
Identification of the Light-Harvesting Chlorophyll a/b Binding Protein Gene Family in Peach (Prunus persica L.) and Their Expression under Drought Stress
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genome-Wide Identification of PpLhc Genes
2.2. Analysis of Chromosome Distribution and Physicochemical Properties of PpLhc Proteins
2.3. Phylogenetic Relationships Analysis
2.4. Gene Structure and Protein Motif Analysis
2.5. Prediction of Subcellular Localization and Transmembrane Domain
2.6. Collinearity and Promoter Element Analysis
2.7. RNA-Seq Data Analysis
2.8. Plant Materials and Treatments
2.9. RNA Extraction and RT-qPCR
2.10. Data Analysis
3. Results
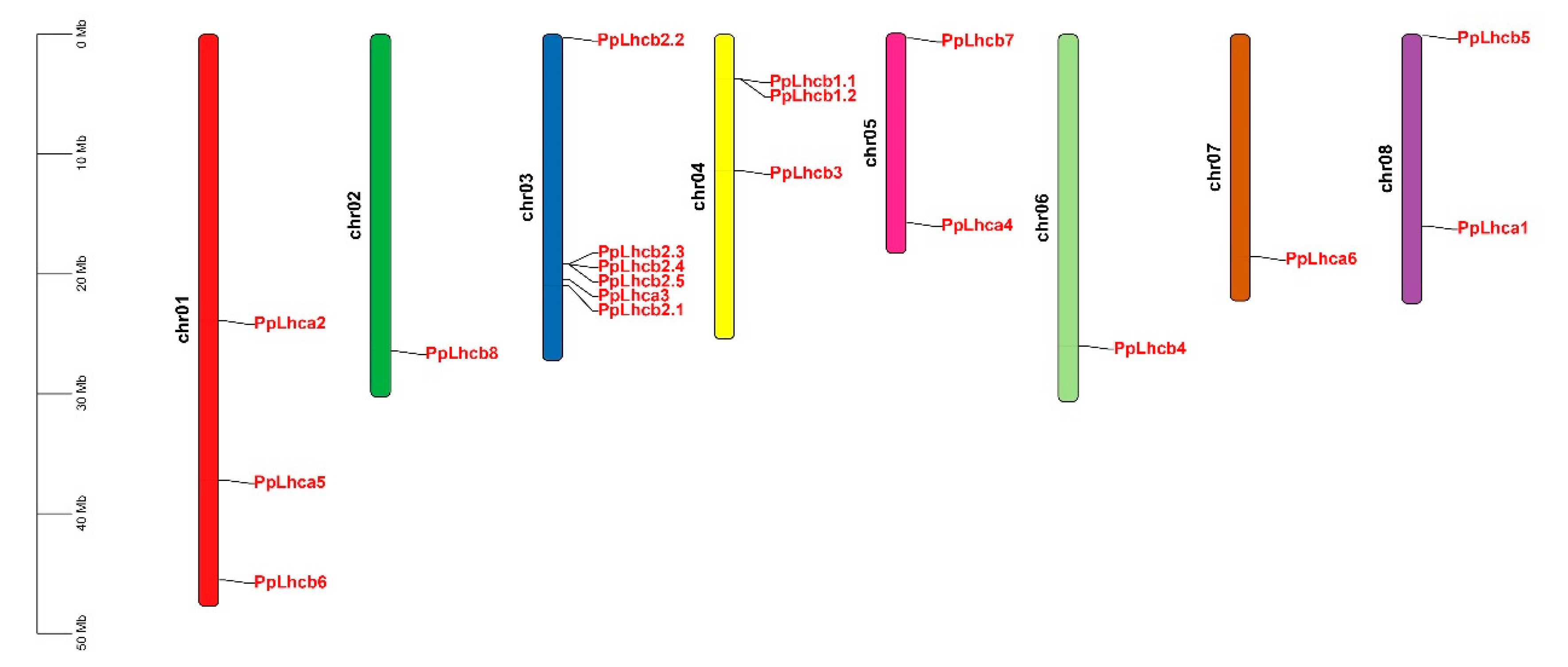
3.1. Genome-Wide Identification and Characteristics of PpLhc Genes
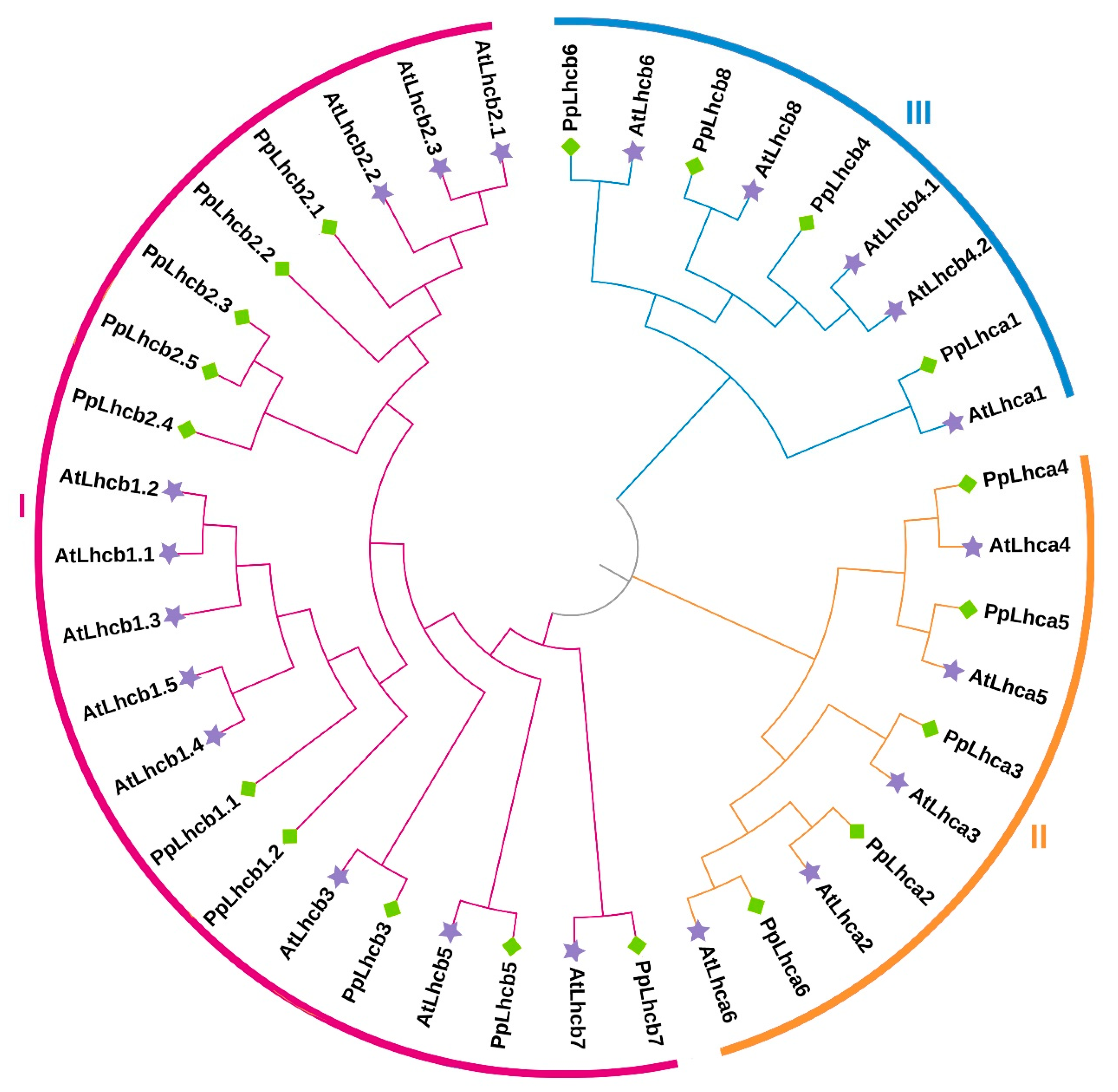
3.2. Phylogenetic Relationship of Peach Lhc Proteins
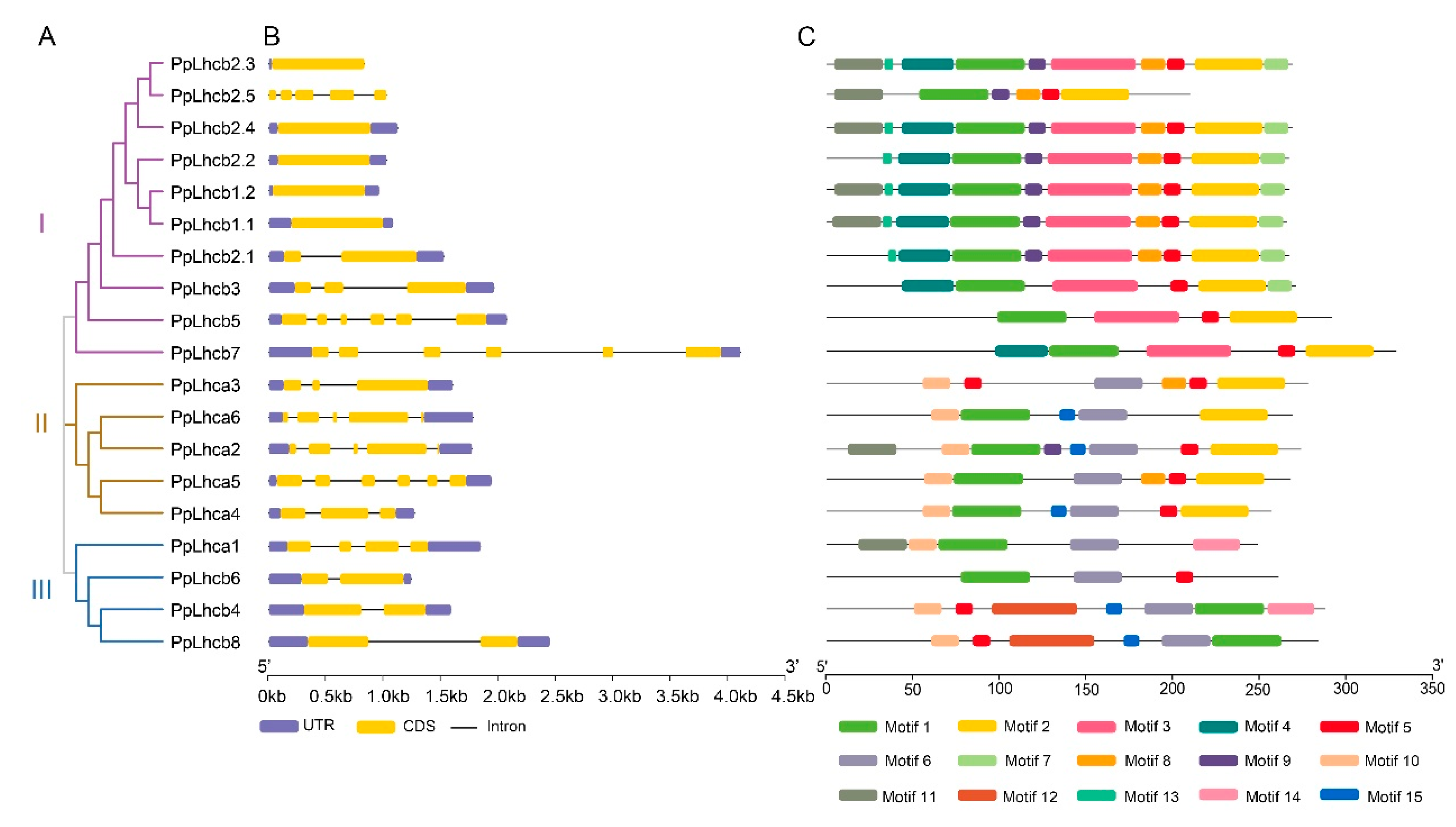
3.3. Gene Structure and Protein Motif Analyses of PpLhc Members
3.4. Subcellular Localization and Transmembrane Prediction of PpLhc Proteins
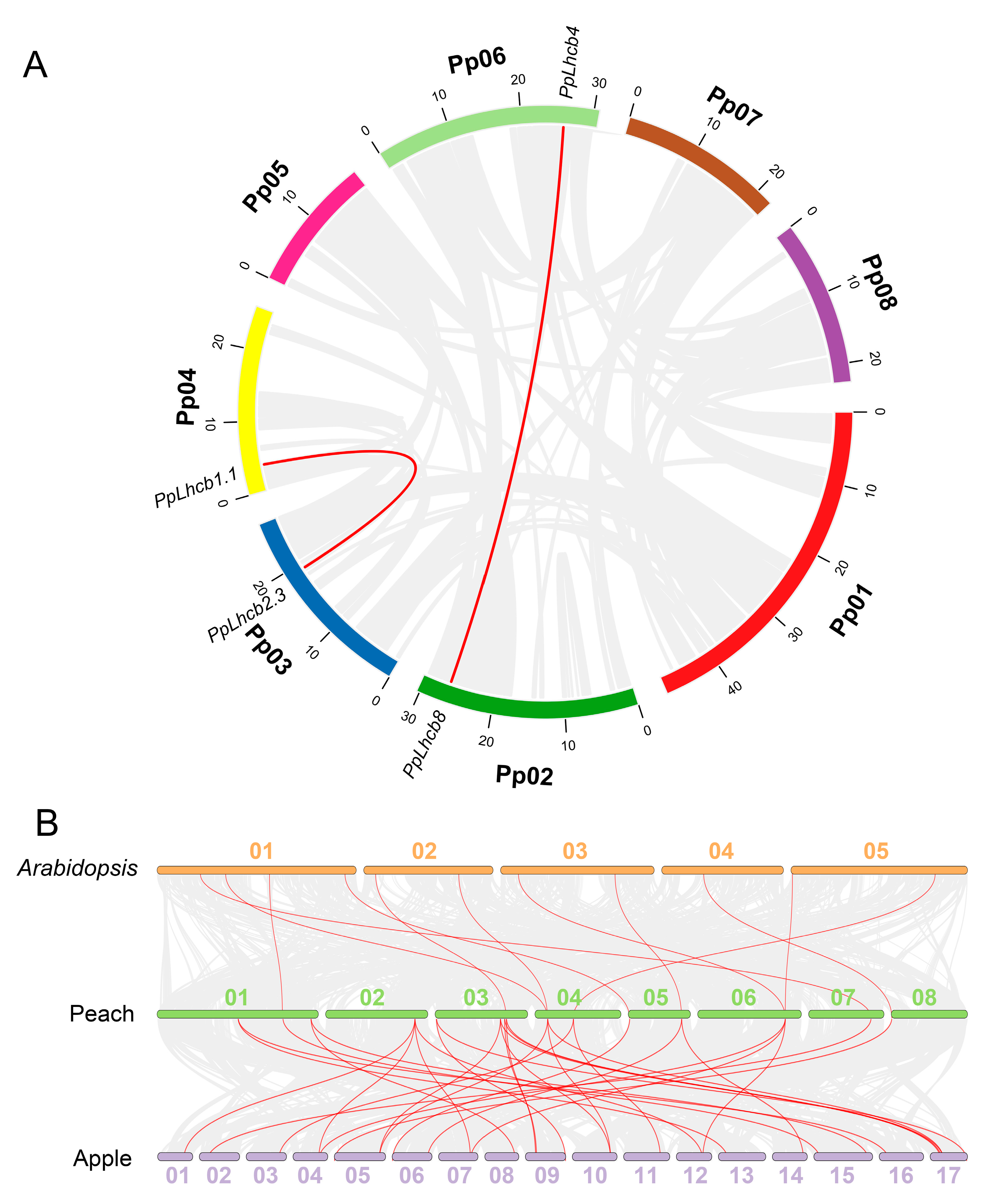
3.5. Collinearity Analysis of PpLhc Genes
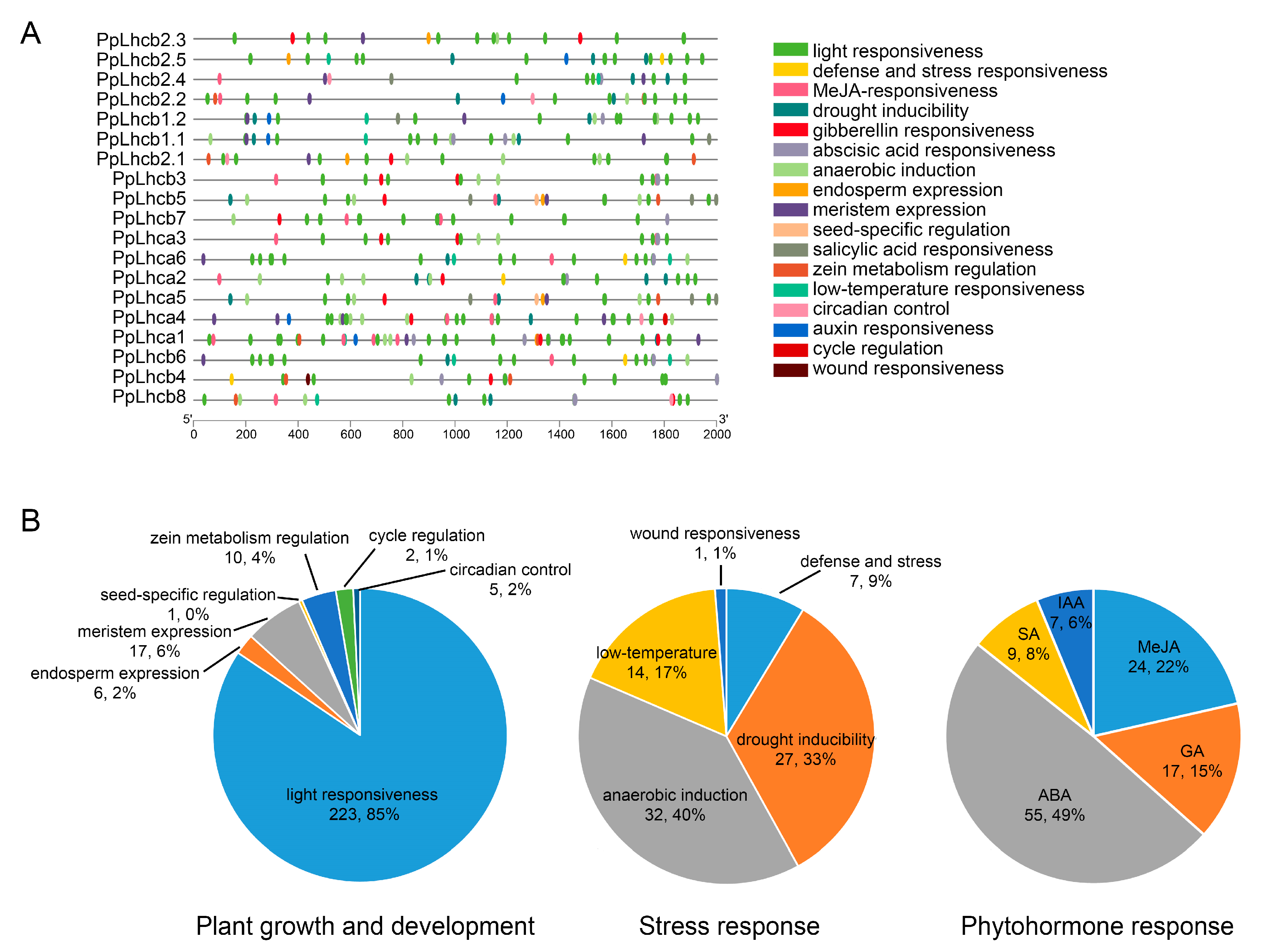
3.6. Identification of Cis-Regulatory Elements in the Promoter of PpLhc Genes
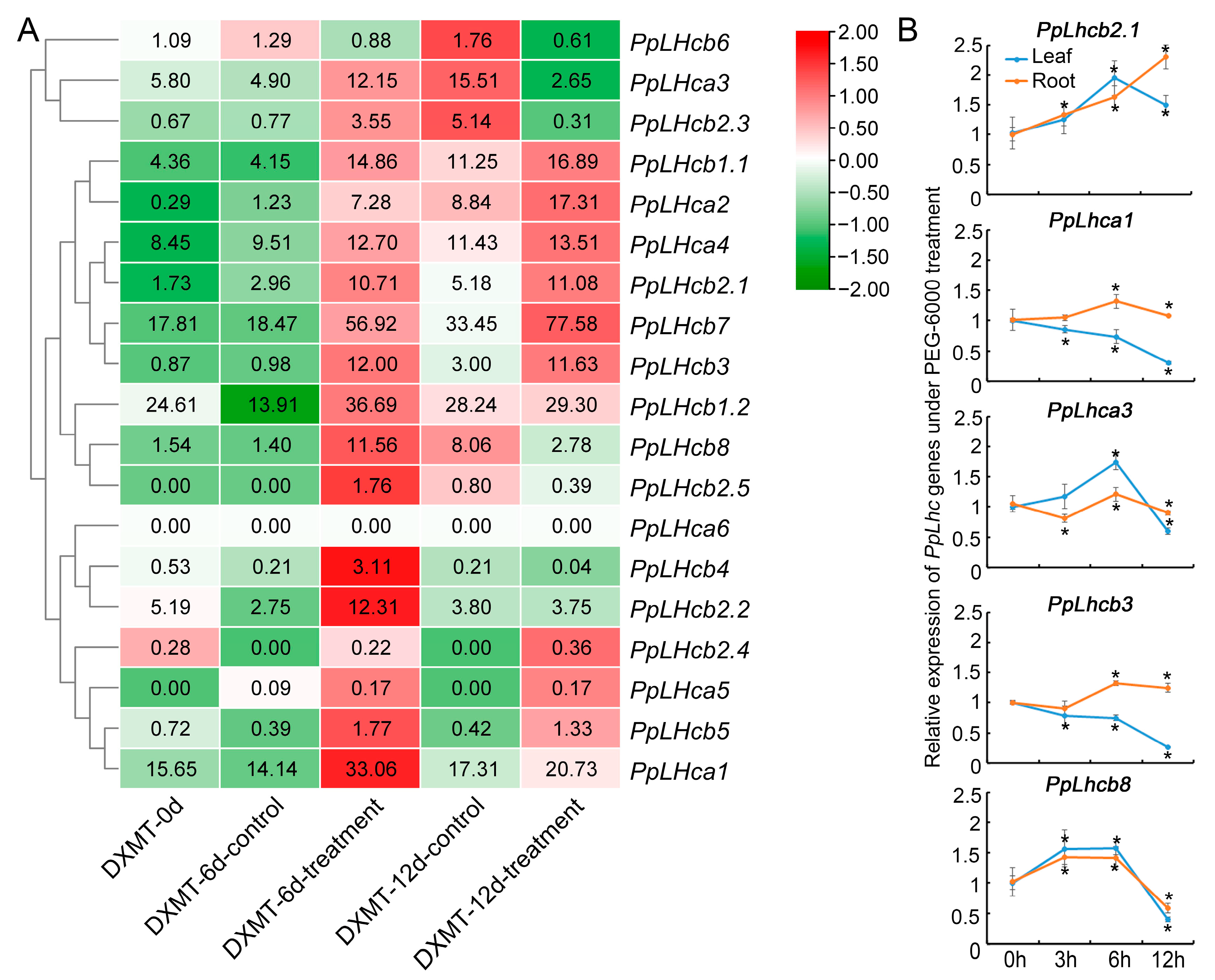
3.7. Expression Pattern Analysis of PpLhc Genes under Drought Stress
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bag, P.; Chukhutsina, V.; Zhang, Z.; Paul, S.; Ivanov, A.; Shutova, T.; Croce, R.; Holzwarth, A.; Jansson, S. Direct energy transfer from photosystem II to photosystem I confers winter sustainability in Scots Pine. Nat. Commun. 2020, 11, 6388. [Google Scholar] [CrossRef] [PubMed]
- Hey, D.; Grimm, B. ONE-HELIX PROTEIN2 (OHP2) Is Required for the Stability of OHP1 and Assembly Factor HCF244 and Is Functionally Linked to PSII Biogenesis. Plant Physiol. 2018, 177, 1453–1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietrzykowska, M.; Suorsa, M.; Semchonok, D.; Tikkanen, M.; Boekema, E.; Aro, E.; Jansson, S. The light-harvesting chlorophyll a/b binding proteins Lhcb1 and Lhcb2 play complementary roles during state transitions in Arabidopsis. Plant Cell 2014, 26, 3646–3660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, R.; Pichersky, E. Hypothesis for the evolution of three-helix Chl a/b and Chl a/c light-harvesting antenna proteins from two-helix and four-helix ancestors. Photosyn. Res. 1994, 39, 149–162. [Google Scholar] [CrossRef]
- Nelson, N.; Junge, W. Structure and energy transfer in photosystems of oxygenic photosynthesis. Annu. Rev. Biochem. 2015, 84, 659–683. [Google Scholar] [CrossRef]
- Jansson, S. A guide to the Lhc genes and their relatives in Arabidopsis. Trends Plant Sci. 1999, 4, 236–240. [Google Scholar] [CrossRef]
- Standfuss, J.; van Scheltinga, A.T.; Lamborghini, M.; Kühlbrandt, W. Mechanisms of photoprotection and nonphotochemical quenching in pea light-harvesting complex at 2.5 A resolution. EMBO J. 2005, 24, 919–928. [Google Scholar] [CrossRef] [Green Version]
- Klimmek, F.; Sjödin, A.; Noutsos, C.; Leister, D.; Jansson, S. Abundantly and rarely expressed Lhc protein genes exhibit distinct regulation patterns in plants. Plant Physiol. 2006, 140, 793–804. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Leebens-Mack, J.; Ayyampalayam, S.; Bowers, J.; McKain, M.; McNeal, J.; Rolf, M.; Ruzicka, D.; Wafula, E.; Wickett, N.; et al. A genome triplication associated with early diversification of the core eudicots. Genome Biol. 2012, 13, R3. [Google Scholar] [CrossRef] [Green Version]
- Kuhlbrandt, W.; Wang, D.N.; Fujiyoshi, Y. Atomic model of plant light-harvesting complex by electron crystallography. Nature 1994, 367, 614–621. [Google Scholar] [CrossRef]
- Rogl, H.; Kuhlbrandt, W. Mutant trimers of light-harvesting complex II exhibit altered pigment content and spectroscopic features. Biochemistry 1999, 38, 16214–16222. [Google Scholar] [CrossRef] [PubMed]
- Ben-Shem, A.; Frolow, F.; Nelson, N. Crystal structure of plant photosystem I. Nature 2003, 426, 630–635. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yan, H.; Wang, K.; Kuang, T.; Zhang, J.; Gui, L.; An, X.; Chang, W. Crystal structure of spinach major light-harvesting complex at 2.72 A resolution. Nature 2004, 428, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Suga, M.; Kuang, T.; Shen, J. Photosynthesis. Structural basis for energy transfer pathways in the plant PSI-LHCI supercomplex. Science 2015, 348, 989–995. [Google Scholar] [CrossRef]
- Liu, R.; Xu, Y.; Jiang, S.; Lu, K.; Lu, Y.; Feng, X.; Wu, Z.; Liang, S.; Yu, Y.; Wang, X.; et al. Light-harvesting chlorophyll a/b-binding proteins, positively involved in abscisic acid signalling, require a transcription repressor, WRKY40, to balance their function. J. Exp. Bot. 2013, 64, 5443–5456. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Abid, M.; Tu, J.; Gao, P.; Wang, Z.; Huang, H. Genome-Wide Identification of the LHC Gene Family in Kiwifruit and Regulatory Role of AcLhcb3.1/3.2 for Chlorophyll a Content. Int. J. Mol. Sci. 2022, 23, 6528. [Google Scholar] [CrossRef]
- Zhang, Q.; Ma, C.; Wang, X.; Ma, Q.; Fan, S.; Zhang, C. Genome-wide identification of the light-harvesting chlorophyll a/b binding (Lhc) family in Gossypium hirsutum reveals the influence of GhLhcb2.3 on chlorophyll a synthesis. Plant Biol. 2021, 23, 831–842. [Google Scholar] [CrossRef]
- Zhang, M.; Senoura, T.; Yang, X.; Chao, Y.; Nishizawa, N. Lhcb2 gene expression analysis in two ecotypes of Sedum alfredii subjected to Zn/Cd treatments with functional analysis of SaLhcb2 isolated from a Zn/Cd hyperaccumulator. Biotechnol. Lett. 2011, 33, 1865–1871. [Google Scholar] [CrossRef]
- Xia, Y.; Ning, Z.; Bai, G.; Li, R.; Yan, G.; Siddique, K.H.; Baum, M.; Guo, P. Allelic variations of a light harvesting chlorophyll a/b-binding protein gene (Lhcb1) associated with agronomic traits in barley. PLoS ONE 2012, 7, e37573. [Google Scholar] [CrossRef] [Green Version]
- Yadavalli, V.; Neelam, S.; Rao, A.; Reddy, A.; Subramanyam, R. Differential degradation of photosystem I subunits under iron deficiency in rice. J. Plant Physiol. 2012, 169, 753–759. [Google Scholar] [CrossRef]
- Jiang, Q.; Xu, Z.; Wang, F.; Li, M.; Ma, J.; Xiong, A. Effects of abiotic stresses on the expression of Lhcb1 gene and photosynthesis of Oenanthe javanica and Apium graveolens. Biol. Plant. 2014, 58, 256–264. [Google Scholar] [CrossRef]
- Xu, Y.H.; Liu, R.; Yan, L.; Liu, Z.Q.; Jiang, S.C.; Shen, Y.Y.; Wang, X.F.; Zhang, D.P. Light-harvesting chlorophyll a/b-binding proteins are required for stomatal response to abscisic acid in Arabidopsis. J. Exp. Bot. 2012, 63, 1095–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.-S.; Song, Z.-B.; Sun, Z.; Zhang, J.; Mei, Y.; Nian, H.J.; Li, K.Z.; Chen, L.M. Effects of Formaldehyde Stress on Physiological Characteristics and Gene Expression Associated with Photosynthesis in Arabidopsis thaliana. Plant Mol. Biol. Rep. 2012, 30, 1291–1302. [Google Scholar] [CrossRef]
- Deng, Y.; Kong, F.; Zhou, B.; Zhang, S.; Yue, M.; Meng, Q. Heterology expression of the tomato LeLhcb2 gene confers elevated tolerance to chilling stress in transgenic tobacco. Plant Physiol. Biochem. 2014, 80, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Gao, H.; Luo, J.; Wang, H.; Dong, Q.; Wang, Y.; Yang, K.; Mao, K.; Ma, F. Genome-wide analysis of the light-harvesting chlorophyll a/b-binding gene family in apple (Malus domestica) and functional characterization of MdLhcb4.3, which confers tolerance to drought and osmotic stress. Plant Physiol. Biochem. 2020, 154, 517–529. [Google Scholar] [CrossRef]
- Chen, L.; Yang, W.; Liu, S.; Meng, Y.; Zhu, Z.; Liang, R.; Cao, K.; Xie, Y.; Li, X. Genome-wide analysis and identification of light-harvesting chlorophyll a/b binding (LHC) gene family and BSMV-VIGS silencing TaLHC86 reduced salt tolerance in wheat. Int. J. Biol. Macromol. 2023, 242, 124930. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Hung, J.H.; Weng, Z. Sequence Alignment and Homology Search with BLAST and ClustalW. Cold Spring Harb. Protoc. 2016, 11, 1016–1021. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Brown, J.; Pirrung, M.; McCue, L.A. FQC Dashboard: Integrates FastQC results into a web-based, interactive, and extensible FASTQ quality control tool. Bioinformatics 2017, 33, 3137–3139. [Google Scholar] [CrossRef] [Green Version]
- Sewe, S.O.; Silva, G.; Sicat, P.; Seal, S.E.; Visendi, P. Trimming and Validation of Illumina Short Reads Using Trimmomatic, Trinity Assembly, and Assessment of RNA-Seq Data. Methods Mol. Biol. 2022, 2443, 211–232. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Tan, Q.; Sun, M.; Li, D.; Fu, X.; Chen, X.; Xiao, W.; Li, L.; Gao, D. Genome-wide identification of WRKY family genes in peach and analysis of WRKY expression during bud dormancy. Mol. Genet. Genom. 2016, 291, 1319–1332. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- MEHARI Teame Gereziher; Yanchao, X.; Odongo, M.R.; Jawad, U.M.; Nyangasi, K.J.; Xiaoyan, C.; Yuqing, H.; Yuhong, W.; Shuxun, Y.; Kunbo, W.; et al. Genome wide identification and characterization of light-harvesting Chloro a/b binding (LHC) genes reveals their potential role in enhancing drought tolerance in Gossypium hirsutum. J. Cotton Res. 2021, 4, 15. [Google Scholar] [CrossRef]
- Magwanga, R.; Kirungu, J.; Lu, P.; Yang, X.; Dong, Q.; Cai, X.; Xu, Y.; Wang, X.; Zhou, Z.; Hou, Y.; et al. Genome wide identification of the trihelix transcription factors and overexpression of Gh_A05G2067 (GT-2), a novel gene contributing to increased drought and salt stresses tolerance in cotton. Physiol. Plant. 2019, 167, 447–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umate, P. Genome-wide analysis of the family of light-harvesting chlorophyll a/b-binding proteins in Arabidopsis and rice. Plant Signal. Behav. 2010, 5, 1537–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Z.; Xiao, Y.; Zhang, L.; Zhao, Y. Analysis of Lhc family genes reveals development regulation and diurnal fluctuation expression patterns in Cyperus esculentus, a Cyperaceae plant. Planta 2023, 257, 59. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Ran, K.; Zhao, S.; Cheng, F. Genome-Wide Identification of the Light-Harvesting Chlorophyll a/b Binding Protein Gene Family in Pyrus bretschneideri and Their Transcriptomic Features under Drought Stress. Horticulturae 2022, 9, 522. [Google Scholar] [CrossRef]
- Wei, Y.; Lu, X.; Bao, J.; Zhang, C.; Yan, H.; Li, K.; Gong, M.; Li, S.; Ma, S. Identification and expression analysis of chlorophyll a/b binding protein gene family in grape (Vitis vinifera). Physiol. Mol. Biol. Plants 2022, 28, 1147–1158. [Google Scholar] [CrossRef]
- Zou, Z.; Li, M.; Jia, R.; Zhao, H.; He, P.; Zhang, Y.; Guo, A. Genes encoding light-harvesting chlorophyll a/b-binding proteins in papaya (Carica papaya L.) and insight into lineage-specific evolution in Brassicaceae. Gene 2020, 748, 144685. [Google Scholar] [CrossRef]
- Yan, M.; Yuan, Z. Genome-wide analysis of the family of light-harvesting chlorophyll a/b-binding proteins in pomegranate (Punica granatum L.). In Proceedings of the International Horticultural Congress IHC2018: V International Symposium on Plant Genetic Resources and International Symposium on Applied Functional Molecular Biology, Istanbul, Turkey, 12–16 August 2020; Volume 1297. [Google Scholar] [CrossRef]
- Li, X.; Zhu, Y.; Chen, C.; Geng, Z.; Li, X.; Ye, T.; Mao, X.; Du, F. Cloning and characterization of two chlorophyll A/B binding protein genes and analysis of their gene family in Camellia sinensis. Sci. Rep. 2020, 10, 4602. [Google Scholar] [CrossRef] [Green Version]
- Zou, Z.; Yang, J. Genomics analysis of the light-harvesting chlorophyll a/b-binding (Lhc) superfamily in cassava (Manihot esculenta Crantz). Gene 2019, 702, 171–181. [Google Scholar] [CrossRef]
- Cao, P.B.; Tran, T.T.H.; Nguyen, V.D.; La, V.H.; Azar, S. Genome-scale Identification and Analysis of Genes Encoding Putative Light-harvesting Chlorophyll a/b-binding Proteins in Potato (Solanum tuberosum L.). Chiang Mai J. Sci. 2019, 46, 867–879. [Google Scholar]
- Kong, F.; Zhou, Y.; Sun, P.P.; Cao, M.; Li, H.; Mao, Y.X. Identification of Light-Harvesting Chlorophyll a/b-Binding Protein Genes of Zostera marina L. and Their Expression Under Different Environmental Conditions. J. Ocean Univ. China 2016, 15, 152–162. [Google Scholar] [CrossRef]
- Gao, L.; Li, H.M. Identification of a Light-Harvesting Chlorophyll a/b-Binding Protein Gene in Gardenia jasminoides. Aer. Adv. Eng. Res. 2015, 22, 142–145. [Google Scholar]
- Zou, Z.; An, F. Genome-wide Identification, Classification and Expression Analysis of Lhc Supergene Family in Castor Bean (Ricinus communis L.). Agri. Biotechnol. 2013, 6, 44–51. [Google Scholar]
- Qin, D.; Xu, F.; Ge, S.; Xu, Q.; Li, M. Genome-wide identification and characterization of light harvesting chlorophyll a/b binding protein genes in barley (Hordeum vulgare L.). Adv. Crop Sci. Technol. 2017, 5, 301. [Google Scholar] [CrossRef]
- Shen, X.-X.; Salichos, L.; Rokas, A. A Genome-Scale Investigation of How Sequence, Function, and Tree-Based Gene Properties Influence Phylogenetic Inference. Genome Biol. Evol. 2016, 8, 2565–2580. [Google Scholar] [CrossRef] [Green Version]
- Gabaldon, T.; Koonin, E.V. Functional and evolutionary implications of gene orthology. Nat. Rev. Genet. 2013, 14, 360–366. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Yang, Y.; Jiang, L.; Liu, S. The catalase gene family in cucumber: Genome-wide identification and organization. Genet. Mol. Biol. 2016, 39, 408–415. [Google Scholar] [CrossRef] [Green Version]
- Peterson, R.; Schultes, N. Light-harvesting complex B7 shifts the irradiance response of photosynthetic light-harvesting regulation in leaves of Arabidopsis thaliana. J. Plant Physiol. 2014, 171, 311–318. [Google Scholar] [CrossRef]
- Saito, A.; Iino, T.; Sonoike, K.; Miwa, E.; Higuchi, K. Remodeling of the Major Light-Harvesting Antenna Protein of PSII Protects the Young Leaves of Barley (Hordeum vulgare L.) from Photoinhibition under Prolonged Iron Deficiency. Plant Cell Physiol. 2010, 51, 2013–2030. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Locus ID | Chr | Gene Location | Protein Length (AA) | Molecular Weight (kDa) | Theoretical pI | Aliphatic Index | GRAVY |
|---|---|---|---|---|---|---|---|---|
| PpLhca1 | Prupe.8G142700 | 8 | 15990280…15992121 forward | 247 | 26.72 | 6.11 | 83.36 | −0.10 |
| PpLhca2 | Prupe.1G225700 | 1 | 23908446…23910213 reverse | 272 | 29.15 | 6.42 | 77.61 | −0.05 |
| PpLhca3 | Prupe.3G191900 | 3 | 20429662…20431261 forward | 276 | 29.75 | 7.85 | 85.94 | −0.05 |
| PpLhca4 | Prupe.5G195100 | 5 | 15839308…15840574 reverse | 255 | 28.17 | 6.91 | 86.04 | −0.09 |
| PpLhca5 | Prupe.1G435900 | 1 | 37201367…37203304 forward | 266 | 28.84 | 7.13 | 97.93 | 0.15 |
| PpLhca6 | Prupe.7G195500 | 7 | 18520951…18522728 forward | 267 | 29.91 | 6.12 | 81.12 | −0.10 |
| PpLhcb1.1 | Prupe.4G076200 | 4 | 3725892…3726970 reverse | 264 | 28.03 | 5.14 | 79.17 | −0.01 |
| PpLhcb1.2 | Prupe.4G076300 | 4 | 3728016…3728975 forward | 265 | 28.13 | 5.14 | 78.87 | −0.01 |
| PpLhcb2.1 | Prupe.3G201000 | 3 | 20963264…20964787 forward | 265 | 28.48 | 5.47 | 81.02 | −0.01 |
| PpLhcb2.2 | Prupe.3G004100 | 3 | 271589…272612 forward | 265 | 28.33 | 5.29 | 78.11 | −0.04 |
| PpLhcb2.3 | Prupe.3G174600 | 3 | 19179155…19179985 reverse | 267 | 28.33 | 5.3 | 81.16 | −0.02 |
| PpLhcb2.4 | Prupe.3G174700 | 3 | 19181948…19183068 reverse | 267 | 28.33 | 5.3 | 81.16 | −0.01 |
| PpLhcb2.5 | Prupe.3G174800 | 3 | 19184322…19185343 reverse | 208 | 22.59 | 5.93 | 74.62 | −0.20 |
| PpLhcb3 | Prupe.4G190500 | 4 | 11367294…11369253 reverse | 269 | 29.12 | 5.11 | 84.5 | 0.02 |
| PpLhcb4 | Prupe.6G276200 | 6 | 25997046…25998631 reverse | 286 | 31.20 | 5.46 | 81.61 | −0.16 |
| PpLhcb5 | Prupe.8G001200 | 8 | 114709…116781 forward | 290 | 30.92 | 5.73 | 90.31 | 0.00 |
| PpLhcb6 | Prupe.1G557800 | 1 | 45538766…45538766 reverse | 259 | 27.68 | 7.78 | 81.12 | 0.04 |
| PpLhcb7 | Prupe.5G002700 | 5 | 404568…408672 reverse | 327 | 36.16 | 7.7 | 101.1 | 0.09 |
| PpLhcb8 | Prupe.2G248900 | 2 | 26441288…26443733 reverse | 282 | 30.87 | 5.15 | 81.31 | −0.12 |
| Protein | Subcellular Localization Prediction | TM Prediction | ||
|---|---|---|---|---|
| SoftBerry | Predotar | WoLF PROST | DeepTMHMM | |
| PpLhcb6 | Membrane-bound Chloroplast | Possibly plastid | Chloroplast | 3 |
| PpLhca6 | Membrane-bound Chloroplast | Possibly plastid | Chloroplast | 3 |
| PpLhca2 | Membrane-bound Chloroplast | Plastid | Chloroplast | 1 |
| PpLhca5 | Membrane-bound Chloroplast | none | Chloroplast | 3 |
| PpLhcb4 | Membrane-bound Chloroplast | Plastid | Chloroplast | 3 |
| PpLhcb8 | Membrane-bound Chloroplast | Plastid | Plasma membrane | 3 |
| PpLhca4 | Membrane-bound Chloroplast | Plastid | Chloroplast | 3 |
| PpLhca3 | Membrane-bound Chloroplast | Plastid | Chloroplast | 3 |
| PpLhcb3 | Membrane-bound Chloroplast | Plastid | Chloroplast | 3 |
| PpLhca1 | Membrane-bound Chloroplast | Plastid | Chloroplast | 3 |
| PpLhcb2.3 | Membrane-bound Chloroplast | Plastid | Chloroplast | 3 |
| PpLhcb2.4 | Membrane-bound Chloroplast | Plastid | Chloroplast | 3 |
| PpLhcb1.2 | Membrane-bound Chloroplast | Plastid | Chloroplast | 3 |
| PpLhcb1.1 | Membrane-bound Chloroplast | Plastid | Chloroplast | 3 |
| PpLhcb2.2 | Membrane-bound Chloroplast | Plastid | Chloroplast | 3 |
| PpLhcb5 | Membrane-bound Chloroplast | Plastid | Chloroplast | 3 |
| PpLhcb2.1 | Membrane-bound Chloroplast | Plastid | Chloroplast | 3 |
| PpLhcb7 | Membrane-bound Chloroplast | Plastid | Plasma membrane | 1 |
| PpLhcb2.5 | Membrane-bound Chloroplast | Plastid | Chloroplast | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Wei, J.; Shi, X.; Qian, W.; Mehmood, J.; Yin, Y.; Jia, H. Identification of the Light-Harvesting Chlorophyll a/b Binding Protein Gene Family in Peach (Prunus persica L.) and Their Expression under Drought Stress. Genes 2023, 14, 1475. https://doi.org/10.3390/genes14071475
Wang L, Wei J, Shi X, Qian W, Mehmood J, Yin Y, Jia H. Identification of the Light-Harvesting Chlorophyll a/b Binding Protein Gene Family in Peach (Prunus persica L.) and Their Expression under Drought Stress. Genes. 2023; 14(7):1475. https://doi.org/10.3390/genes14071475
Chicago/Turabian StyleWang, Li, Jia Wei, Xingyun Shi, Weihong Qian, Jan Mehmood, Yiming Yin, and Huijuan Jia. 2023. "Identification of the Light-Harvesting Chlorophyll a/b Binding Protein Gene Family in Peach (Prunus persica L.) and Their Expression under Drought Stress" Genes 14, no. 7: 1475. https://doi.org/10.3390/genes14071475