
Assessment of the Genetic Diversity and Structure of the Korean Endemic Freshwater Fish Microphysogobio longidorsalis (Gobioninae) Using Microsatellite Markers: A First Glance from Population Genetics


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and DNA Extraction
2.2. Whole-Genome Sequencing for Microsatellite Screening
2.3. Microsatellite Genotyping
2.4. Microsatellite Genetic Diversity Analyses
2.5. Population Genetic Structure Analysis
3. Results
3.1. Microsatellite Genetic Diversity
3.2. Bottleneck Analysis
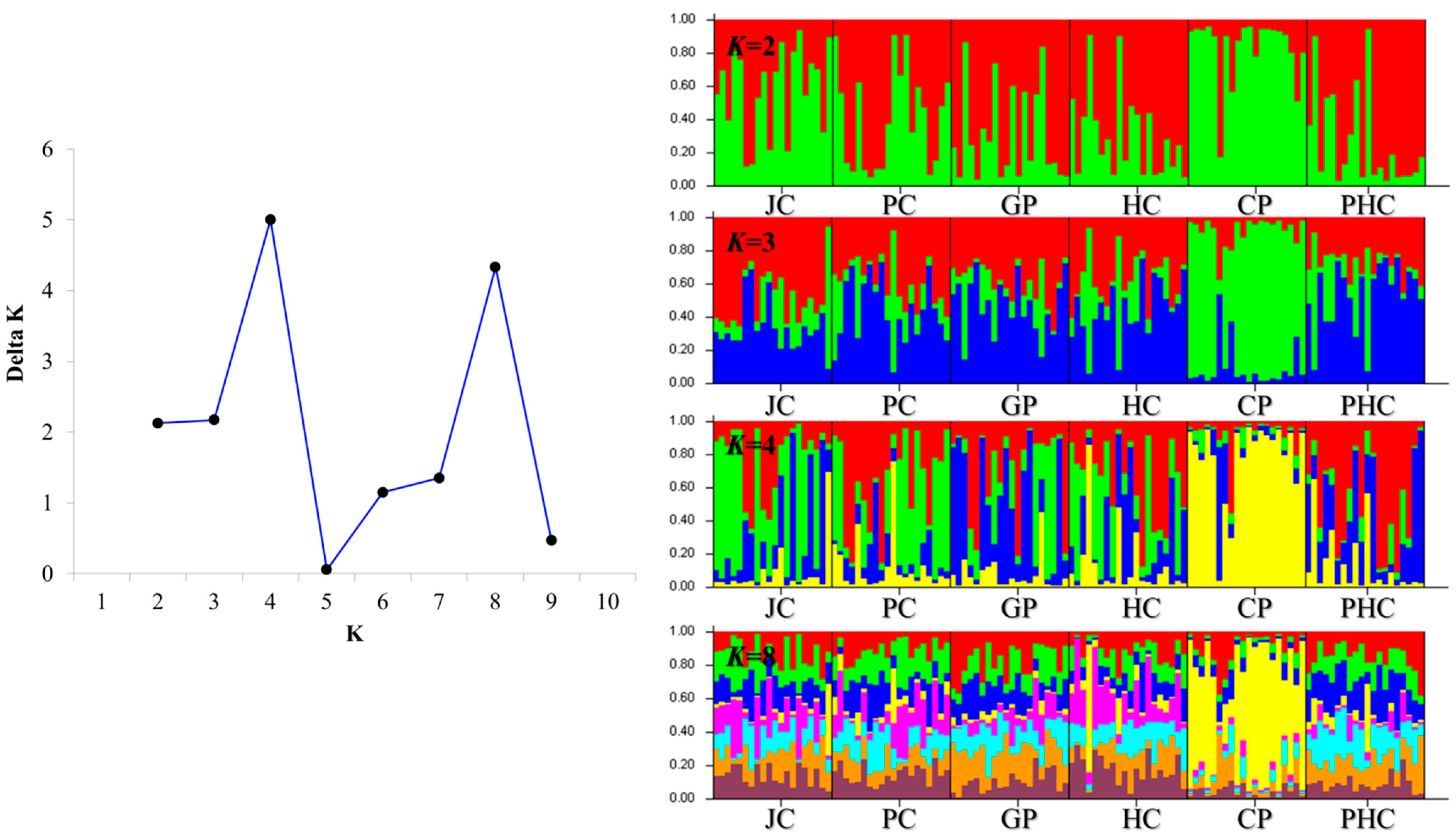
3.3. Population Structure and Genetic Differentiation Analyses
4. Discussion
4.1. Genetic Diversity and Population Bottleneck
4.2. Genetic Structure Populations
4.3. Conservation Implications
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Frankham, R. Conservation genetics. Annu. Rev. Genet. 1995, 29, 305–327. [Google Scholar] [CrossRef]
- Buj, I.; Marčić, Z.; Flauder, E.; Šanda, R.; Vukić, J. Population genetic structure of endemic fish species facilitating their survival in changing environments—A case study on the genus Telestes in Croatia. Diversity 2022, 14, 529. [Google Scholar] [CrossRef]
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344, 1246752. [Google Scholar] [CrossRef]
- Frankham, R. Inbreeding and extinction: Island populations. Conserv. Biol. 1998, 12, 665–675. [Google Scholar] [CrossRef]
- Furlan, E.; Stoklosa, J.; Griffiths, J.; Gust, N.; Ellis, R.; Huggins, R.M.; Weeks, A.R. Small population size and extremely low levels of genetic diversity in island populations of the platypus, Ornithorhynchus anatinus. Ecol. Evol. 2012, 2, 844–857. [Google Scholar] [CrossRef]
- Whitlock, M.C. Fixation of new alleles and the extinction of small populations: Drift load, beneficial alleles, and sexual selection. Evolution 2000, 54, 1855–1861. [Google Scholar] [CrossRef]
- Frankham, R.; Ballou, J.; Briscoe, D. Introduction to Conservation Genetics, 2nd ed.; Cambridge University Press: New York, NY, USA, 2010. [Google Scholar]
- Barnosky, A.D.; Matzke, N.; Tomiya, S.; Wogan, G.O.; Swartz, B.; Quental, T.B.; Marshall, C.; McGuire, J.L.; Lindsey, E.L.; Maguire, K.C.; et al. Has the Earth’s sixth mass extinction already arrived? Nature 2011, 471, 51–57. [Google Scholar] [CrossRef]
- Maxwell, S.L.; Fuller, R.A.; Brooks, T.M.; Watson, J.E. Biodiversity: The ravages of guns, nets and bulldozers. Nature 2016, 536, 143–145. [Google Scholar] [CrossRef]
- Markert, J.A.; Champlin, D.M.; Gutjahr-Gobell, R.; Grear, J.S.; Kuhn, A.; McGreevy, T.J.; Roth, A.; Bagley, M.J.; Nacci, D.E. Population genetic diversity and fitness in multiple environments. BMC Evol. Biol. 2010, 10, 205. [Google Scholar] [CrossRef]
- Frankham, R. Genetics and extinction. Biol. Conserv. 2005, 126, 131–140. [Google Scholar] [CrossRef]
- Brauer, C.J.; Beheregaray, L.B. Recent and rapid anthropogenic habitat fragmentation increases extinction risk for freshwater biodiversity. Evol. Appl. 2020, 13, 2857–2869. [Google Scholar] [CrossRef]
- Gurevitch, J.; Padilla, D.K. Are invasive species a major cause of extinctions? Trends Ecol. Evol. 2004, 19, 470–474. [Google Scholar] [CrossRef]
- Jeon, H.B.; Song, H.Y.; Suk, H.Y.; Bang, I.C. Phylogeography of the Korean endemic Coreoleuciscus (Cypriniformes: Gobionidae): The genetic evidence of colonization through Eurasian continent to the Korean Peninsula during Late Plio-Pleistocene. Genes Genom. 2022, 44, 709–719. [Google Scholar] [CrossRef]
- López-Pujol, J.; Martinell, M.C.; Massó, S.; Blanché, C.; Sáez, L. The ‘paradigm of extremes’: Extremely low genetic diversity in an extremely narrow endemic species, Coristospermum huteri (Umbelliferae). Plant Syst. Evol. 2013, 299, 439–446. [Google Scholar] [CrossRef]
- Kim, I.S.; Choi, Y.; Lee, C.L.; Lee, Y.J.; Kim, B.J.; Kim, J.H. Illustrated Book of Korean Fishes; Kyo-Hak Publishing Co.: Seoul, Republic of Korea, 2005; p. 615. [Google Scholar]
- Hong, Y.K.; Sung, H.C.; Ko, M.H.; Kim, K.S.; Bang, I.C. Distribution status and habitat characteristics of the endangered freshwater fish, Microphysogobio rapidus (Cyprinidae). Anim. Cells Syst. 2017, 21, 286–293. [Google Scholar] [CrossRef]
- Palsbøll, P.J.; Berube, M.; Allendorf, F.W. Identification of management units using population genetic data. Trends Ecol. Evol. 2007, 22, 11–16. [Google Scholar] [CrossRef]
- Kim, D.Y.; Suk, H.Y. Genetic diversity of the slender shinner (Pseudopuntungia tenuicorpa) and its conservational implications. Korean J. Ichthyol. 2020, 32, 39–48. [Google Scholar] [CrossRef]
- Hong, Y.K.; Kim, K.R.; Kim, K.S.; Bang, I.C. The impact of weir construction in Korea’s Nakdong River on the population genetic variability of the endangered fish species, rapid small gudgeon (Microphysogobio rapidus). Genes 2023, 14, 1611. [Google Scholar] [CrossRef]
- Guo, X.Z.; Chen, H.M.; Wang, A.B.; Qian, X.Q. Population genetic structure of the yellow catfish (Pelteobagrus fulvidraco) in China inferred from microsatellite analyses: Implications for fisheries management and breeding. J. World Aquacult. Soc. 2022, 53, 174–191. [Google Scholar] [CrossRef]
- Kim, K.R.; Kwak, Y.H.; Sung, M.S.; Cho, S.J.; Bang, I.C. Population structure and genetic diversity of the endangered fish black shinner Pseudopungtungia nigra (Cyprinidae) in Korea: A wild and restoration population. Sci. Rep. 2023, 13, 9692. [Google Scholar] [CrossRef]
- Wright, J.M.; Bentzen, P. Microsatellites: Genetic markers for the future. In Molecular Genetics in Fisheries; Springer: Dordrecht, The Netherlands, 1995; pp. 1121–1171. [Google Scholar]
- Hodel, R.G.; Segovia-Salcedo, M.C.; Landis, J.B.; Crowl, A.A.; Sun, M.; Liu, X.; Gitzendanner, M.A.; Douglas, N.A.; Germain-Aubrey, C.C.; Chen, S.; et al. The report of my death was an exaggeration: A review for researchers using microsatellites in the 21st century. Appl. Plant Sci. 2016, 4, 1600025. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Li, H. Aligning Sequence Reads, Clone Sequences and Assembly Contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Schuelke, M. An economic method for the fluorescent labeling of PCR fragments. Nat. Biotechnol. 2000, 18, 233–234. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol. Ecol. 2007, 16, 1099–1106. [Google Scholar] [CrossRef]
- Raymond, M.; Rousset, F. Population genetics software for exact test and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Piry, S.; Luikart, G.; Cornuet, J.M. Computer note. BOTTLENECK: A computer program for detecting recent reductions in the effective size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Maruyama, T.; Fuerst, P.A. Population bottlenecks and nonequilibrium models in population genetics. II. Number of alleles in a small population that was formed by a recent bottleneck. Genetics 1985, 111, 675–689. [Google Scholar] [CrossRef]
- Cornuet, J.M.; Luikart, G. Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data. Genetics 1996, 144, 2001–2014. [Google Scholar] [CrossRef]
- Luikart, G.; Cornuet, J.M. Empirical evaluation of a test for identifying recently bottlenecked populations from allele frequency data. Conserv. Biol. 1998, 12, 228–237. [Google Scholar] [CrossRef]
- Do, C.; Waples, R.S.; Peel, D.; Macbeth, G.M.; Tillett, B.J.; Ovenden, J.R. NeEstimator v2: Re-implementation of software for the estimation of contemporary effective population size (Ne) from genetic data. Mol. Ecol. Resour. 2014, 14, 209–214. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Earl, D.A.; VonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jombart, T. Adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef]
- Kim, K.R.; Choi, H.K.; Lee, T.W.; Lee, H.J.; Yu, J.N. Population structure and genetic diversity of the spotted sleeper Odontobutis interrupta (Odontobutidae), a fish endemic to Korea. Diversity 2023, 15, 913. [Google Scholar] [CrossRef]
- Weeks, A.R.; Sgro, C.M.; Young, A.G.; Frankham, R.; Mitchell, N.J.; Miller, K.A.; Byrne, M.; Coates, D.J.; Eldridge, M.D.; Sunnucks, P.; et al. Assessing the benefits and risks of translocations in changing environments: A genetic perspective. Evol. Appl. 2011, 4, 709–725. [Google Scholar] [CrossRef] [PubMed]
- Aitken, S.N.; Bemmels, J.B. Time to get moving: Assisted gene flow of forest trees. Evol. Appl. 2016, 9, 271–290. [Google Scholar] [CrossRef]
- Miller, J.M.; Cullingham, C.I.; Peery, R.M. The influence of a priori grouping on inference of genetic clusters: Simulation study and literature review of the DAPC method. Heredity 2020, 125, 269–280. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Water System | N | NA | HO | HE | PHWE | FIS |
|---|---|---|---|---|---|---|---|
| JC | Namhangang River | 20 | 8.47 | 0.625 | 0.741 | 0.000 | 0.132 *** |
| PC | Namhangang River | 20 | 9.00 | 0.683 | 0.775 | 0.000 | 0.092 *** |
| GP | Bukhangang River | 20 | 9.16 | 0.660 | 0.773 | 0.000 | 0.129 *** |
| HC | Bukhangang River | 20 | 9.47 | 0.668 | 0.757 | 0.000 | 0.141 *** |
| CP | Bukhangang River | 20 | 7.74 | 0.675 | 0.766 | 0.000 | 0.079 *** |
| PHC | Imjingang River | 20 | 9.32 | 0.650 | 0.779 | 0.000 | 0.123 *** |
| Population ID | N | Wilcoxon Signed-Rank Test | Ne | (95% CI) | |||
|---|---|---|---|---|---|---|---|
| PIAM | PTPM | PSMM | Mode-Shift | ||||
| JC | 20 | 0.000 *** | 0.059 | 0.275 | L-shaped | 688 | (101–∞) |
| PC | 20 | 0.007 ** | 0.290 | 0.600 | L-shaped | 503 | (99–∞) |
| GP | 20 | 0.004 ** | 0.152 | 0.533 | L-shaped | 196 | (75–∞) |
| HC | 20 | 0.001 ** | 0.173 | 0.483 | L-shaped | ∞ | (68–∞) |
| CP | 20 | 0.000 *** | 0.044 * | 0.132 | Shifted | 78 | (48–212) |
| PHC | 20 | 0.000 *** | 0.033 * | 0.142 | L-shaped | ∞ | (54–∞) |
| JC | PC | GP | HC | CP | PHC | |
|---|---|---|---|---|---|---|
| JC | - | 0.018 | 0.000 | 0.000 | 0.000 | 0.000 |
| PC | 0.013 | - | 0.054 | 0.000 | 0.000 | 0.054 |
| GP | 0.020 | 0.010 | - | 0.000 | 0.000 | 0.243 |
| HC | 0.034 | 0.015 | 0.019 | - | 0.000 | 0.153 |
| CP | 0.035 | 0.027 | 0.041 | 0.038 | - | 0.000 |
| PHC | 0.026 | 0.011 | 0.007 | 0.008 | 0.036 | - |
| K | Mean LnP (K) |
|---|---|
| 1 | −8928.47 |
| 2 | −8930.61 |
| 3 | −9034.84 |
| 4 | −8931.72 |
| 5 | −9091.46 |
| 6 | −9240.74 |
| 7 | −9546.24 |
| 8 | −9198.53 |
| 9 | −9453.05 |
| 10 | −9585.59 |
| Source of Variation | d.f. | Sum of Squares | Variance Components | Percentage of Variance | F-Statistics |
|---|---|---|---|---|---|
| Microsatellite (two groups based on the water system (Namhangang River: JC, PC vs. Bukhangang and Imjingang Rivers: GP, HC, CP, PHC) | |||||
| Among groups | 1 | 11.787 | 0.00618 | 0.10 | 0.023 *** |
| Among populations within groups | 4 | 44.513 | 0.13287 | 2.23 | 0.022 *** |
| Within populations | 234 | 1360.300 | 5.81325 | 97.66 | 0.001 * |
| Total | 239 | 1416.600 | 5.95230 | 100.00 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, K.-R.; Sung, M.-S.; Hwang, Y.; Jeong, J.H.; Yu, J.-N. Assessment of the Genetic Diversity and Structure of the Korean Endemic Freshwater Fish Microphysogobio longidorsalis (Gobioninae) Using Microsatellite Markers: A First Glance from Population Genetics. Genes 2024, 15, 69. https://doi.org/10.3390/genes15010069
Kim K-R, Sung M-S, Hwang Y, Jeong JH, Yu J-N. Assessment of the Genetic Diversity and Structure of the Korean Endemic Freshwater Fish Microphysogobio longidorsalis (Gobioninae) Using Microsatellite Markers: A First Glance from Population Genetics. Genes. 2024; 15(1):69. https://doi.org/10.3390/genes15010069
Chicago/Turabian StyleKim, Kang-Rae, Mu-Sung Sung, Yujin Hwang, Ju Hui Jeong, and Jeong-Nam Yu. 2024. "Assessment of the Genetic Diversity and Structure of the Korean Endemic Freshwater Fish Microphysogobio longidorsalis (Gobioninae) Using Microsatellite Markers: A First Glance from Population Genetics" Genes 15, no. 1: 69. https://doi.org/10.3390/genes15010069