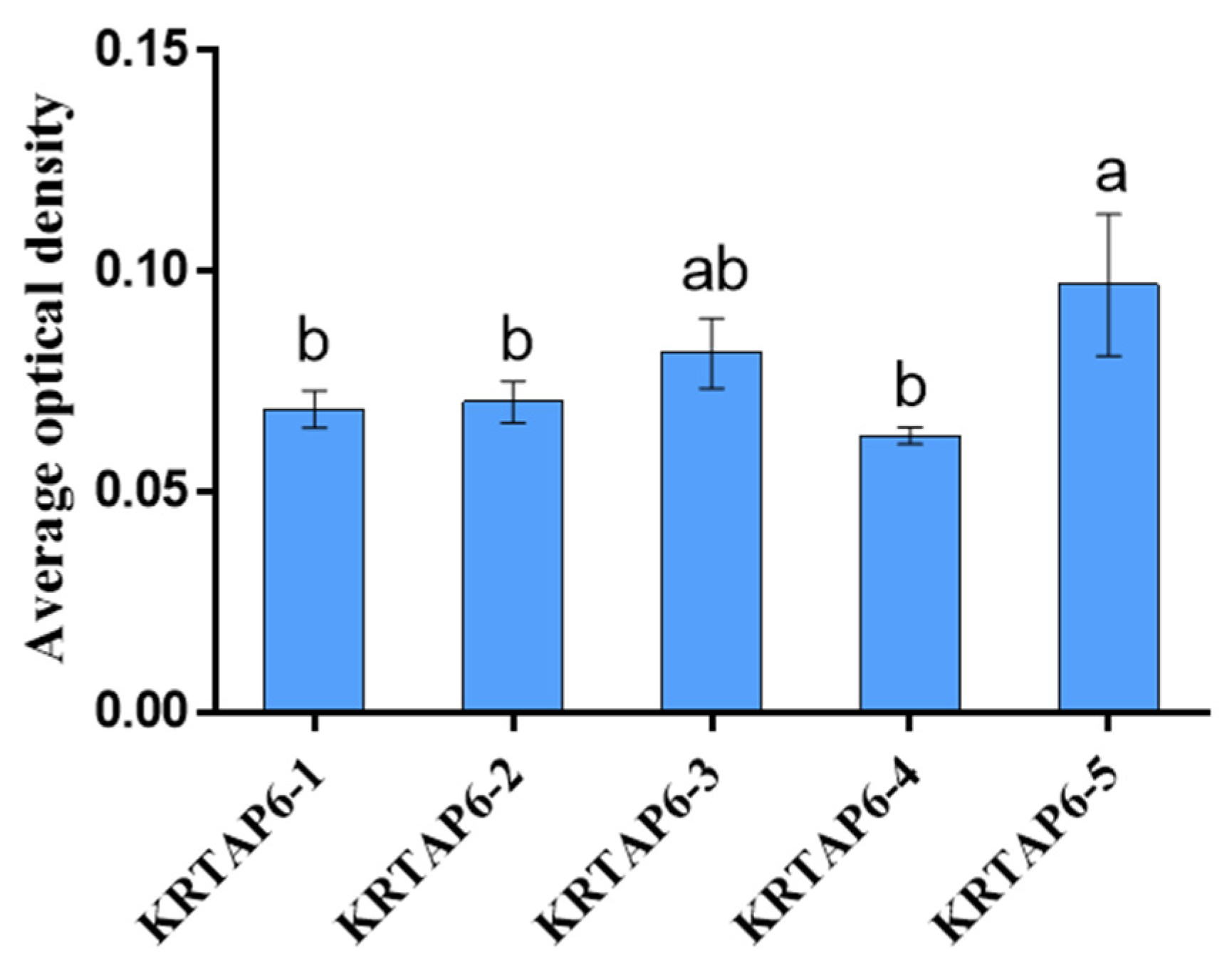
3.2. AOD Values of KRTAP6 Family at 180 Days of Hair Follicle in the Skin
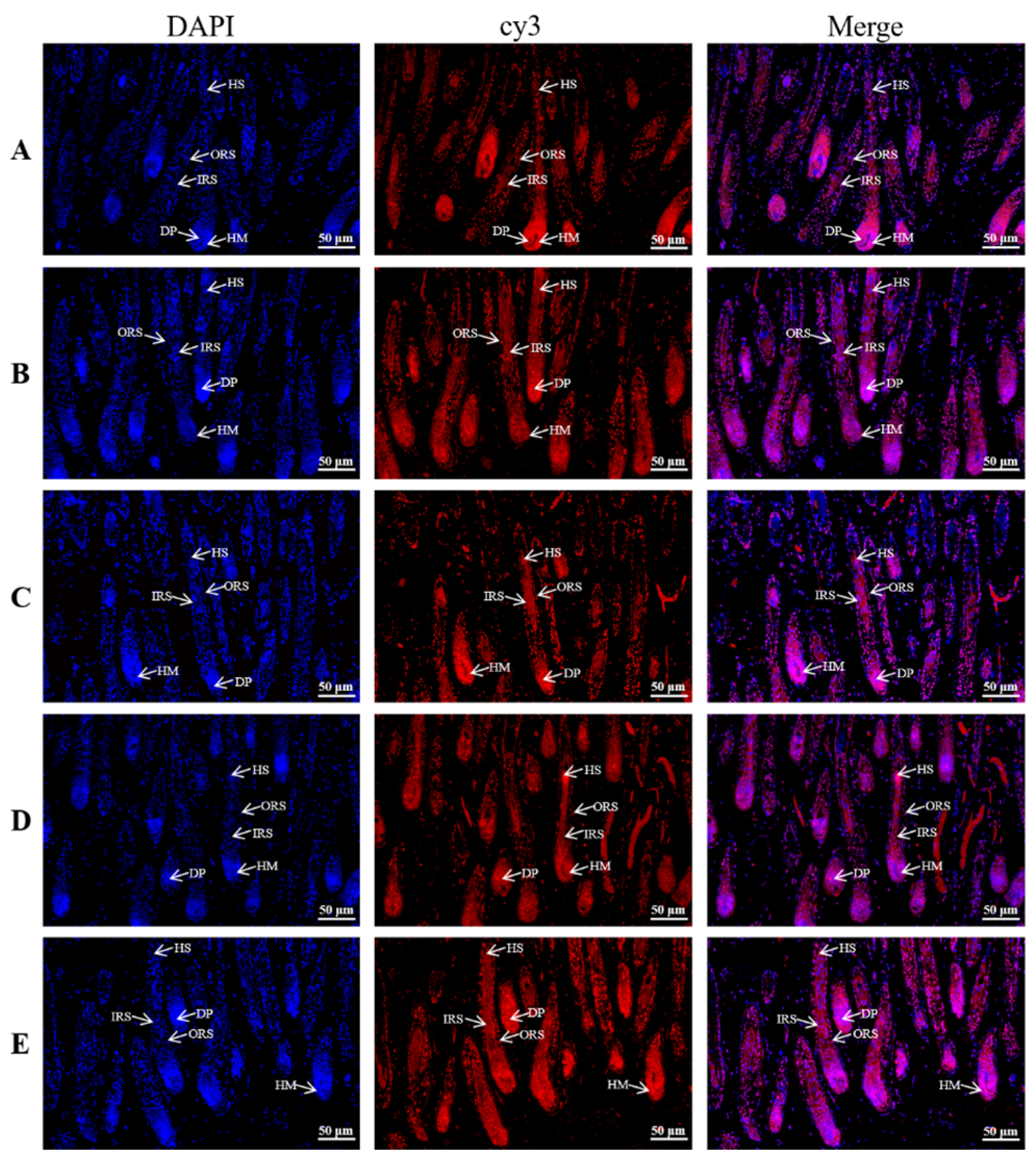
3.3. Distribution and Localization of KRTAP6 Family in 180 Days of Hair Follicle Cycle Development in the Skin
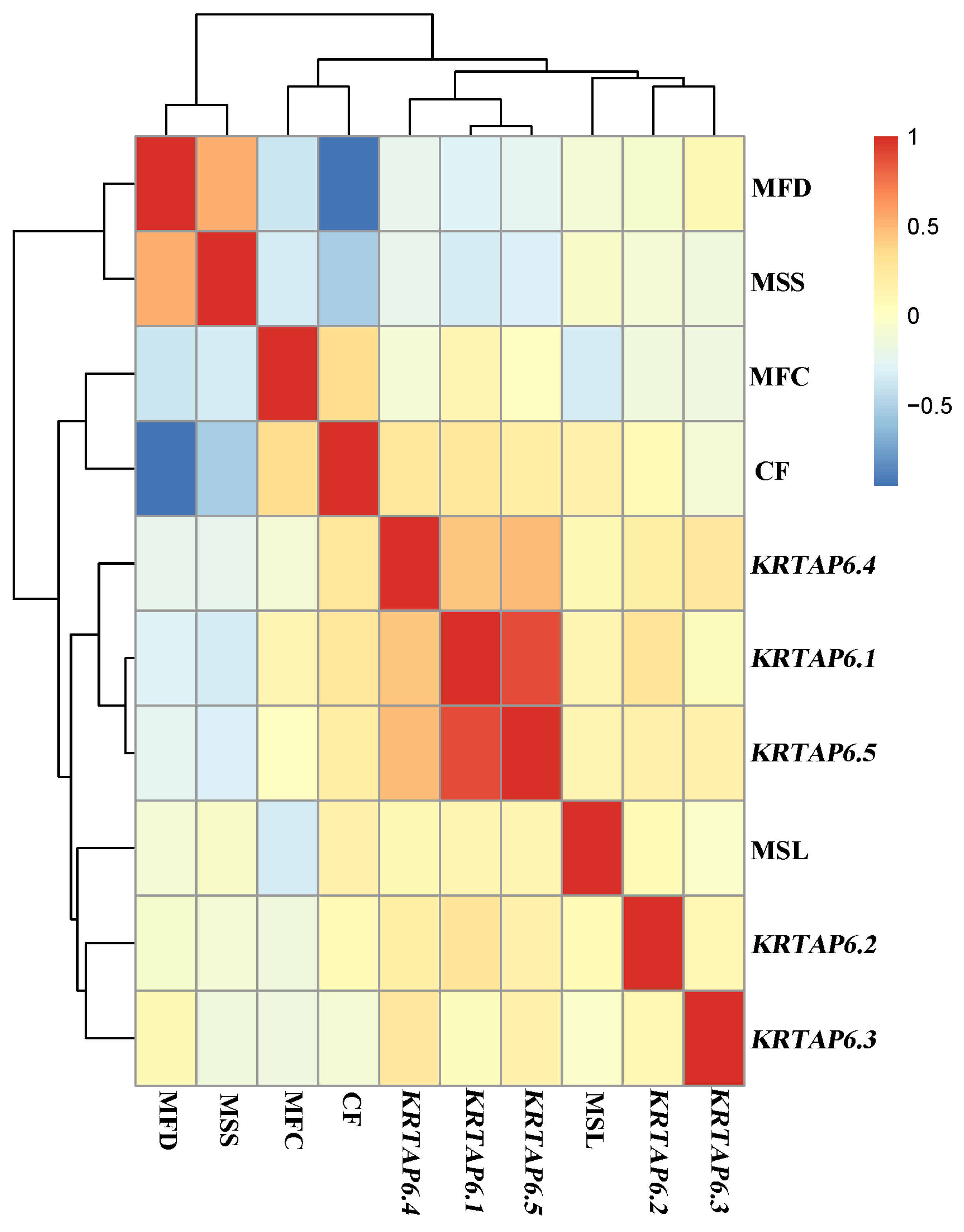
3.4. Association Analysis of mRNA Expression Levels of KRTAP6 Family with Wool Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laitala, K.; Klepp, I.; Henry, B.J.S. Does use matter? Comparison of environmental impacts of clothing based on fiber type. Sustainability 2018, 10, 2524. [Google Scholar] [CrossRef]
- Pekhtasheva, E.; Neverov, A.; Kubica, S.; Zaikov, G.J.C. Biodegradation and biodeterioration of some natural polymers. Chem. Chem. Technol. 2012, 6, 263–280. [Google Scholar] [CrossRef]
- Allafi, F.; Hossain, M.S.; Lalung, J.; Shaah, M.; Salehabadi, A.; Ahmad, M.I.; Shadi, A. Advancements in applications of natural wool fiber: Review. J. Nat. Fibers 2022, 19, 497–512. [Google Scholar] [CrossRef]
- Mcneil, S.J.; Sunderland, M.R.; Leighs, S.J. The Utilisation of Wool as a Catalyst and as a Support for Catalysts. Appl. Catal. A Gen. 2017, 541, 120–140. [Google Scholar] [CrossRef]
- Schmidt, A.C.; Jensen, A.A.; Clausen, A.U.; Kamstrup, O.; Postlethwaite, D. A comparative life cycle assessment of building insulation products made of stone wool, paper wool and flax. Int. J. Life Cycle Assess. 2004, 9, 122. [Google Scholar] [CrossRef]
- Mansour, E. Wool Fibres for the Sorption of Volatile Organic Compounds (VOCs) from Indoor Air. Ph.D. Thesis, Bangor University, Bangor, UK, 2018. [Google Scholar]
- Hanzlíková, Z.; Braniša, J.; Jomová, K.; Fülöp, M.; Hybler, P.; Porubská, M. Electron beam irradiated sheep wool—prospective sorbent for heavy metals in wastewater. Sep. Purif. Technol. 2018, 193, 345–350. [Google Scholar] [CrossRef]
- Marshall, R.C.; Orwin, D.F.; Gillespie, J.M. Structure and biochemistry of mammalian hard keratin. Electron. Microsc. Rev. 1991, 4, 47–83. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.M.; Ko, J.W.; Choi, C.W.; Lee, Y.; Seo, Y.-J.; Lee, J.-H.; Kim, C.-D. Deficiency of crif1 in hair follicle stem cells retards hair growth cycle in adult mice. PLoS ONE 2020, 15, e0232206. [Google Scholar] [CrossRef]
- Stenn, K.S.; Paus, R. Controls of hair follicle cycling. Physiol. Rev. 2001, 81, 449–494. [Google Scholar] [CrossRef]
- Müller-Röver, S.; Foitzik, K.; Paus, R.; Handjiski, B.; van der Veen, C.; Eichmüller, S.; McKay, I.A.; Stenn, K.S. A comprehensive guide for the accurate classification of murine hair follicles in distinct hair cycle stages. J. Investig. Dermatol. 2001, 117, 3–15. [Google Scholar] [CrossRef]
- Yang, K.; Tang, Y.; Ma, Y.; Liu, Q.; Huang, Y.; Zhang, Y.; Shi, X.; Zhang, L.; Zhang, Y.; Wang, J.; et al. Hair growth promoting effects of 650 nm red light stimulation on human hair follicles and study of its mechanisms via RNA sequencing transcriptome analysis. Ann. Dermatol. 2021, 33, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Grymowicz, M.; Rudnicka, E.; Podfigurna, A.; Napierala, P.; Smolarczyk, R.; Smolarczyk, K.; Meczekalski, B. Hormonal effects on hair follicles. Int. J. Mol. Sci. 2020, 21, 5342. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, R.; Wen, L.; Fan, Z.; Guo, Y.; Hu, Z.; Miao, Y. Recent progress in the understanding of the effect of sympathetic nerves on hair follicle growth. Front. Cell Dev. Biol. 2021, 26, 736738. [Google Scholar] [CrossRef]
- Elliott, K.; Stephenson, T.J.; Messenger, A.G. Differences in hair follicle dermal papilla volume are due to extracellular matrix volume and cell number: Implications for the control of hair follicle size and androgen responses. J. Investig. Dermatol. 1999, 113, 873–877. [Google Scholar] [CrossRef] [PubMed]
- Chi, W.; Wu, E.; Morgan, B.A. Dermal papilla cell number specifies hair size, shape and cycling and its reduction causes follicular decline. Development 2013, 140, 1676–1683. [Google Scholar] [CrossRef]
- Mou, C.; Thomason, H.A.; Willan, P.M.; Clowes, C.; Harris, W.E.; Drew, C.F.; Dixon, J.; Dixon, M.J.; Headon, D.J. Enhanced ectodysplasin-A receptor (EDAR) signaling alters multiple fiber characteristics to produce the East Asian hair form. Hum. Mutat. 2008, 29, 1405–1411. [Google Scholar] [CrossRef]
- Fujikawa, H.; Fujimoto, A.; Farooq, M.; Ito, M.; Shimomura, Y. Characterization of the human hair keratin–associated protein 2 (krtap2) gene family. J. Investig. Dermatol. 2012, 132, 1806–1813. [Google Scholar] [CrossRef]
- Shimomura, Y.; Ito, M. Human hair keratin-associated proteins. J. Investig. Dermatol. Symp. Proc. 2005, 10, 230–233. [Google Scholar] [CrossRef]
- Powell, B.C.; Rogers, G.E. The role of keratin proteins and their genes in the growth, structure and properties of hair. EXS 1997, 78, 59–148. [Google Scholar]
- Popescu, C.; Hcker, H. Hair—The most sophisticated biological composite material. Chem. Soc. Rev. 2007, 36, 1282–1291. [Google Scholar] [CrossRef]
- Gillespie, J.M.J.S.U. The proteins of hair and other hard α-keratins. In Cellular and Molecular Biology of Intermediate Filaments; Springer: Boston, MA, USA, 1990; pp. 95–128. [Google Scholar]
- Powell, B.C.; Beltrame, J.S. Characterization of a hair (wool) keratin intermediate filament gene domain. J. Investig. Dermatol. 1994, 102, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A.; Langbein, L.; Praetzel-Wunder, S.; Winter, H.; Cytology, J.S.J.I.R.o. Human hair keratin-associated proteins (kaps). Int. Rev. Cytol. 2006, 251, 209. [Google Scholar]
- Zhou, H.; Gong, H.; Wang, J.; Luo, Y.; Li, S.; Tao, J.; Hickford, J.G.H. The complexity of the ovine and caprine keratin-associated protein genes. Int. J. Mol. Sci. 2021, 22, 12838. [Google Scholar] [CrossRef]
- Fratini, A.; Powell, B.C.; Rogers, G.E. Sequence, expression, and evolutionary conservation of a gene encoding a glycine/tyrosine-rich keratin-associated protein of hair. J. Biol. Chem. 1993, 268, 4511–4518. [Google Scholar] [CrossRef]
- Zhou, H.; Gong, H.; Li, S.; Luo, Y.; Hickford, J.G. A 57-bp deletion in the ovine kap6-1 gene affects wool fibre diameter. J. Anim. Breed. Genet. 2015, 132, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhou, H.; Gong, H.; Zhao, F.; Wang, J.; Luo, Y.; Hickford, J.G.H. Variation in the ovine kap6-3 gene (krtap6-3) is associated with variation in mean fibre diameter-associated wool traits. Genes 2017, 8, 204. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Li, C.; Li, Y.; Zhou, G.; Gao, Y.; Ma, S.; Chen, Y.; Song, J.; Wang, X. Whole-genome bisulfite sequencing of goat skins identifies signatures associated with hair cycling. BMC Genom. 2018, 19, 638. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Bao, Z.; Zhao, B.; Zhou, T.; Li, J.; Liu, M.; Hu, S.; Yang, N.; Chen, Y.; Wu, X. Characterization and functional analysis of krtap11-1 during hair follicle development in angora rabbits (oryctolagus cuniculus). Genes Genom. 2020, 42, 1281–1290. [Google Scholar] [CrossRef]
- Shanhe, W.; Wei, G.; Zhixin, L.; Yang, G.; Beilei, J.; Lei, Q.; Zhiying, Z.; Xin, W.J.B.G. Integrated analysis of coding genes and non-coding rnas during hair follicle cycle of cashmere goat (capra hircus). BMC Genom. 2017, 18, 767. [Google Scholar]
- Shanhe, W.; Zhixin, L.; Yuelang, Z.; Dan, Y.; Wei, G.; Xin, W.J.B.G. The inconsistent regulation of hoxc13 on different keratins and the regulation mechanism on hoxc13 in cashmere goat (capra hircus). BMC Genom. 2018, 19, 630. [Google Scholar]
- Zhao, Z.; Liu, G.; Li, X.; Huang, J.; Xiao, Y.; Du, X.; Yu, M. Characterization of the promoter regions of two sheep keratin-associated protein genes for hair cortex-specific expression. PLoS ONE 2016, 11, e0153936. [Google Scholar] [CrossRef]
- Plowman, J.E.; Paton, L.N.; Bryson, W.G. The differential expression of proteins in the cortical cells of wool and hair fibres. Exp. Dermatol. 2007, 16, 707–714. [Google Scholar] [CrossRef]
- Yu, Z.; Gordon, S.W.; Nixon, A.J.; Bawden, C.S.; Rogers, M.A.; Wildermoth, J.E.; Maqbool, N.J.; Pearson, A.J. Expression patterns of keratin intermediate filament and keratin associated protein genes in wool follicles. Differentiation 2009, 77, 307–316. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Forrest, R.H.; Li, S.; Wang, J.; Dyer, J.M.; Luo, Y.; Hickford, J.G. Wool keratin-associated protein genes in sheep-a review. Genes 2016, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Alibardi, L. Ultrastructural localization of hair keratins, high sulfur keratin-associated proteins and sulfhydryl oxidase in the human hair. Anat. Sci. Int. 2017, 92, 248–261. [Google Scholar] [CrossRef]
- Rogers, M.A.; Winter, H.; Langbein, L.; Wollschläger, A.; Praetzel-Wunder, S.; Jave-Suarez, L.F.; Schweizer, J. Characterization of human kap24.1, a cuticular hair keratin-associated protein with unusual amino-acid composition and repeat structure. J. Investig. Dermatol. 2007, 127, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Powell, B.C.; Nesci, A.; Rogers, G.E. Regulation of keratin gene expression in hair follicle differentiation. Ann. N. Y. Acad. Sci. 1991, 642, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Chen, M.; Li, S.; Dai, C.; Chen, Y. Study on structure and properties of hu sheep wool. J. Nat. Fibers 2023, 20, 2160405. [Google Scholar] [CrossRef]
- Sitotaw, D.B.; Tesema, A.F.; Woldemariam, A.H. Diameter and strength of ethiopian pure and cross-breed sheep wool fibers. J. Eng. Fibers Fabr. 2021, 16, 15589250211046931. [Google Scholar] [CrossRef]
- Beh, K.J.; Callaghan, M.J.; Leish, Z.; Hulme, D.J.; Maddox, J.F.J.W.T.; Breeding, S. A genome scan for qtl affecting fleece and wool traits in merino sheep. Wool Technol. Sheep Breed. 2001, 49, 88–97. [Google Scholar]
- Cao, J.; Wang, J.; Zhou, H.; Hu, J.; Liu, X.; Li, S.; Luo, Y.; Hickford, J.G.H. Sequence variation in caprine krtap6-2 affects cashmere fiber diameter. Animals 2022, 12, 2040. [Google Scholar] [CrossRef]
- Tao, J.; Zhou, H.; Gong, H.; Yang, Z.; Ma, Q.; Cheng, L.; Ding, W.; Li, Y.; Hickford, J. Variation in the kap6-1 gene in chinese tan sheep and associations with variation in wool traits. Small Rumin. Res. 2017, 154, 129–132. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probe Name | Probe Sequence (5′–3′) | Probe Concentration (nM) | Hybridization Temperature (°C) |
|---|---|---|---|
| KRTAP6-1 | GAGGGCACGAAGCAAGTCTTTAGTAGG | 500 | 40 |
| KRTAP6-2 | GTTCAGTATACGTGGTGTTCAGAACTGG | 500 | 40 |
| KRTAP6-3 | ATGATGCAGCCTAACCATCACCCAGGG | 500 | 40 |
| KRTAP6-4 | TGTAGTGTTGTGCATCATCTCAGGGT | 500 | 40 |
| KRTAP6-5 | GTTAAAAGGTAGGCTAGGAATAGTCCA | 500 | 40 |
| Gene | GenBank Accession No. | Primer Sequence (5′–3′) | Product Length (bp) | Annealing Temperature (◦C) |
|---|---|---|---|---|
| KRTAP6-1 | NM_001193399.1 | F: TGCATGGAAGTCAAAAGAGAGTT | 154 | 60 |
| R: CCTTCCGGTGCCCTCTACAT | ||||
| KRTAP6-2 | KT725832.1 | F: CCTCGGCTGTGGAAGCTAT | 174 | 60 |
| R: TAGCCACAGCCATAGAGAGG | ||||
| KRTAP6-3 | MF061690.1 | F: CCTCGGCTGTGGAAGCTAT | 128 | 60 |
| R: CAGAGCCATAGCCGCATC | ||||
| KRTAP6-4 | KT725840.1 | F: TCTCCTCCATCCAAGAACAACC | 155 | 60 |
| R: TCCATAGCCATAGCTTCCACAG | ||||
| KRTAP6-5 | KT725846.1 | F: CCTTTTTCCCTGGGTGATGC | 199 | 60 |
| R: GACAAAGACAGCATGGAAGGC | ||||
| β–actin | NM_001009784 | F: AGCCTTCCTTCCTGGGCATGGA | 113 | 60 |
| R: GGACAGCACCGTGTTGGCGTAGA |
| Days | Corneum | Inner Root Sheath | Outer Root Sheath | Hair Medulla | Dermal Papilla |
|---|---|---|---|---|---|
| KRTAP6-1 | ++ | +++ | +++ | + | ++++ |
| KRTAP6-2 | − | +++ | ++++ | − | ++++ |
| KRTAP6-3 | − | +++ | ++++ | − | ++++ |
| KRTAP6-4 | − | +++ | ++++ | − | ++++ |
| KRTAP6-5 | − | +++ | ++++ | − | ++++ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, H.; He, Z.; Zhao, F.; Hu, J.; Wang, J.; Liu, X.; Zhao, Z.; Li, M.; Luo, Y.; Li, S. Spatiotemporal Expression Characterization of KRTAP6 Family Genes and Its Effect on Wool Traits. Genes 2024, 15, 95. https://doi.org/10.3390/genes15010095
Sun H, He Z, Zhao F, Hu J, Wang J, Liu X, Zhao Z, Li M, Luo Y, Li S. Spatiotemporal Expression Characterization of KRTAP6 Family Genes and Its Effect on Wool Traits. Genes. 2024; 15(1):95. https://doi.org/10.3390/genes15010095
Chicago/Turabian StyleSun, Hongxian, Zhaohua He, Fangfang Zhao, Jiang Hu, Jiqing Wang, Xiu Liu, Zhidong Zhao, Mingna Li, Yuzhu Luo, and Shaobin Li. 2024. "Spatiotemporal Expression Characterization of KRTAP6 Family Genes and Its Effect on Wool Traits" Genes 15, no. 1: 95. https://doi.org/10.3390/genes15010095