
Idiopathic Epilepsy Risk Allele Trends in Belgian Tervuren: A Longitudinal Genetic Analysis
 , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ekenstedt, K.J.; Patterson, E.E.; Mickelson, J.R. Canine Epilepsy Genetics. Mamm. Genome 2012, 23, 28–39. [Google Scholar] [CrossRef]
- Knowles, K. Idiopathic Epilepsy. Clin. Tech. Small Anim. Pract. 1998, 13, 144–151. [Google Scholar] [CrossRef]
- Weber, Y.G.; Lerche, H. Genetic Mechanisms in Idiopathic Epilepsies. Dev. Med. Child Neurol. 2008, 50, 648–654. [Google Scholar] [CrossRef]
- Jallon, P.; Latour, P. Epidemiology of Idiopathic Generalized Epilepsies. Epilepsia 2005, 46, 10–14. [Google Scholar] [CrossRef] [PubMed]
- De Risio, L.; Bhatti, S.; Muñana, K.; Penderis, J.; Stein, V.; Tipold, A.; Berendt, M.; Farqhuar, R.; Fischer, A.; Long, S.; et al. International Veterinary Epilepsy Task Force Consensus Proposal: Diagnostic Approach to Epilepsy in Dogs. BMC Vet. Res. 2015, 11, 148. [Google Scholar] [CrossRef]
- Folkard, E.; Niel, L.; Gaitero, L.; James, F.M.K. Tools and Techniques for Classifying Behaviours in Canine Epilepsy. Front. Vet. Sci. 2023, 10, 1211515. [Google Scholar] [CrossRef]
- Kearsley-Fleet, L.; O’Neill, D.G.; Volk, H.A.; Church, D.B.; Brodbelt, D.C. Prevalence and Risk Factors for Canine Epilepsy of Unknown Origin in the UK. Vet. Rec. 2013, 172, 338. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, R.; Adams, V.; Keys, D.; Platt, S.R. Canine Idiopathic Epilepsy: Prevalence, Risk Factors and Outcome Associated with Cluster Seizures and Status Epilepticus. J. Small Anim. Pract. 2012, 53, 526–530. [Google Scholar] [CrossRef]
- Heske, L.; Berendt, M.; Jäderlund, K.H.; Egenvall, A.; Nødtvedt, A. Validation of the Diagnosis Canine Epilepsy in a Swedish Animal Insurance Database against Practice Records. Prev. Vet. Med. 2014, 114, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Berendt, M.; Gulløv, C.H.; Christensen, S.L.K.; Gudmundsdottir, H.; Gredal, H.; Fredholm, M.; Alban, L. Prevalence and Characteristics of Epilepsy in the Belgian Shepherd Variants Groenendael and Tervueren Born in Denmark 1995–2004. Acta Vet. Scand. 2008, 50, 51. [Google Scholar] [CrossRef]
- Gulløv, C.H.; Toft, N.; Berendt, M. A Longitudinal Study of Survival in Belgian Shepherds with Genetic Epilepsy. J. Vet. Intern. Med. 2012, 26, 1115–1120. [Google Scholar] [CrossRef] [PubMed]
- Famula, T.R.; Oberbauer, A.M.; Brown, K.N. Heritability of Epileptic Seizures in the Belgian Tervueren. J. Small Anim. Pract. 1997, 38, 349–352. [Google Scholar] [CrossRef]
- Pergande, A.E.; Belshaw, Z.; Volk, H.A.; Packer, R.M.A. “We Have a Ticking Time Bomb”: A Qualitative Exploration of the Impact of Canine Epilepsy on Dog Owners Living in England. BMC Vet. Res. 2020, 16, 443. [Google Scholar] [CrossRef] [PubMed]
- Wessmann, A.; Volk, H.A.; Packer, R.M.A.; Ortega, M.; Anderson, T.J. Quality-of-Life Aspects in Idiopathic Epilepsy in Dogs. Vet. Rec. 2016, 179, 229. [Google Scholar] [CrossRef]
- Hobbs, S.L.; Blackwell, E.J.; Wetz, K.E.; Packer, R.M.A. Owner Reported Management of Interictal Anxiety Behaviours in Canine Epilepsy. Vet. Rec. 2022, 190, e1321. [Google Scholar] [CrossRef] [PubMed]
- Winter, J.; Packer, R.M.A.; Volk, H.A. Preliminary Assessment of Cognitive Impairments in Canine Idiopathic Epilepsy. Vet. Rec. 2018, 182, 633. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, S.L.; Law, T.H.; Volk, H.A.; Younis, C.; Casey, R.A.; Packer, R.M.A. Impact of Canine Epilepsy on Judgement and Attention Biases. Sci. Rep. 2020, 10, 17719. [Google Scholar] [CrossRef]
- Packer, R.M.A.; Lucas, R.; Volk, H.A. Owner Perception of Focal Seizures in Canine Epilepsy. Vet. Rec. 2017, 180, 150. [Google Scholar] [CrossRef] [PubMed]
- Packer, R.M.A.; Volk, H.A. Epilepsy beyond Seizures: A Review of the Impact of Epilepsy and Its Comorbidities on Health-Related Quality of Life in Dogs. Vet. Rec. 2015, 177, 306–315. [Google Scholar] [CrossRef]
- Peek, S.I.; Meller, S.; Twele, F.; Packer, R.M.A.; Volk, H.A. Epilepsy Is More than a Simple Seizure Disorder: Parallels between Human and Canine Cognitive and Behavioural Comorbidities. Vet. J. 2024, 303, 106060. [Google Scholar] [CrossRef]
- Shihab, N.; Bowen, J.; Volk, H.A. Behavioral Changes in Dogs Associated with the Development of Idiopathic Epilepsy. Epilepsy Behav. 2011, 21, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.A. A Clinical and Diagnostic Approach to the Patient with Seizures. Top. Companion Anim. Med. 2013, 28, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, F.C.; Sica, R.E.P. Canine Electroencephalographic Recording Technique: Findings in Normal and Epileptic Dogs. Clin. Neurophysiol. 2004, 115, 477–487. [Google Scholar] [CrossRef]
- Berendt, M.; Gredal, H.; Ersbøll, A.K.; Alving, J. Premature Death, Risk Factors, and Life Patterns in Dogs with Epilepsy. J. Vet. Intern. Med. 2007, 21, 754–759. [Google Scholar] [CrossRef] [PubMed]
- Booth, S.; Meller, S.; Packer, R.M.; Farquhar, R.; Maddison, J.E.; Volk, H.A. Owner Compliance in Canine Epilepsy. Vet. Rec. 2021, 188, e16. [Google Scholar] [CrossRef]
- Arrol, L.; Penderis, J.; Garosi, L.; Cripps, P.; Gutierrez-Quintana, R.; Gonçalves, R. Aetiology and Long-Term Outcome of Juvenile Epilepsy in 136 Dogs. Vet. Rec. 2012, 170, 335. [Google Scholar] [CrossRef]
- Charalambous, M.; Brodbelt, D.; Volk, H.A. Treatment in Canine Epilepsy—A Systematic Review. BMC Vet. Res. 2014, 10, 257. [Google Scholar] [CrossRef] [PubMed]
- Casal, M.L.; Munuve, R.M.; Janis, M.A.; Werner, P.; Henthorn, P.S. Epilepsy in Irish Wolfhounds. J. Vet. Intern. Med. 2006, 20, 131–135. [Google Scholar] [CrossRef]
- Patterson, E.E.; Mickelson, J.R.; Da, Y.; Roberts, M.C.; McVey, A.S.; O’Brien, D.P.; Johnson, G.S.; Armstrong, P.J. Clinical Characteristics and Inheritance of Idiopathic Epilepsy in Vizslas. J. Vet. Intern. Med. 2003, 17, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Licht, B.G.; Lin, S.; Luo, Y.; Hyson, L.L.; Licht, M.H.; Harper, K.M.; Sullivan, S.A.; Fernandez, S.A.; Johnston, E.V. Clinical Characteristics and Mode of Inheritance of Familial Focal Seizures in Standard Poodles. J. Am. Vet. Med. Assoc. 2007, 231, 1520–1528. [Google Scholar] [CrossRef]
- Patterson, E.E. Canine Epilepsy: An Underutilized Model. ILAR J. 2014, 55, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Barker, J. Epilepsy in the Dog–a Comparative Approach. J. Small Anim. Pract. 1973, 14, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Famula, T.R.; Oberbauer, A.M. Reducing the Incidence of Epileptic Seizures in the Belgian Tervuren through Selection. Prev. Vet. Med. 1998, 33, 251–259. [Google Scholar] [CrossRef]
- Seppälä, E.H.; Koskinen, L.L.E.; Gulløv, C.H.; Jokinen, P.; Karlskov-Mortensen, P.; Bergamasco, L.; Körberg, I.B.; Cizinauskas, S.; Oberbauer, A.M.; Berendt, M.; et al. Identification of a Novel Idiopathic Epilepsy Locus in Belgian Shepherd Dogs. PLoS ONE 2012, 7, e33549. [Google Scholar] [CrossRef]
- Belanger, J.M.; Famula, T.R.; Gershony, L.C.; Palij, M.K.; Oberbauer, A.M. Genome-Wide Association Analysis of Idiopathic Epilepsy in the Belgian Shepherd. Canine Med. Genet. 2020, 7, 12. [Google Scholar] [CrossRef]
- Bithell, A.; Alberta, J.; Hornby, F.; Stiles, C.D.; Williams, B.P. Expression of the Guanine Nucleotide Exchange Factor, Mr-Gef, Is Regulated during the Differentiation of Specific Subsets of Telencephalic Neurons. Dev. Brain Res. 2003, 146, 107–118. [Google Scholar] [CrossRef]
- Winden, K.D.; Bragin, A.; Engel, J.; Geschwind, D.H. Molecular Alterations in Areas Generating Fast Ripples in an Animal Model of Temporal Lobe Epilepsy. Neurobiol. Dis. 2015, 78, 35–44. [Google Scholar] [CrossRef]
- Seppälä, E.H.; Jokinen, T.S.; Fukata, M.; Fukata, Y.; Webster, M.T.; Karlsson, E.K.; Kilpinen, S.K.; Steffen, F.; Dietschi, E.; Leeb, T.; et al. LGI2 Truncation Causes a Remitting Focal Epilepsy in Dogs. PLoS Genet. 2011, 7, e1002194. [Google Scholar] [CrossRef] [PubMed]
- Pakozdy, A.; Patzl, M.; Zimmermann, L.; Jokinen, T.S.; Glantschnigg, U.; Kelemen, A.; Hasegawa, D. LGI Proteins and Epilepsy in Human and Animals. J. Vet. Intern. Med. 2015, 29, 997–1005. [Google Scholar] [CrossRef]
- Laub, F.; Lei, L.; Sumiyoshi, H.; Kajimura, D.; Dragomir, C.; Smaldone, S.; Puche, A.C.; Petros, T.J.; Mason, C.; Parada, L.F.; et al. Transcription Factor KLF7 Is Important for Neuronal Morphogenesis in Selected Regions of the Nervous System. Mol. Cell. Biol. 2005, 25, 5699–5711. [Google Scholar] [CrossRef]
- Lei, L.; Laub, F.; Lush, M.; Romero, M.; Zhou, J.; Luikart, B.; Klesse, L.; Ramirez, F.; Parada, L.F. The Zinc Finger Transcription Factor Klf7 Is Required for TrkA Gene Expression and Development of Nociceptive Sensory Neurons. Genes Dev. 2005, 19, 1354–1364. [Google Scholar] [CrossRef]
- Caiazzo, M.; Colucci-D’Amato, L.; Esposito, M.T.; Parisi, S.; Stifani, S.; Ramirez, F.; di Porzio, U. Transcription Factor KLF7 Regulates Differentiation of Neuroectodermal and Mesodermal Cell Lineages. Exp. Cell Res. 2010, 316, 2365–2376. [Google Scholar] [CrossRef] [PubMed]
- Kajimura, D.; Dragomir, C.; Ramirez, F.; Laub, F. Identification of Genes Regulated by Transcription Factor KLF7 in Differentiating Olfactory Sensory Neurons. Gene 2007, 388, 34–42. [Google Scholar] [CrossRef]
- Belanger, J.M.; Heinonen, T.; Famula, T.R.; Mandigers, P.J.J.; Leegwater, P.A.; Hytönen, M.K.; Lohi, H.; Oberbauer, A.M. Validation of a Chromosome 14 Risk Haplotype for Idiopathic Epilepsy in the Belgian Shepherd Dog Found to Be Associated with an Insertion in the RAPGEF5 Gene. Genes 2022, 13, 1124. [Google Scholar] [CrossRef]
- Hoeppner, M.P.; Lundquist, A.; Pirun, M.; Meadows, J.R.S.; Zamani, N.; Johnson, J.; Sundström, G.; Cook, A.; FitzGerald, M.G.; Swofford, R.; et al. An Improved Canine Genome and a Comprehensive Catalogue of Coding Genes and Non-Coding Transcripts. PLoS ONE 2014, 9, e91172. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R; RStudio Team: Boston, MA, USA, 2023. [Google Scholar]
- Lampi, S.; Donner, J.; Anderson, H.; Pohjoismäki, J. Variation in Breeding Practices and Geographic Isolation Drive Subpopulation Differentiation, Contributing to the Loss of Genetic Diversity within Dog Breed Lineages. Canine Med. Genet. 2020, 7, 5. [Google Scholar] [CrossRef]
- Cramér, H. Mathematical Methods of Statistics (PMS-9), Volume 9; Princeton University Press: Princeton, NJ, USA, 1946. [Google Scholar] [CrossRef]
- Lowry, R. VassarStats: Statistical Computation Web Site. Available online: http://vassarstats.net/ (accessed on 18 December 2023).
- Fisher, R.A. Statistical Methods for Research Workers, 5th ed.; Oliver and Boyd: Edinburgh, UK, 1934. [Google Scholar]
- van der Velden, N.A. Fits in Tervueren Shepherd Dogs: A Presumed Hereditary Trait. J. Small Anim. Pract. 1968, 9, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Koskinen, L.L.E.; Seppälä, E.H.; Weissl, J.; Jokinen, T.S.; Viitmaa, R.; Hänninen, R.L.; Quignon, P.; Fischer, A.; André, C.; Lohi, H. ADAM23 Is a Common Risk Gene for Canine Idiopathic Epilepsy. BMC Genet. 2017, 18, 8. [Google Scholar] [CrossRef] [PubMed]
- Forsgård, J.A.; Metsähonkala, L.; Kiviranta, A.-M.; Cizinauskas, S.; Junnila, J.J.T.; Laitinen-Vapaavuori, O.; Jokinen, T.S. Seizure-Precipitating Factors in Dogs with Idiopathic Epilepsy. J. Vet. Intern. Med. 2019, 33, 701–707. [Google Scholar] [CrossRef]
- Gulcebi, M.I.; Bartolini, E.; Lee, O.; Lisgaras, C.P.; Onat, F.; Mifsud, J.; Striano, P.; Vezzani, A.; Hildebrand, M.S.; Jimenez-Jimenez, D.; et al. Climate Change and Epilepsy: Insights from Clinical and Basic Science Studies. Epilepsy Behav. 2021, 116, 107791. [Google Scholar] [CrossRef] [PubMed]
- Griffin, S.; Stabile, F.; De Risio, L. Cross Sectional Survey of Canine Idiopathic Epilepsy Management in Primary Care in the United Kingdom. Front. Vet. Sci. 2022, 9, 907313. [Google Scholar] [CrossRef] [PubMed]
- Potschka, H.; Fischer, A.; Löscher, W.; Volk, H.A. Pathophysiology of Drug-Resistant Canine Epilepsy. Vet. J. 2023, 296–297, 105990. [Google Scholar] [CrossRef]
- Cocostîrc, V.; Paștiu, A.I.; Pusta, D.L. An Overview of Canine Inherited Neurological Disorders with Known Causal Variants. Animals 2023, 13, 3568. [Google Scholar] [CrossRef] [PubMed]


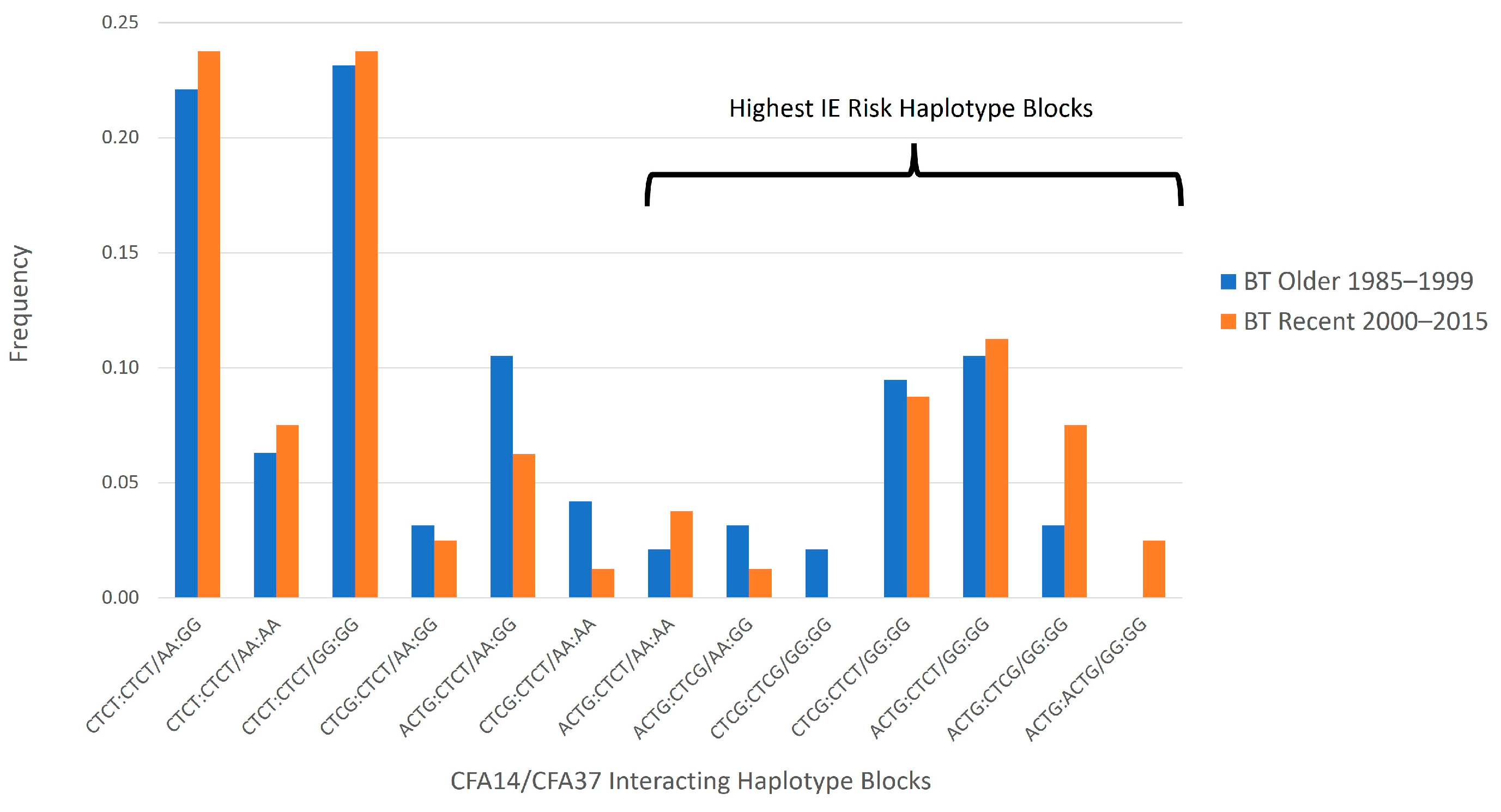{kind=link}
| CFA14—BICF2S23230472 | CFA37—BICF2P271491 | |||||
|---|---|---|---|---|---|---|
| Risk-Allele A Frequency Older Cohort | Risk-Allele A Frequency Recent Cohort | p-Value | Risk-Allele G Frequency Older Cohort | Risk-Allele G Frequency Recent Cohort | p-Value | |
| All dogs | ||||||
| Control | 0.14 (n = 85) | 0.15 (n = 66) | 0.41 | 0.62 (n = 85) | 0.73 (n = 66) | 0.03 |
| IE | 0.25 (n = 16) | 0.21 (n = 36) | 0.41 | 0.91 (n = 16) | 0.74 (n = 36) | 0.04 |
| Total | 0.15 (n = 101) | 0.17 (n = 102) | 0.36 | 0.67 (n = 101) | 0.74 (n = 102) | 0.09 |
| Unrelated dogs | ||||||
| Control | 0.13 (n = 80) | 0.14 (n = 52) | 0.45 | 0.64 (n = 80) | 0.72 (n = 52) | 0.10 |
| IE | 0.23 (n = 15) | 0.23 (n = 28) | 0.60 | 0.90 (n = 15) | 0.68 (n = 28) | 0.02 |
| Total | 0.15 (n = 95) | 0.18 (n = 80) | 0.29 | 0.68 (n = 95) | 0.71 (n = 80) | 0.33 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kinsey, N.; Belanger, J.M.; Mandigers, P.J.J.; Leegwater, P.A.; Heinonen, T.; Hytönen, M.K.; Lohi, H.; Ostrander, E.A.; Oberbauer, A.M. Idiopathic Epilepsy Risk Allele Trends in Belgian Tervuren: A Longitudinal Genetic Analysis. Genes 2024, 15, 114. https://doi.org/10.3390/genes15010114
Kinsey N, Belanger JM, Mandigers PJJ, Leegwater PA, Heinonen T, Hytönen MK, Lohi H, Ostrander EA, Oberbauer AM. Idiopathic Epilepsy Risk Allele Trends in Belgian Tervuren: A Longitudinal Genetic Analysis. Genes. 2024; 15(1):114. https://doi.org/10.3390/genes15010114
Chicago/Turabian StyleKinsey, Nathan, Janelle M. Belanger, Paul J. J. Mandigers, Peter A. Leegwater, Tiina Heinonen, Marjo K. Hytönen, Hannes Lohi, Elaine A. Ostrander, and Anita M. Oberbauer. 2024. "Idiopathic Epilepsy Risk Allele Trends in Belgian Tervuren: A Longitudinal Genetic Analysis" Genes 15, no. 1: 114. https://doi.org/10.3390/genes15010114