
Analysis of the Selection Signal of the Tibetan Black Chicken Genome Based on Whole-Genome Sequencing
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Whole-Genome Resequencing
2.2. Read Mapping, Genomic Variant Calling and Annotation
2.3. Genome-Wide Selective Sweep Test
2.4. Gene Ontology (GO) and KEGG Analysis
3. Results
3.1. Sample Information and Sequencing Results
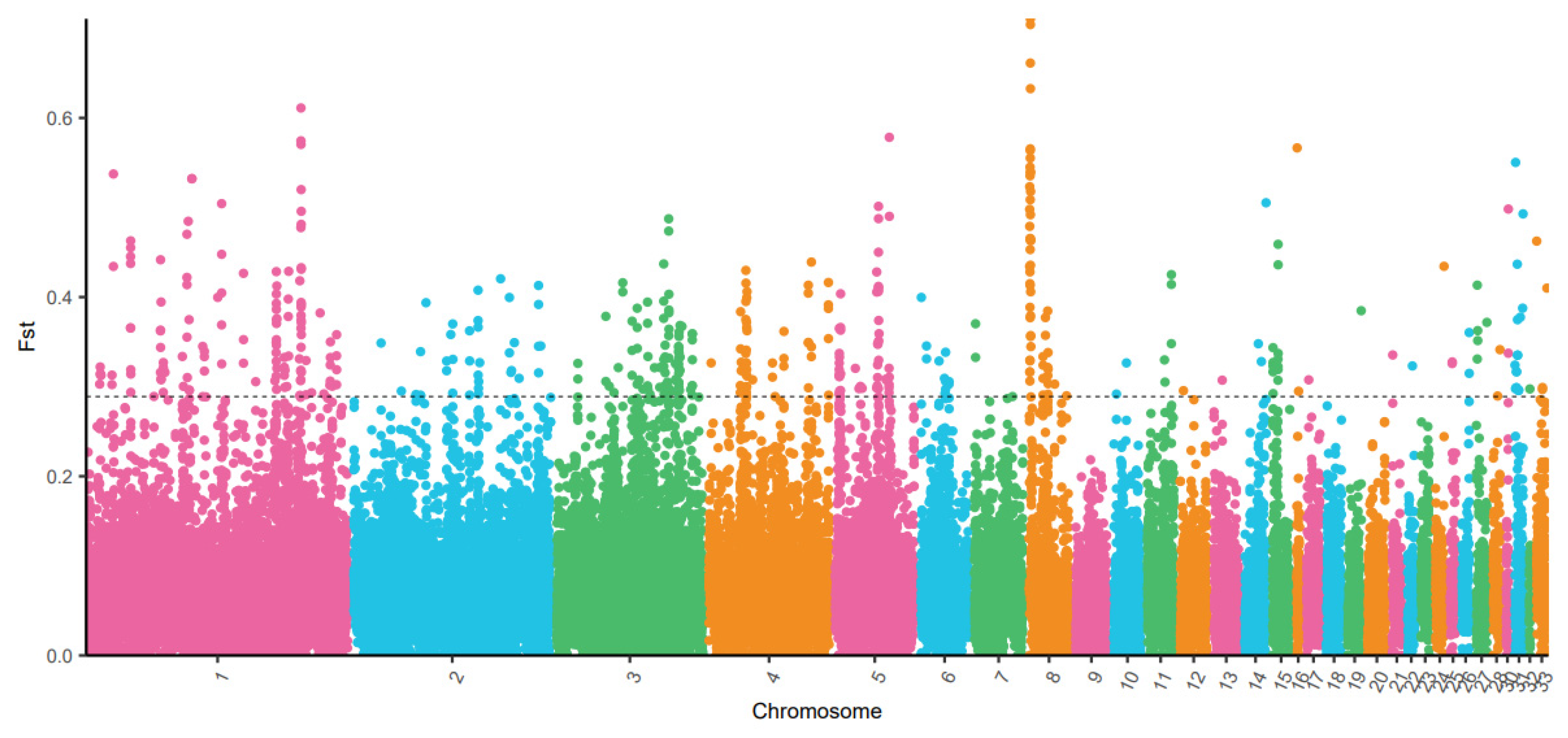
3.2. Population Differentiation Based on Fst
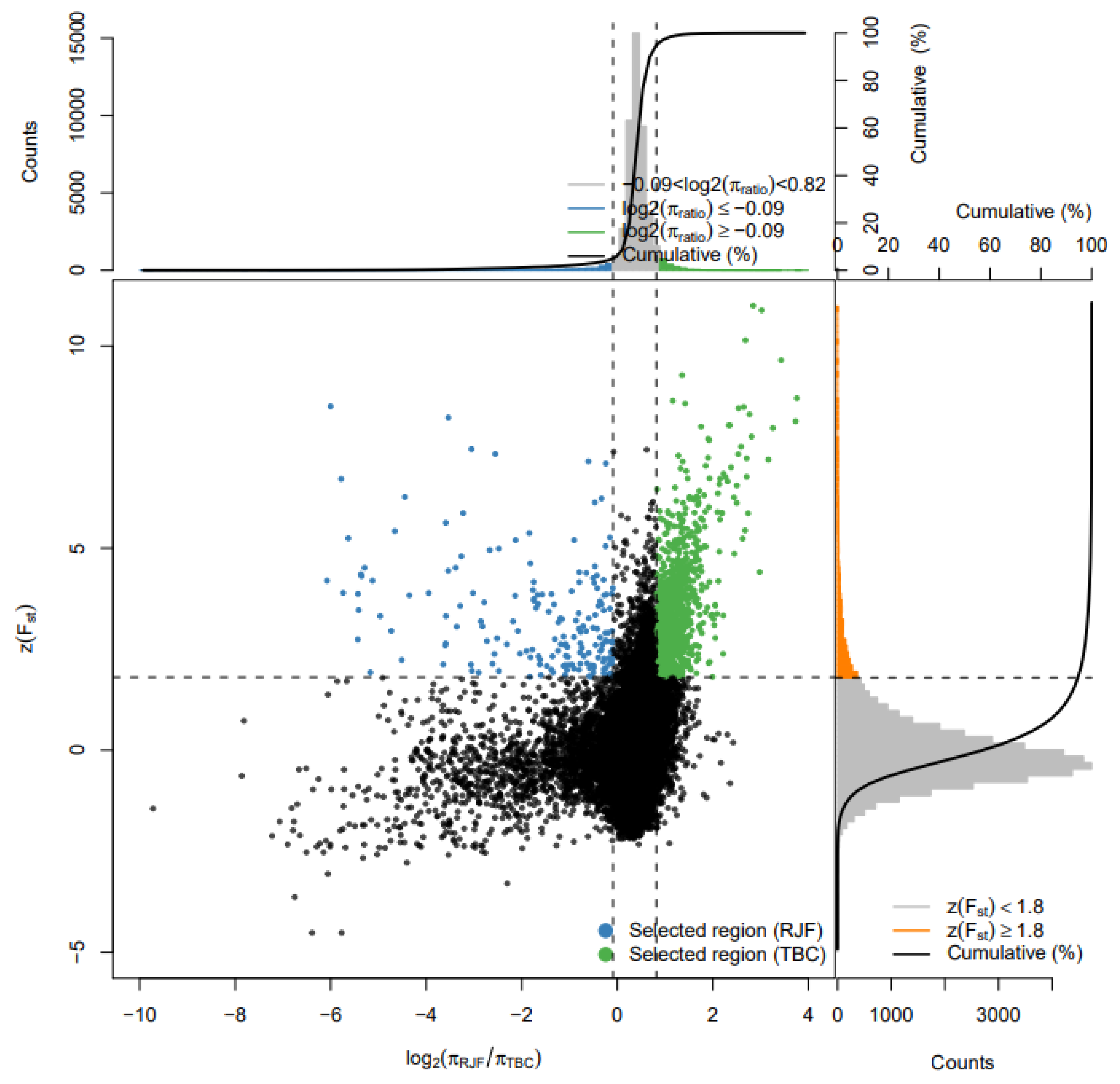
3.3. Sweeping Selection Based on Fst and Pi
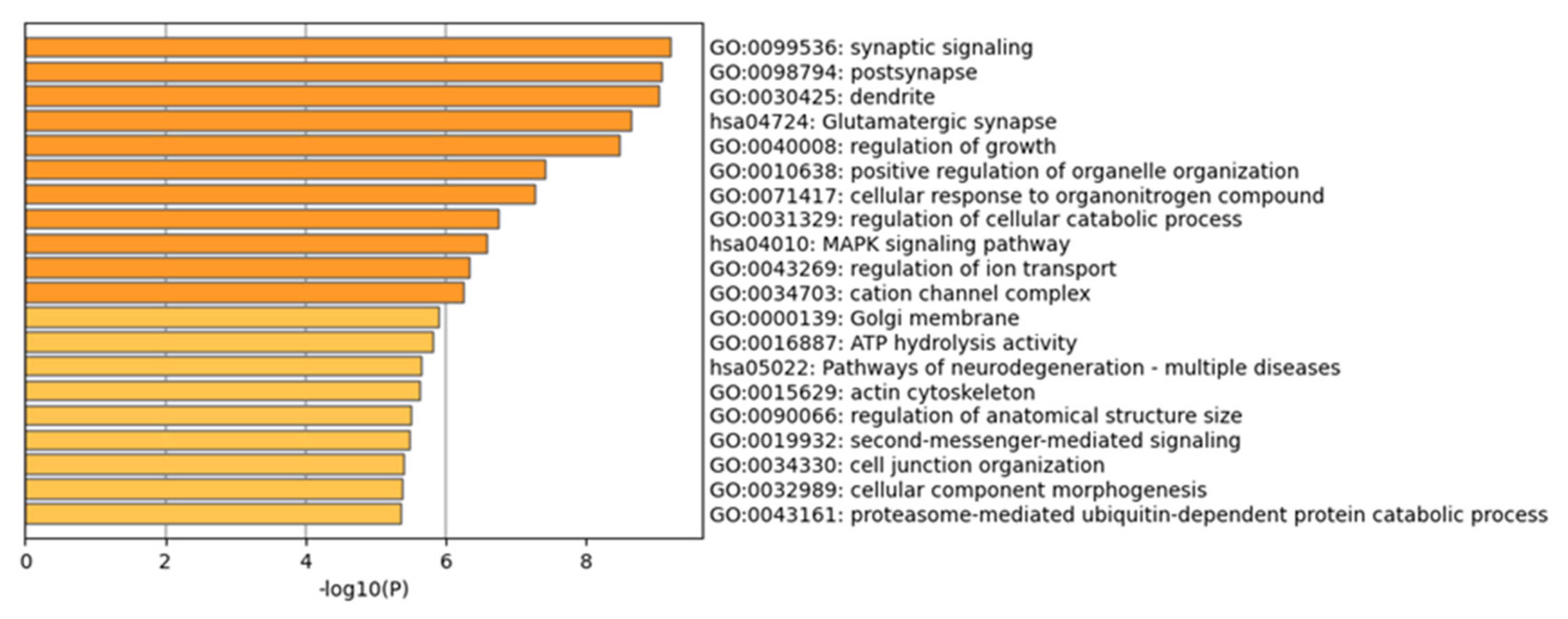
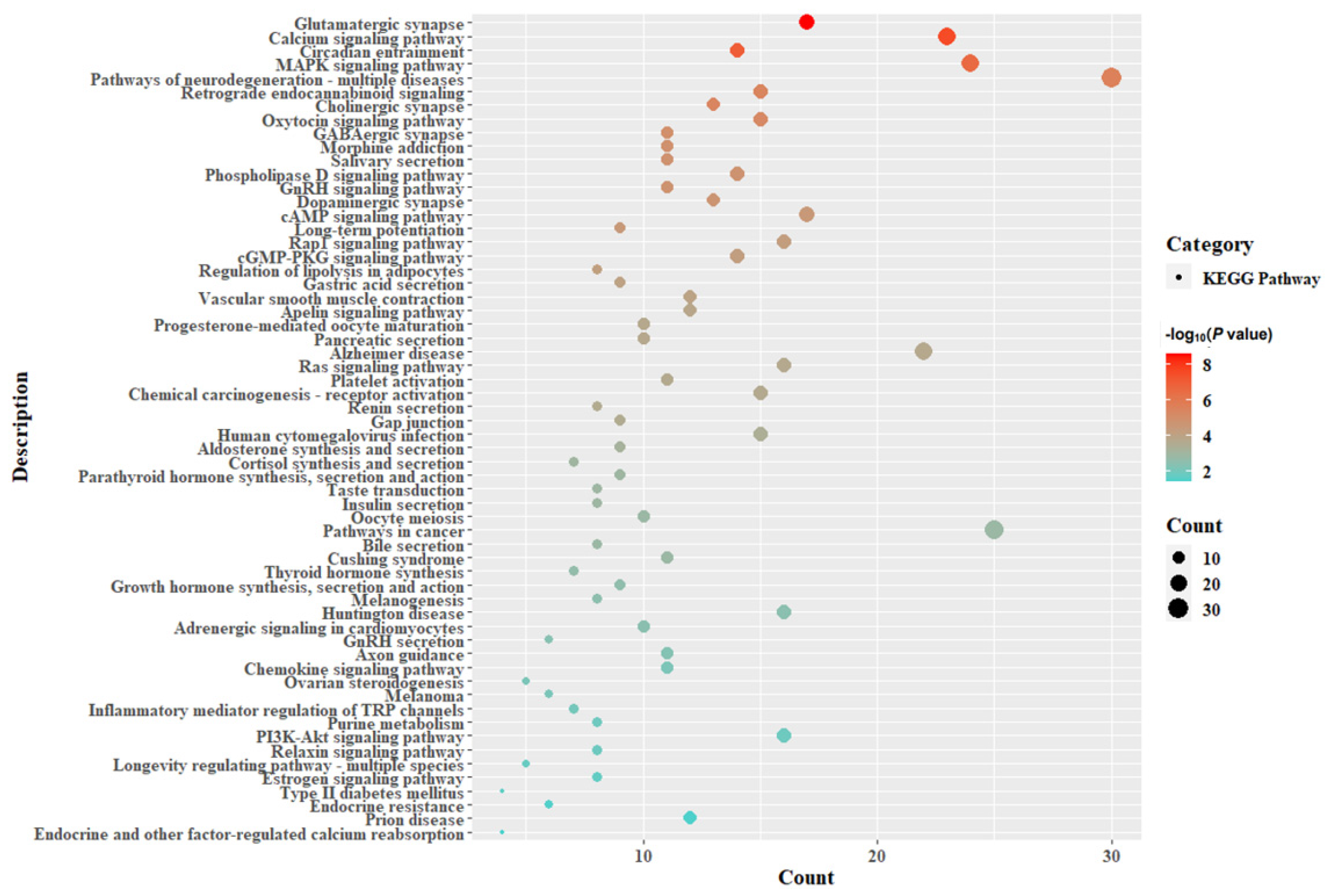
3.4. GO and KEGG Enrichment Analysis
4. Discussion
4.1. Functional Annotation of TBCs
4.2. Potential Disease Resistance in TBCs
4.3. High-Altitude Adaptation in TBCs
4.4. Egg Production
4.5. Aggressiveness
4.6. Potentially Selected Genes Related to Vision, Hearing and Circadian Rhythms
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, M.S.; Li, Y.; Peng, M.S.; Zhong, L.; Wang, Z.J.; Li, Q.Y.; Tu, X.L.; Dong, Y.; Zhu, C.L.; Wang, L.; et al. Genomic Analyses Reveal Potential Independent Adaptation to High Altitude in Tibetan Chickens. Mol. Biol. Evol. 2015, 32, 1880–1889. [Google Scholar] [CrossRef] [PubMed]
- Storz, J.F.; Scott, G.R.; Cheviron, Z.A. Phenotypic plasticity and genetic adaptation to high-altitude hypoxia in vertebrates. J. Exp. Biol. 2010, 213, 4125–4136. [Google Scholar] [CrossRef] [PubMed]
- Shang, P.; Dong, Y.N.; Zhu, X.H. Research status and suggestions on Tibetan chicken breeding. Chin. Poult. 2019, 41, 45–49. [Google Scholar]
- Chen, B.; Li, D.; Ran, B.; Zhang, P.; Wang, T. Key miRNAs and Genes in the High-Altitude Adaptation of Tibetan Chickens. Front. Vet. Sci. 2022, 9, 911685. [Google Scholar] [CrossRef]
- Guo, X.Y.; Zhang, J.; Li, M.R. Analysis of microsatellite characteristics of Tibetan chicken genome. Chin. Poult. 2020, 42, 122–126. [Google Scholar]
- Jia, F.C.; Qin, L.Y.; Li, D.D. Based on high-throughput sequencing to analyze the distribution characteristics of the intestinal bacterial community of Hegu Tibetan chicken and Bailaihang chicken under the same rearing mode. Plateau Agric. 2018, 2, 576–584. [Google Scholar]
- Stephan, W. Genetic hitchhiking versus background selection: The controversy and its implications. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 1245–1253. [Google Scholar] [CrossRef]
- Fujito, N.T.; Satta, Y.; Hayakawa, T.; Takahata, N. A new inference method for detecting an ongoing selective sweep. Genes Genet. Syst. 2018, 93, 149–161. [Google Scholar] [CrossRef]
- Quilez, J.; Short, A.D.; Martinez, V.; Kennedy, L.J.; Ollier, W.; Sanchez, A.; Altet, L.; Francino, O. A selective sweep of >8 Mb on chromosome 26 in the Boxer genome. BMC Genom. 2011, 12, 339. [Google Scholar] [CrossRef]
- Stephan, W. Selective Sweeps. Genetics 2019, 211, 5–13. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhang, Q.; Zeng, S.; Gu, L.; Si, W.; Zhang, X.; Tian, D.; Yang, S.; Wang, L. Selective sweep with significant positive selection serves as the driving force for the differentiation of japonica and indica rice cultivars. BMC Genom. 2017, 18, 307. [Google Scholar] [CrossRef] [PubMed]
- Alachiotis, N.; Pavlidis, P. RAiSD detects positive selection based on multiple signatures of a selective sweep and SNP vectors. Commun. Biol. 2018, 1, 79. [Google Scholar] [CrossRef] [PubMed]
- Kozma, R.; Rodin-Morch, P.; Hoglund, J. Genomic regions of speciation and adaptation among three species of grouse. Sci. Rep. 2019, 9, 812. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, Y.; Li, M.; Che, T.; Tian, S.; Chen, B.; Zhou, X.; Zhang, G.; Gaur, U.; Luo, M.; et al. Population genomics identifies patterns of genetic diversity and selection in chicken. BMC Genom. 2019, 20, 263. [Google Scholar] [CrossRef]
- Zhang, Q.; Gou, W.; Wang, X.; Zhang, Y.; Ma, J.; Zhang, H.; Zhang, Y.; Zhang, H. Genome Resequencing Identifies Unique Adaptations of Tibetan Chickens to Hypoxia and High-Dose Ultraviolet Radiation in High-Altitude Environments. Genome Biol. Evol. 2016, 8, 765–776. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.S.; Thakur, M.; Peng, M.S.; Jiang, Y.; Frantz, L.A.F.; Li, M.; Zhang, J.J.; Wang, S.; Peters, J.; Otecko, N.O.; et al. 863 genomes reveal the origin and domestication of chicken. Cell Res. 2020, 30, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.G.; Zhao, C.J.; Li, J.Y.; Wu, C.X. Sequencing and alignment of mitochondrial genomes of Tibetan chicken and two lowland chicken breeds. Chin. Sci. Life Sci. Engl. Ed. 2008, 51, 47–51. [Google Scholar] [CrossRef]
- Tian, S.; Zhou, X.; Zhang, D.; Ning, C.; Li, D.; Zhao, H. Genomic Analyses Reveal Genetic Adaptations to Tropical Climates in Chickens; Social Science Electronic Publishing: Rochester, NY, USA, 2020. [Google Scholar]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Mckenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; Depristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Kai, W.; Li, M.; Hakon, H. ANNOVAR: Functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010, 38, e164. [Google Scholar]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Hanif, Q.; Cao, Y.; Yu, Y.; Zhao, Y. Whole Genome Scan and Selection Signatures for Climate Adaption in Yanbian Cattle. Front. Genet. 2020, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- Rubin, C.J.; Zody, M.C.; Eriksson, J.; Meadows, J.R.; Sherwood, E.; Webster, M.T.; Jiang, L.; Ingman, M.; Sharpe, T.; Ka, S.; et al. Whole-genome resequencing reveals loci under selection during chicken domestication. Nature 2010, 464, 587–591. [Google Scholar] [CrossRef]
- Liisa, L.; Thomas, M.G.; Ross, B.; Richard, A.; Naomi, S.; Paxinos, P.D.; Ophélie, L.; Keith, D.; Joris, P.; Andrea, M. Inferring Allele Frequency Trajectories from Ancient DNA Indicates That Selection on a Chicken Gene Coincided with Changes in Medieval Husbandry Practices. Mol. Biol. Evol. 2017, 34, 1981–1990. [Google Scholar]
- Lawal, R.A.; Al-Atiyat, R.M.; Aljumaah, R.S.; Pradeepa, S.; Mwacharo, J.M.; Olivier, H. Whole-Genome Resequencing of Red Junglefowl and Indigenous Village Chicken Reveal New Insights on the Genome Dynamics of the Species. Front. Genet. 2018, 9, 264. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Huh, J.; Kim, W.; Jeong, S.; do Min, S.; Jung, Y. Phospholipase D activates HIF-1-VEGF pathway via phosphatidic acid. Exp. Mol. Med. 2014, 46, e126. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.W.; Choi, K.Y.; do Min, S. Phospholipase D meets Wnt signaling: A new target for cancer therapy. Cancer Res. 2011, 71, 293–297. [Google Scholar] [CrossRef]
- Gandhi, R.; Bonaccorsi, S.; Wentworth, D.; Doxsey, S.; Gatti, M.; Pereira, A. The Drosophila Kinesin-like Protein KLP67A Is Essential for Mitotic and Male Meiotic Spindle Assembly. Mol. Biol. Cell 2004, 15, 121–131. [Google Scholar] [CrossRef]
- Liu, X.S.; Zhao, X.D.; Wang, X.; Yao, Y.X.; Wang, Z.G. Germinal Cell Aplasia in Kif18a Mutant Male Mice Due to Impaired Chromosome Congression and Dysregulated BubR1 and CENP-E. Genes. Cancer 2010, 1, 26–39. [Google Scholar] [CrossRef]
- Li, M.; Sun, C.; Xu, N.; Bian, P.; Tian, X.; Wang, X.; Wang, Y.; Jia, X.; Rasmus, H.; Wang, M. De Novo Assembly of 20 Chicken Genomes Reveals the Undetectable Phenomenon for Thousands of Core Genes on Microchromosomes and Subtelomeric Regions. Mol. Biol. Evol. 2022, 39, msac066. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.R.; Carniel, E.; Mestroni, L. Cardiomyopathy, familial dilated. Orphanet J. Rare Dis. 2006, 1, 27. [Google Scholar] [CrossRef] [PubMed]
- Stoffel, W.; Jenke, B.; Block, B.; Zumbansen, M.; Koebke, J. Neutral sphingomyelinase 2 (smpd3) in the control of postnatal growth and development. Proc. Natl. Acad. Sci. USA 2005, 102, 4554–4559. [Google Scholar] [CrossRef]
- James, A.W.; Shen, J.; Zhang, X.; Asatrian, G.; Goyal, R.; Kwak, J.H.; Jiang, L.; Bengs, B.; Culiat, C.T.; Turner, A.S.; et al. NELL-1 in the treatment of osteoporotic bone loss. Nat. Commun. 2015, 6, 7362. [Google Scholar] [CrossRef] [PubMed]
- Mure, H.; Matsuzaki, K.; Kitazato, K.T.; Mizobuchi, Y.; Kuwayama, K.; Kageji, T.; Nagahiro, S. Akt2 and Akt3 play a pivotal role in malignant gliomas. Neuro Oncol. 2010, 12, 221–232. [Google Scholar] [CrossRef]
- Jun, I.; Choi, Y.J.; Kim, B.R.; Seo, K.Y.; Kim, T.I. Activation of ADRB2/PKA Signaling Pathway Facilitates Lipid Synthesis in Meibocytes, and β-Blocker Glaucoma Drug Impedes PKA-Induced Lipid Synthesis by Inhibiting ADRB2. Int. J. Mol. Sci. 2022, 23, 9478. [Google Scholar] [CrossRef]
- Balakrishnan, B.; Gupta, A.; Basri, R.; Sharma, V.M.; Slayton, M.; Gentner, K.; Becker, C.C.; Karki, S.; Muturi, H.; Najjar, S.M.; et al. Endothelial-Specific Expression of CIDEC Improves High-Fat Diet-Induced Vascular and Metabolic Dysfunction. Diabetes 2023, 72, 19–32. [Google Scholar] [CrossRef]
- Xue, J.Y.; Grigelioniene, G.; Wang, Z.; Nishimura, G.; Iida, A.; Matsumoto, N.; Tham, E.; Miyake, N.; Ikegawa, S.; Guo, L. SLC4A2 Deficiency Causes a New Type of Osteopetrosis. J. Bone Min. Res. 2022, 37, 226–235. [Google Scholar] [CrossRef]
- Qureshi, O.S.; Zheng, Y.; Nakamura, K.; Attridge, K.; Manzotti, C.; Schmidt, E.M.; Baker, J.; Jeffery, L.E.; Kaur, S.; Briggs, Z.; et al. Trans-endocytosis of CD80 and CD86: A molecular basis for the cell-extrinsic function of CTLA-4. Science 2011, 332, 600–603. [Google Scholar] [CrossRef]
- Most, P.; Jong, B.D.; Parmentier, H.K.; Verhulst, S. Trade-off between growth and immune function: A meta-analysis of selection experiment. Funct. Ecol. 2010, 25, 74. [Google Scholar] [CrossRef]
- Shwan, N.A.A.; Louzada, S.; Yang, F.; Armour, J.A.L. Recurrent Rearrangements of Human Amylase Genes Create Multiple Independent CNV Series. Hum. Mutat. 2017, 38, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Yao, Y.; Yin, H.; Cai, Z.; Wang, Y.; Bai, L.; Kern, C.; Halstead, M.; Chanthavixay, G.; Trakooljul, N.; et al. Pig genome functional annotation enhances the biological interpretation of complex traits and human disease. Nat. Commun. 2021, 12, 5848. [Google Scholar] [CrossRef] [PubMed]
- Feng, P.; Zeng, T.; Hua, Y.; Chen, G.; Lu, L. Whole-genome Resequencing Provides Insights into the Population Structure and Domestication Signatures of Ducks in Eastern China. BMC Genom. 2020, 22, 401. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Sun, C.; Qu, L.; Wang, K.; Yang, N. Genome-Wide Detection of Selective Signatures in Chicken through High Density SNPs. PLoS ONE 2016, 11, e0166146. [Google Scholar]
- Smaldino, P.J.; Read, D.F.; Pratt-Hyatt, M.; Hopper, A.K.; Engelke, D.R. The cytoplasmic and nuclear populations of the eukaryote tRNA-isopentenyl transferase have distinct functions with implications in human cancer. Gene 2015, 556, 13–18. [Google Scholar]
- Hunsucker, S.A.; Spychala, J.; Mitchell, B.S. Human cytosolic 5′-nucleotidase I: Characterization and role in nucleoside analog resistance. J. Biol. Chem. 2001, 276, 10498–10504. [Google Scholar] [CrossRef]
- Burtscher, M.; Millet, G.P.; Burtscher, J. Hypoxia Conditioning for High-Altitude Pre-acclimatization. J. Sci. Sport. Exerc. 2022, 4, 331–345. [Google Scholar] [CrossRef]
- Kathiriya, I.S.; King, I.N.; Murakami, M.; Nakagawa, M.; Astle, J.M.; Gardner, K.A.; Gerard, R.D.; Olson, E.N.; Srivastava, D.; Nakagawa, O. Hairy-related Transcription Factors Inhibit GATA-dependent Cardiac Gene Expression through a Signal-responsive Mechanism. J. Biol. Chem. 2004, 279, 54937–54943. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, C.X.; Chamba, Y.; Ling, Y. Blood Characteristics for High Altitude Adaptation in Tibetan Chickens 1. Poult. Sci. 2007, 86, 1384–1389. [Google Scholar] [CrossRef]
- Kviklyte, S.; Vertommen, D.; Yerna, X.; Andersén, H.; Xu, X.; Gailly, P.; Bohlooly, Y.M.; Oscarsson, J.; Rider, M.H. Effects of genetic deletion of soluble 5’-nucleotidases NT5C1A and NT5C2 on AMPK activation and nucleotide levels in contracting mouse skeletal muscles. Am. J. Physiol. Endocrinol. Metab. 2017, 313, E48–E62. [Google Scholar] [CrossRef]
- Stoffel, W.; Hammels, I.; Jenke, B.; Binczek, E.; Schmidt-Soltau, I.; Brodesser, S.; Schauss, A.; Etich, J.; Heilig, J.; Zaucke, F. Neutral sphingomyelinase (SMPD3) deficiency disrupts the Golgi secretory pathway and causes growth inhibition. Cell Death Dis. 2016, 7, e2488. [Google Scholar] [CrossRef] [PubMed]
- Stoffel, W.; Hammels, I.; Jenke, B.; Schmidt-Soltau, I.; Niehoff, A. Neutral Sphingomyelinase 2 (SMPD3)-Deficiency in Mice Causes Chondrodysplasia with Unimpaired Skeletal Mineralization. Am. J. Pathol. 2019, 189, 1831–1845. [Google Scholar] [CrossRef] [PubMed]
- Dunham-Snary, K.J.; Wu, D.; Potus, F.; Sykes, E.A.; Mewburn, J.D.; Charles, R.L.; Eaton, P.; Sultanian, R.A.; Archer, S.L. Ndufs2, a Core Subunit of Mitochondrial Complex I, Is Essential for Acute Oxygen-Sensing and Hypoxic Pulmonary Vasoconstriction. Circ. Res. 2019, 124, 1727–1746. [Google Scholar] [CrossRef] [PubMed]
- Czechanski, A.; Kim, H.; Byers, C.; Greenstein, I.; Stumpff, J.; Reinholdt, L.G. Kif18a is specifically required for mitotic progression during germ line development. Dev. Biol. 2015, 402, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Sun, C.; Dou, T.; Yi, G.; Qu, L.; Qu, L.; Wang, K.; Yang, N. Identification of Promising Mutants Associated with Egg Production Traits Revealed by Genome-Wide Association Study. PLoS ONE 2015, 10, e0140615. [Google Scholar] [CrossRef]
- Tiunova, A.A.; Komissarova, N.V.; Anokhin, K.V. Prenatal Sensory Stimulation Induces BDNF Gene Expression in the Brain and Potentiates the Development of Species-Specific Predisposition in Newborn Chicks. Bull. Exp. Biol. Med. 2018, 166, 229–232. [Google Scholar] [CrossRef]
- Tolosa, A.; Zhou, X.; Spittau, B.; Krieglstein, K. Establishment of a Survival and Toxic Cellular Model for Parkinson’s Disease from Chicken Mesencephalon. Neurotox. Res. 2013, 24, 119–129. [Google Scholar] [CrossRef]
- Guo, X.; Fang, Q.; Ma, C.; Zhou, B.; Wan, Y.; Jiang, R. Whole-genome resequencing of Xishuangbanna fighting chicken to identify signatures of selection. Genet. Sel. Evol. 2016, 48, 62. [Google Scholar] [CrossRef]
- Yokoyama, S. Molecular evolution of color vision in vertebrates. Gene 2002, 300, 69–78. [Google Scholar] [CrossRef]
- Wang, M.S.; Zhang, R.W.; Su, L.Y.; Li, Y.; Peng, M.S.; Liu, H.Q.; Zeng, L.; Irwin, D.M.; Du, J.L.; Yao, Y.G.; et al. Positive selection rather than relaxation of functional constraint drives the evolution of vision during chicken domestication. Cell Res. 2016, 26, 556–573. [Google Scholar] [CrossRef]
- Crouse, J.A.; Lopes, V.S.; Sanagustin, J.T.; Keady, B.T.; Williams, D.S.; Pazour, G.J. Distinct functions for IFT140 and IFT20 in opsin transport. Cytoskeleton 2014, 71, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Krock, B.L.; Perkins, B.D. The intraflagellar transport protein IFT57 is required for cilia maintenance and regulates IFT-particle–kinesin-II dissociation in vertebrate photoreceptors. J. Cell Sci. 2008, 121, 1907–1915. [Google Scholar] [CrossRef] [PubMed]
- Okada, I.; Hamanoue, H.; Terada, K.; Tohma, T.; Megarbane, A.; Chouery, E.; Abou-Ghoch, J.; Jalkh, N.; Cogulu, O.; Ozkinay, F.; et al. SMOC1 is essential for ocular and limb development in humans and mice. Am. J. Hum. Genet. 2011, 88, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chai, R. Hear the sounds: The role of G protein-coupled receptors in the cochlea. Am. J. Physiol. Cell Physiol. 2022, 323, C1088–C1099. [Google Scholar] [CrossRef] [PubMed]
- Santos-Cortez, R.L.; Lee, K.; Giese, A.P.; Ansar, M.; Amin-Ud-Din, M.; Rehn, K.; Wang, X.; Aziz, A.; Chiu, I.; Hussain Ali, R.; et al. Adenylate cyclase 1 (ADCY1) mutations cause recessive hearing impairment in humans and defects in hair cell function and hearing in zebrafish. Hum. Mol. Genet. 2014, 23, 3289–3298. [Google Scholar] [CrossRef]
- Lai, W.; Yu, L. Elevated MicroRNA 183 Impairs Trophoblast Migration and Invasiveness by Downregulating FOXP1 Expression and Elevating GNG7 Expression during Preeclampsia. Mol. Cell. Biol. 2020, 41, e00236-20. [Google Scholar] [CrossRef]
- Van der Veen, D.R.; Laing, E.E.; Bae, S.E.; Johnston, J.D.; Dijk, D.J.; Archer, S.N. A Topological Cluster of Differentially Regulated Genes in Mice Lacking PER3. Front. Mol. Neurosci. 2020, 13, 15. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Clean Reads | Coverage Depth (X) | Raw Base (G) | Clean Base (G) | Effective Rate (%) | Q20 (%) | Q30(%) | GC Content (%) |
|---|---|---|---|---|---|---|---|---|
| 1 | 70346492 | 8.59 | 10.57 | 10.55 | 99.83 | 95.95 | 89.99 | 43.21 |
| 2 | 67577374 | 7.33 | 10.16 | 10.14 | 99.74 | 97.47 | 93.6 | 42.88 |
| 3 | 77612348 | 9.67 | 11.66 | 11.64 | 99.84 | 96.13 | 89.64 | 42.36 |
| 4 | 65717850 | 6.81 | 9.88 | 9.86 | 99.76 | 97.53 | 93.59 | 42.97 |
| 5 | 69322110 | 7.47 | 10.42 | 10.40 | 99.76 | 97.32 | 93.16 | 42.37 |
| 6 | 67171880 | 7.1 | 10.10 | 10.08 | 99.7 | 97.08 | 92.73 | 43.00 |
| 7 | 73091286 | 7.36 | 10.99 | 10.96 | 99.72 | 97.33 | 93.22 | 42.52 |
| 8 | 70688764 | 7.57 | 10.64 | 10.60 | 99.67 | 97.22 | 93.09 | 42.09 |
| 9 | 66844024 | 7.32 | 10.06 | 10.03 | 99.7 | 96.69 | 91.73 | 42.27 |
| 10 | 94968550 | 10.64 | 14.27 | 14.25 | 99.84 | 96.68 | 91.56 | 42.56 |
| 11 | 69673108 | 7.55 | 10.48 | 10.45 | 99.74 | 97.14 | 93.16 | 42.95 |
| 12 | 75912388 | 9.31 | 11.41 | 11.39 | 99.84 | 95.82 | 89.75 | 42.7 |
| 13 | 77491138 | 9.36 | 11.64 | 11.62 | 99.84 | 95.78 | 89.57 | 42.77 |
| 14 | 71980470 | 8.86 | 10.81 | 10.80 | 99.84 | 96.34 | 90.7 | 42.55 |
| 15 | 76923340 | 8.22 | 11.58 | 11.54 | 99.67 | 97.00 | 92.81 | 43.37 |
| 16 | 71735168 | 7.71 | 10.79 | 10.76 | 99.74 | 97.11 | 92.79 | 43.77 |
| 17 | 97405862 | 11.71 | 14.64 | 14.61 | 99.83 | 96.02 | 90.19 | 42.49 |
| 18 | 70090036 | 7.47 | 10.55 | 10.51 | 99.59 | 96.48 | 91.77 | 43.13 |
| 19 | 72877790 | 7.7 | 10.97 | 10.93 | 99.65 | 96.67 | 91.97 | 42.9 |
| 20 | 70938848 | 8.06 | 10.67 | 10.64 | 99.69 | 96.94 | 92.42 | 42.81 |
| 21 | 70079014 | 7.53 | 10.55 | 10.51 | 99.6 | 96.64 | 91.98 | 42.7 |
| 22 | 64508976 | 6.96 | 9.70 | 9.68 | 99.7 | 97.25 | 93.19 | 43.27 |
| 23 | 68653894 | 7.35 | 10.32 | 10.30 | 99.73 | 97.44 | 93.53 | 43.22 |
| 24 | 69469802 | 7.54 | 10.45 | 10.42 | 99.69 | 97.06 | 92.9 | 43.64 |
| 25 | 68040032 | 7.23 | 10.23 | 10.21 | 99.71 | 96.8 | 92.13 | 42.65 |
| 26 | 67368530 | 7.24 | 10.13 | 10.11 | 99.73 | 96.78 | 92.06 | 42.53 |
| 27 | 84735694 | 10.4 | 12.73 | 12.71 | 99.84 | 95.58 | 89.35 | 42.33 |
| 28 | 63404212 | 3.04 | 9.56 | 9.51 | 99.51 | 95.61 | 90.52 | 48.93 |
| 29 | 71416314 | 7.56 | 10.74 | 10.71 | 99.73 | 97.22 | 92.92 | 42.52 |
| 30 | 71800010 | 7.85 | 10.79 | 10.77 | 99.74 | 96.81 | 91.97 | 42.65 |
| Total SNP | 9,490,690 | 100% | |
| Upstream | 112,547 | 1.19% | |
| Exonic | Nonsynonymous | 91,261 | 2.09% |
| Stopgain | 424 | ||
| Stoploss | 68 | ||
| Synonymous | 106,556 | ||
| Intronic | 4,427,978 | 46.66% | |
| Upstream/Downstream | 7921 | 0.08% | |
| Downstream | 116,946 | 1.23% | |
| Intergenic | 4,565,787 | 48.11% | |
| Transitions | 6,814,921 | ||
| Transversions | 2,675,769 | ||
| Transitions/Transversions | 2.55 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, J.; Zhu, W.; Shi, H.; Peng, D.; Zang, L.; Wang, Y.; ZhaXi, L.; BaiMa, J.; Amevor, F.K.; Wang, X.; et al. Analysis of the Selection Signal of the Tibetan Black Chicken Genome Based on Whole-Genome Sequencing. Genes 2023, 14, 1672. https://doi.org/10.3390/genes14091672
Feng J, Zhu W, Shi H, Peng D, Zang L, Wang Y, ZhaXi L, BaiMa J, Amevor FK, Wang X, et al. Analysis of the Selection Signal of the Tibetan Black Chicken Genome Based on Whole-Genome Sequencing. Genes. 2023; 14(9):1672. https://doi.org/10.3390/genes14091672
Chicago/Turabian StyleFeng, Jing, Wei Zhu, Hairen Shi, Da Peng, Lei Zang, Yan Wang, Luobu ZhaXi, Jiancai BaiMa, Felix Kwame Amevor, Xiaoqi Wang, and et al. 2023. "Analysis of the Selection Signal of the Tibetan Black Chicken Genome Based on Whole-Genome Sequencing" Genes 14, no. 9: 1672. https://doi.org/10.3390/genes14091672