
Mutations in Structural Genes of the Mitochondrial Complex IV May Influence Breast Cancer


Abstract
:1. Introduction
2. Materials and Methods
2.1. Characterization of the Study
2.2. Study Design
2.3. Sampling from Public Databases
2.3.1. Analysis of Variants in Mitochondrial Genes (P1)
2.3.2. Analysis of Variants in Nuclear Genes (P2)
2.4. Selected Samples
2.5. In Silico Association and Functional Enrichment
2.6. Statistical Analyses
2.7. Pathogenicity of Characterized Variants
3. Results and Discussion
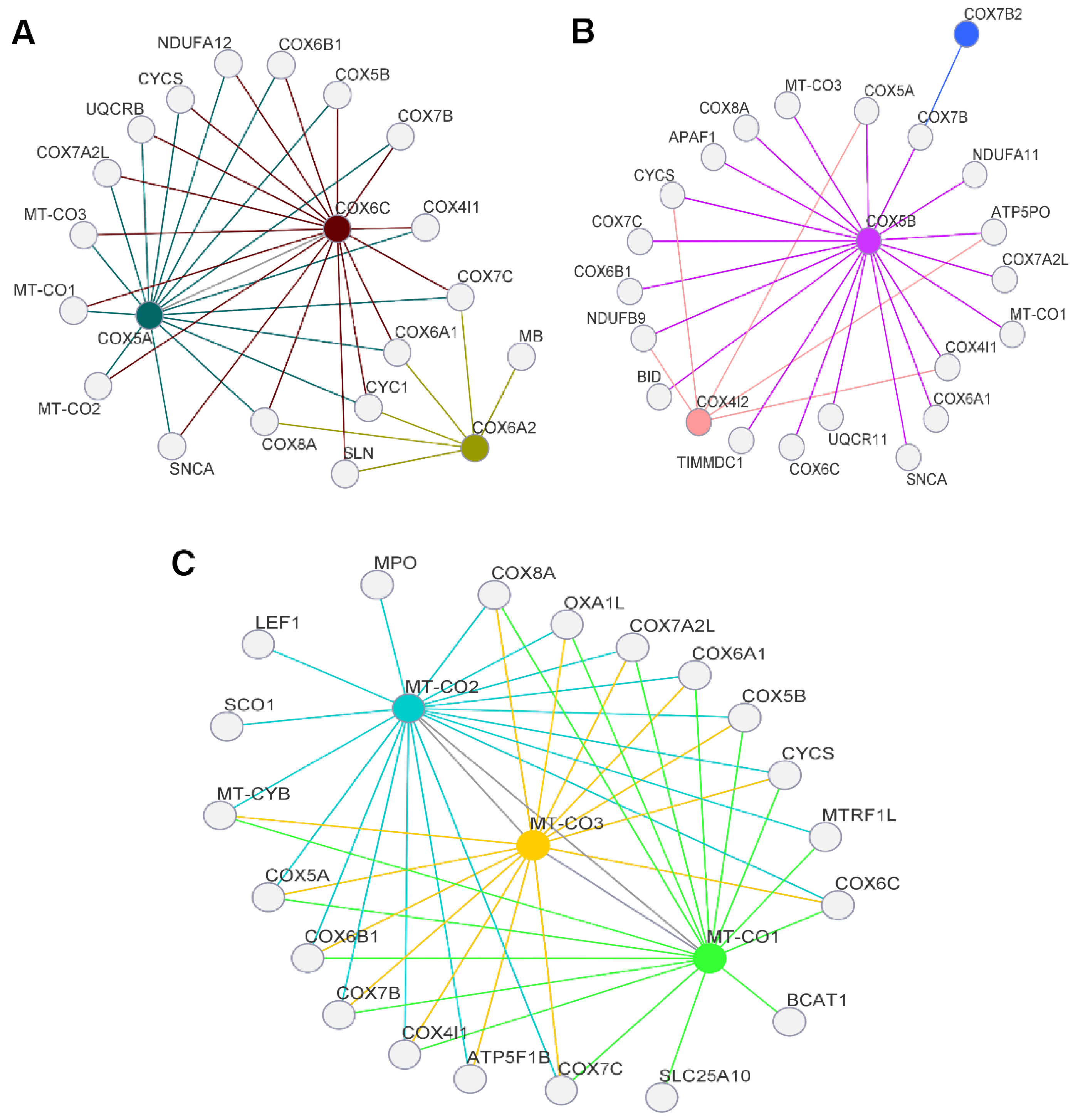
3.1. Presence of Alterations in Mitochondrial Genes
3.2. Presence of Alterations in Nuclear Genes
3.3. Clinical Characterization of Nuclear Gene Databases
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Binotto, M.; Schwartsmann, G. Qualidade de Vida Relacionada à Saúde de Pacientes Com Câncer de Mama: Revisão Integrativa Da Literatura. Rev. Bras. Cancerol. 2020, 66, e-06405. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer Statistics for the Year 2020: An Overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, D.; Pal, D.; Sharma, R.; Garg, V.K.; Goel, N.; Koundal, D.; Zaguia, A.; Koundal, S.; Belay, A. Global Increase in Breast Cancer Incidence: Risk Factors and Preventive Measures. BioMed Res. Int. 2022, 2022, 9605439. [Google Scholar] [CrossRef]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular Principles of Metastasis: A Hallmark of Cancer Revisited. Signal Transduct. Target. Ther. 2020, 5, 28. [Google Scholar] [CrossRef]
- Warburg, O. The Metabolism of Carcinoma Cells. J. Cancer Res. 1925, 9, 148–163. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J. Energy Metabolism of Cancer: Glycolysis versus Oxidative Phosphorylation (Review). Oncol. Lett. 2012, 4, 1151–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Icard, P.; Shulman, S.; Farhat, D.; Steyaert, J.-M.; Alifano, M.; Lincet, H. How the Warburg Effect Supports Aggressiveness and Drug Resistance of Cancer Cells? Drug Resist. Updates 2018, 38, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Enríquez, S.; Carreño-Fuentes, L.; Gallardo-Pérez, J.C.; Saavedra, E.; Quezada, H.; Vega, A.; Marín-Hernández, A.; Olín-Sandoval, V.; Torres-Márquez, M.E.; Moreno-Sánchez, R. Oxidative Phosphorylation Is Impaired by Prolonged Hypoxia in Breast and Possibly in Cervix Carcinoma. Int. J. Biochem. Cell Biol. 2010, 42, 1744–1751. [Google Scholar] [CrossRef] [PubMed]
- Whitaker-Menezes, D.; Martinez-Outschoorn, U.E.; Flomenberg, N.; Birbe, R.; Witkiewicz, A.K.; Howell, A.; Pavlides, S.; Tsirigos, A.; Ertel, A.; Pestell, R.G.; et al. Hyperactivation of Oxidative Mitochondrial Metabolism in Epithelial Cancer Cells in Situ: Visualizing the Therapeutic Effects of Metformin in Tumor Tissue. Cell Cycle 2011, 10, 4047–4064. [Google Scholar] [CrossRef] [Green Version]
- Cazzaniga, M.; Bonanni, B. Relationship Between Metabolic Reprogramming and Mitochondrial Activity in Cancer Cells. Understanding The Anticancer Effect of Metformin and Its Clinical Implications. Anticancer Res. 2015, 35, 5789–5796. [Google Scholar] [PubMed]
- Giampazolias, E.; Tait, S.W.G. Mitochondria and the Hallmarks of Cancer. FEBS J. 2016, 283, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Xiang, L.; Li, T.; Bai, Z. Cancer Hallmarks, Biomarkers and Breast Cancer Molecular Subtypes. J. Cancer 2016, 7, 1281–1294. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, S.; Guha, M.; Dong, D.W.; Whelan, K.A.; Ruthel, G.; Uchikado, Y.; Natsugoe, S.; Nakagawa, H.; Avadhani, N.G. Disruption of Cytochrome c Oxidase Function Induces the Warburg Effect and Metabolic Reprogramming. Oncogene 2016, 35, 1585–1595. [Google Scholar] [CrossRef] [Green Version]
- Kadenbach, B.; Ramzan, R.; Vogt, S. High Efficiency versus Maximal Performance—The Cause of Oxidative Stress in Eukaryotes: A Hypothesis. Mitochondrion 2013, 13, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kadenbach, B. Complex IV—The Regulatory Center of Mitochondrial Oxidative Phosphorylation. Mitochondrion 2021, 58, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Bender, D.A. Oxidative Phosphorylation. In Encyclopedia of Food Sciences and Nutrition; Elsevier: Amsterdam, The Netherlands, 2003; pp. 4295–4301. ISBN 9780122270550. [Google Scholar]
- Cardol, P.; Figueroa, F.; Remacle, C.; Franzén, L.-G.; González-Halphen, D. Oxidative Phosphorylation. In the Chlamydomonas Sourcebook; Elsevier: Amsterdam, The Netherlands, 2009; pp. 469–502. ISBN 9780123708731. [Google Scholar]
- Kadenbach, B.; Hüttemann, M. The Subunit Composition and Function of Mammalian Cytochrome c Oxidase. Mitochondrion 2015, 24, 64–76. [Google Scholar] [CrossRef]
- Hu, Y.; Xu, W.; Zeng, H.; He, Z.; Lu, X.; Zuo, D.; Qin, G.; Chen, W. OXPHOS-Dependent Metabolic Reprogramming Prompts Metastatic Potential of Breast Cancer Cells under Osteogenic Differentiation. Br. J. Cancer 2020, 123, 1644–1655. [Google Scholar] [CrossRef] [PubMed]
- Putignani, L.; Raffa, S.; Pescosolido, R.; Aimati, L.; Signore, F.; Torrisi, M.R.; Grammatico, P. Alteration of Expression Levels of the Oxidative Phosphorylation System (OXPHOS) in Breast Cancer Cell Mitochondria. Breast Cancer Res. Treat. 2008, 110, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Yim, A.; Koti, P.; Bonnard, A.; Marchiano, F.; Dürrbaum, M.; Garcia-Perez, C.; Villaveces, J.; Gamal, S.; Cardone, G.; Perocchi, F.; et al. mitoXplorer, a Visual Data Mining Platform to Systematically Analyze and Visualize Mitochondrial Expression Dynamics and Mutations. Nucleic Acids Res. 2020, 48, 605–632. [Google Scholar] [CrossRef] [Green Version]
- Brischigliaro, M.; Zeviani, M. Cytochrome c Oxidase Deficiency. Biochim. Biophys. Acta (BBA)-Bioenerg. 2021, 1862, 148335. [Google Scholar] [CrossRef]
- Fornuskova, D.; Stiburek, L.; Wenchich, L.; Vinsova, K.; Hansikova, H.; Zeman, J. Novel Insights into the Assembly and Function of Human Nuclear-Encoded Cytochrome c Oxidase Subunits 4, 5a, 6a, 7a and 7b. Biochem. J. 2010, 428, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Timón-Gómez, A.; Bartley-Dier, E.L.; Fontanesi, F.; Barrientos, A. HIGD-Driven Regulation of Cytochrome c Oxidase Biogenesis and Function. Cells 2020, 9, 2620. [Google Scholar] [CrossRef]
- Pitceathly, R.D.S.; Taanman, J.-W. NDUFA4 (Renamed COXFA4) Is a Cytochrome-c Oxidase Subunit. Trends Endocrinol. Metab. 2018, 29, 452–454. [Google Scholar] [CrossRef]
- TCMA. Available online: https://ibl.mdanderson.org/tcma/about.html (accessed on 18 March 2023).
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative Analysis of Complex Cancer Genomics and Clinical Profiles Using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Ju, Y.S.; Kim, Y.; Li, J.; Wang, Y.; Yoon, C.J.; Yang, Y.; Martincorena, I.; Creighton, C.J.; Weinstein, J.N.; et al. Comprehensive Molecular Characterization of Mitochondrial Genomes in Human Cancers. Nat. Genet. 2020, 52, 342–352. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.P.; Roth, A.; Goya, R.; Oloumi, A.; Ha, G.; Zhao, Y.; Turashvili, G.; Ding, J.; Tse, K.; Haffari, G.; et al. The Clonal and Mutational Evolution Spectrum of Primary Triple-Negative Breast Cancers. Nature 2012, 486, 395–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerji, S.; Cibulskis, K.; Rangel-Escareno, C.; Brown, K.K.; Carter, S.L.; Frederick, A.M.; Lawrence, M.S.; Sivachenko, A.Y.; Sougnez, C.; Zou, L.; et al. Sequence Analysis of Mutations and Translocations across Breast Cancer Subtypes. Nature 2012, 486, 405–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Oslo Breast Cancer Consortium (OSBREAC); Stephens, P.J.; Tarpey, P.S.; Davies, H.; Van Loo, P.; Greenman, C.; Wedge, D.C.; Nik-Zainal, S.; Martin, S.; Varela, I.; et al. The Landscape of Cancer Genes and Mutational Processes in Breast Cancer. Nature 2012, 486, 400–404. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, C.; Bachelot, T.; Filleron, T.; Pedrero, M.; Campone, M.; Soria, J.-C.; Massard, C.; Lévy, C.; Arnedos, M.; Lacroix-Triki, M.; et al. Mutational Profile of Metastatic Breast Cancers: A Retrospective Analysis. PLoS Med. 2016, 13, e1002201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoadley, K.A.; Yau, C.; Hinoue, T.; Wolf, D.M.; Lazar, A.J.; Drill, E.; Shen, R.; Taylor, A.M.; Cherniack, A.D.; Thorsson, V.; et al. Cell-of-Origin Patterns Dominate the Molecular Classification of 10,000 Tumors from 33 Types of Cancer. Cell 2018, 173, 291–304.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellrott, K.; Bailey, M.H.; Saksena, G.; Covington, K.R.; Kandoth, C.; Stewart, C.; Hess, J.; Ma, S.; Chiotti, K.E.; McLellan, M.; et al. Scalable Open Science Approach for Mutation Calling of Tumor Exomes Using Multiple Genomic Pipelines. Cell Syst. 2018, 6, 271–281.e7. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.M.; Shih, J.; Ha, G.; Gao, G.F.; Zhang, X.; Berger, A.C.; Schumacher, S.E.; Wang, C.; Hu, H.; Liu, J.; et al. Genomic and Functional Approaches to Understanding Cancer Aneuploidy. Cancer Cell 2018, 33, 676–689.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Q.; Liang, W.-W.; Foltz, S.M.; Mutharasu, G.; Jayasinghe, R.G.; Cao, S.; Liao, W.-W.; Reynolds, S.M.; Wyczalkowski, M.A.; Yao, L.; et al. Driver Fusions and Their Implications in the Development and Treatment of Human Cancers. Cell Rep. 2018, 23, 227–238.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Lichtenberg, T.; Hoadley, K.A.; Poisson, L.M.; Lazar, A.J.; Cherniack, A.D.; Kovatich, A.J.; Benz, C.C.; Levine, D.A.; Lee, A.V.; et al. An Integrated TCGA Pan-Cancer Clinical Data Resource to Drive High-Quality Survival Outcome Analytics. Cell 2018, 173, 400–416.e11. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Vega, F.; Mina, M.; Armenia, J.; Chatila, W.K.; Luna, A.; La, K.C.; Dimitriadoy, S.; Liu, D.L.; Kantheti, H.S.; Saghafinia, S.; et al. Oncogenic Signaling Pathways in The Cancer Genome Atlas. Cell 2018, 173, 321–337.e10. [Google Scholar] [CrossRef] [Green Version]
- Poore, G.D.; Kopylova, E.; Zhu, Q.; Carpenter, C.; Fraraccio, S.; Wandro, S.; Kosciolek, T.; Janssen, S.; Metcalf, J.; Song, S.J.; et al. Microbiome Analyses of Blood and Tissues Suggest Cancer Diagnostic Approach. Nature 2020, 579, 567–574. [Google Scholar] [CrossRef]
- Bhandari, V.; Hoey, C.; Liu, L.Y.; Lalonde, E.; Ray, J.; Livingstone, J.; Lesurf, R.; Shiah, Y.-J.; Vujcic, T.; Huang, X.; et al. Molecular Landmarks of Tumor Hypoxia across Cancer Types. Nat. Genet. 2019, 51, 308–318. [Google Scholar] [CrossRef]
- Ding, L.; Bailey, M.H.; Porta-Pardo, E.; Thorsson, V.; Colaprico, A.; Bertrand, D.; Gibbs, D.L.; Weerasinghe, A.; Huang, K.; Tokheim, C.; et al. Perspective on Oncogenic Processes at the End of the Beginning of Cancer Genomics. Cell 2018, 173, 305–320.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonneville, R.; Krook, M.A.; Kautto, E.A.; Miya, J.; Wing, M.R.; Chen, H.-Z.; Reeser, J.W.; Yu, L.; Roychowdhury, S. Landscape of Microsatellite Instability Across 39 Cancer Types. JCO Precis. Oncol. 2017, 1, 1–15. [Google Scholar] [CrossRef]
- Bonneville, R.; Paruchuri, A.; Wing, M.R.; Krook, M.A.; Reeser, J.W.; Chen, H.-Z.; Dao, T.; Samorodnitsky, E.; Smith, A.M.; Yu, L.; et al. Characterization of Clonal Evolution in Microsatellite Unstable Metastatic Cancers through Multiregional Tumor Sequencing. Mol. Cancer Res. 2021, 19, 465–474. [Google Scholar] [CrossRef]
- Wagle, N.; Painter, C.; Krevalin, M.; Oh, C.; Anderka, K.; Larkin, K.; Lennon, N.; Dillon, D.; Frank, E.; Winer, E.P.; et al. The Metastatic Breast Cancer Project: A National Direct-to-Patient Initiative to Accelerate Genomics Research. JCO 2016, 34, LBA1519. [Google Scholar] [CrossRef]
- Wagle, N.; Painter, C.; Anastasio, E.; Dunphy, M.; McGillicuddy, M.; Kim, D.; Jain, E.; Buendia-Buendia, J.; Cohen, O.; Knelson, E.; et al. The Metastatic Breast Cancer (MBC) Project: Accelerating Translational Research through Direct Patient Engagement. JCO 2017, 35, 1076. [Google Scholar] [CrossRef]
- The Metastatic Breast Cancer Project. Available online: https://mbcproject.org/data-release (accessed on 18 March 2023).
- Krug, K.; Jaehnig, E.J.; Satpathy, S.; Blumenberg, L.; Karpova, A.; Anurag, M.; Miles, G.; Mertins, P.; Geffen, Y.; Tang, L.C.; et al. Proteogenomic Landscape of Breast Cancer Tumorigenesis and Targeted Therapy. Cell 2020, 183, 1436–1456.e31. [Google Scholar] [CrossRef]
- Singh, D.; Garg, R. R, Language for Data Analytics. In Proceedings of the International Conference on Sustainable Computing in Science, Technology and Management (SUSCOM), Jaipur, India, 26–28 February 2019. [Google Scholar]
- Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the Unification of Biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- The Gene Ontology Consortium; Carbon, S.; Douglass, E.; Good, B.M.; Unni, D.R.; Harris, N.L.; Mungall, C.J.; Basu, S.; Chisholm, R.L.; Dodson, R.J.; et al. The Gene Ontology Resource: Enriching a Gold Mine. Nucleic Acids Res. 2021, 49, D325–D334. [Google Scholar] [CrossRef]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA Prediction Server: Biological Network Integration for Gene Prioritization and Predicting Gene Function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chelala, C.; Khan, A.; Lemoine, N.R. SNPnexus: A Web Database for Functional Annotation of Newly Discovered and Public Domain Single Nucleotide Polymorphisms. Bioinformatics 2009, 25, 655–661. [Google Scholar] [CrossRef] [Green Version]
- JASP—A Fresh Way to Do Statistics. Available online: https://jasp-stats.org/ (accessed on 18 March 2023).
- Roland, N.; Porter, G.; Fish, B.; Makura, Z. Tumour Assessment and Staging: United Kingdom National Multidisciplinary Guidelines. J. Laryngol. Otol. 2016, 130, S53–S58. [Google Scholar] [CrossRef] [Green Version]
- Cancer Staging Systems. Available online: https://www.facs.org/quality-programs/cancer-programs/american-joint-committee-on-cancer/cancer-staging-systems/ (accessed on 18 March 2023).
- Ng, P.C. SIFT: Predicting Amino Acid Changes That Affect Protein Function. Nucleic Acids Res. 2003, 31, 3812–3814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adzhubei, I.; Jordan, D.M.; Sunyaev, S.R. Predicting Functional Effect of Human Missense Mutations Using PolyPhen-2. Curr. Protoc. Hum. Genet. 2013, 76, 7–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, S.M.; Riggs, E.R.; Maglott, D.R.; Lee, J.M.; Azzariti, D.R.; Niehaus, A.; Ramos, E.M.; Martin, C.L.; Landrum, M.J.; Rehm, H.L. Using ClinVar as a Resource to Support Variant Interpretation. Curr. Protoc. Hum. Genet. 2016, 89. [Google Scholar] [CrossRef] [Green Version]
- Karadimas, C.L.; Greenstein, P.; Sue, C.M.; Joseph, J.T.; Tanji, K.; Haller, R.G.; Taivassalo, T.; Davidson, M.M.; Shanske, S.; Bonilla, E.; et al. Recurrent Myoglobinuria Due to a Nonsense Mutation in the COX I Gene of Mitochondrial DNA. Neurology 2000, 55, 644–649. [Google Scholar] [CrossRef] [PubMed]
- Jaksch, M.; Hofmann, S.; Kleinle, S.; Liechti-Gallati, S.; Pongratz, D.E.; Muller-Hocker, J.; Jedele, K.B.; Meitinger, T.; Gerbitz, K.D. A Systematic Mutation Screen of 10 Nuclear and 25 Mitochondrial Candidate Genes in 21 Patients with Cytochrome c Oxidase (COX) Deficiency Shows tRNA(Ser)(UCN) Mutations in a Subgroup with Syndromal Encephalopathy. J. Med. Genet. 1998, 35, 895–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greaves, L.C.; Preston, S.L.; Tadrous, P.J.; Taylor, R.W.; Barron, M.J.; Oukrif, D.; Leedham, S.J.; Deheragoda, M.; Sasieni, P.; Novelli, M.R.; et al. Mitochondrial DNA Mutations Are Established in Human Colonic Stem Cells, and Mutated Clones Expand by Crypt Fission. Proc. Natl. Acad. Sci. USA 2006, 103, 714–719. [Google Scholar] [CrossRef]
- Namslauer, I.; Brzezinski, P. A Mitochondrial DNA Mutation Linked to Colon Cancer Results in Proton Leaks in Cytochrome c Oxidase. Proc. Natl. Acad. Sci. USA 2009, 106, 3402–3407. [Google Scholar] [CrossRef]
- Singh, R.K.; Saini, S.K.; Prakasam, G.; Kalairasan, P.; Bamezai, R.N.K. Role of Ectopically Expressed mtDNA Encoded Cytochrome c Oxidase Subunit I (MT-COI) in Tumorigenesis. Mitochondrion 2019, 49, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Danyu, L.; Yanran, L.; Xiuna, J.; Ying, C.; Sudan, P.; Tianen, Z.; Zhifen, Z.; Dezhi, Z.; Kaixun, H.; Yingyu, X.; et al. α-Synuclein Induced Mitochondrial Dysfunction via Cytochrome c Oxidase Subunit 2 in SH-SY5Y Cells. Exp. Cell Res. 2019, 378, 57–65. [Google Scholar] [CrossRef]
- Qattan, A.T.; Radulovic, M.; Crawford, M.; Godovac-Zimmermann, J. Spatial Distribution of Cellular Function: The Partitioning of Proteins between Mitochondria and the Nucleus in MCF7 Breast Cancer Cells. J. Proteome Res. 2012, 11, 6080–6101. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Giorgi, E.E.; Beckman, K.B.; Caberto, C.; Kazma, R.; Lum-Jones, A.; Haiman, C.A.; Marchand, L.L.; Stram, D.O.; Saxena, R.; et al. Association between Mitochondrial Genetic Variation and Breast Cancer Risk: The Multiethnic Cohort. PLoS ONE 2019, 14, e0222284. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Parveen, M.; Basgen, J.M.; Fazel, S.; Meshesha, M.F.; Thames, E.C.; Moore, B.; Martinez, L.; Howard, C.B.; Vergnes, L.; et al. Increased Expression of Beige/Brown Adipose Markers from Host and Breast Cancer Cells Influence Xenograft Formation in Mice. Mol. Cancer Res. 2016, 14, 78–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.C.; Crescitelli, R.; Cvjetkovic, A.; Belgrano, V.; Olofsson Bagge, R.; Sundfeldt, K.; Ochiya, T.; Kalluri, R.; Lötvall, J. Mitochondrial Protein Enriched Extracellular Vesicles Discovered in Human Melanoma Tissues Can Be Detected in Patient Plasma. J. Extracell. Vesicles 2019, 8, 1635420. [Google Scholar] [CrossRef] [Green Version]
- Johns, D.R.; Neufeld, M.J. Cytochrome c Oxidase Mutations in Leber Hereditary Optic Neuropathy. Biochem. Biophys. Res. Commun. 1993, 196, 810–815. [Google Scholar] [CrossRef]
- Johns, D.R.; Neufeld, M.J.; Hedges, T.R. Mitochondrial DNA Mutations in Cuban Optic and Peripheral Neuropathy. J. Neuro-Ophthalmol. 1994, 14, 135–140. [Google Scholar] [CrossRef]
- Oostra, R.J.; Van den Bogert, C.; Nijtmans, L.G.; van Galen, M.J.; Zwart, R.; Bolhuis, P.A.; Bleeker-Wagemakers, E.M. Simultaneous Occurrence of the 11778 (ND4) and the 9438 (COX III) mtDNA Mutations in Leber Hereditary Optic Neuropathy: Molecular, Biochemical, and Clinical Findings. Am. J. Hum. Genet. 1995, 57, 954–957. [Google Scholar]
- Tiranti, V. A Novel Frameshift Mutation of the mtDNA COIII Gene Leads to Impaired Assembly of Cytochrome c Oxidase in a Patient Affected by Leigh-like Syndrome. Hum. Mol. Genet. 2000, 9, 2733–2742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Księżakowska-Łakoma, K.; Kulczycka-Wojdala, D.; Kulig, A.; Baum, M.; Wilczyński, J.R. The Presence of A5935G, G5949A, G6081A, G6267A, T9540C Mutations in MT-CO1 and MT-CO3 Genes and Other Variants of MT-CO1 and MT-CO3 Gene Fragments in the Study Population Diagnosed with Endometrial Cancer. Ginekol. Pol. 2017, 88, 343–348. [Google Scholar] [CrossRef] [Green Version]
- Järviaho, T.; Hurme-Niiranen, A.; Soini, H.K.; Niinimäki, R.; Möttönen, M.; Savolainen, E.-R.; Hinttala, R.; Harila-Saari, A.; Uusimaa, J. Novel Non-Neutral Mitochondrial DNA Mutations Found in Childhood Acute Lymphoblastic Leukemia. Clin. Genet. 2018, 93, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, Y.; Zeng, S.; Gao, H.; Chen, Y.; Zou, X. Identification of COX4I2 as a Hypoxia-Associated Gene Acting through FGF1 to Promote EMT and Angiogenesis in CRC. Cell Mol. Biol. Lett. 2022, 27, 76. [Google Scholar] [CrossRef]
- Haghjoo, N.; Moeini, A.; Masoudi-Nejad, A. Introducing a Panel for Early Detection of Lung Adenocarcinoma by Using Data Integration of Genomics, Epigenomics, Transcriptomics and Proteomics. Exp. Mol. Pathol. 2020, 112, 104360. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.-P.; Sun, H.-F.; Jiang, H.-L.; Li, L.-D.; Hu, X.; Xu, X.-E.; Jin, W. Loss of COX5B Inhibits Proliferation and Promotes Senescence via Mitochondrial Dysfunction in Breast Cancer. Oncotarget 2015, 6, 43363–43374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.-P.; Sun, H.-F.; Fu, W.-Y.; Li, L.-D.; Zhao, Y.; Chen, M.-T.; Jin, W. High Expression of COX5B Is Associated with Poor Prognosis in Breast Cancer. Future Oncol. 2017, 13, 1711–1719. [Google Scholar] [CrossRef]
- Wang, Z.; Embaye, K.S.; Yang, Q.; Qin, L.; Zhang, C.; Liu, L.; Zhan, X.; Zhang, F.; Wang, X.; Qin, S. A Novel Metabolism-Related Signature as a Candidate Prognostic Biomarker for Hepatocellular Carcinoma. JHC 2021, 8, 119–132. [Google Scholar] [CrossRef]
- Liang, H. A Rare Polymorphism of the COX7B2 Gene in a Cantonese Family with Nasopharyngeal Carcinoma. Sci. China Ser. C Life Sci. 2004, 47, 449. [Google Scholar] [CrossRef]
- Ikeda, K.; Horie-Inoue, K.; Suzuki, T.; Hobo, R.; Nakasato, N.; Takeda, S.; Inoue, S. Mitochondrial Supercomplex Assembly Promotes Breast and Endometrial Tumorigenesis by Metabolic Alterations and Enhanced Hypoxia Tolerance. Nat. Commun. 2019, 10, 4108. [Google Scholar] [CrossRef] [Green Version]
- Muñiz Lino, M.A.; Palacios-Rodríguez, Y.; Rodríguez-Cuevas, S.; Bautista-Piña, V.; Marchat, L.A.; Ruíz-García, E.; Astudillo-de La Vega, H.; González-Santiago, A.E.; Flores-Pérez, A.; Díaz-Chávez, J.; et al. Comparative Proteomic Profiling of Triple-Negative Breast Cancer Reveals That up-Regulation of RhoGDI-2 Is Associated to the Inhibition of Caspase 3 and Caspase 9. J. Proteom. 2014, 111, 198–211. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Li, G.; Xia, Y.; Wang, F.; Wang, Y.; Xu, S.; Zhou, Y.; Liu, X.; Xie, X.; Zhang, J. miR-204/COX5A Axis Contributes to Invasion and Chemotherapy Resistance in Estrogen Receptor-Positive Breast Cancers. Cancer Lett. 2020, 492, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Reyimu, A.; Sun, A.; Duoji, Z.; Zhou, W.; Liang, S.; Hu, S.; Dai, W.; Xu, X. Establishment of Prognostic Risk Model and Drug Sensitivity Based on Prognostic Related Genes of Esophageal Cancer. Sci. Rep. 2022, 12, 8008. [Google Scholar] [CrossRef]
- Teerlink, C.C.; Miller, J.B.; Vance, E.L.; Staley, L.A.; Stevens, J.; Tavana, J.P.; Cloward, M.E.; Page, M.L.; Dayton, L.; Alzheimer’s Disease Genetics Consortium; et al. Analysis of High-risk Pedigrees Identifies 11 Candidate Variants for Alzheimer’s Disease. Alzheimer’s Dement. 2022, 18, 307–317. [Google Scholar] [CrossRef]
- Gruvberger, S.; Ringnér, M.; Chen, Y.; Panavally, S.; Saal, L.H.; Borg, A.; Fernö, M.; Peterson, C.; Meltzer, P.S. Estrogen Receptor Status in Breast Cancer Is Associated with Remarkably Distinct Gene Expression Patterns. Cancer Res. 2001, 61, 5979–5984. [Google Scholar] [PubMed]
- Chang, F.-W.; Fan, H.-C.; Liu, J.-M.; Fan, T.-P.; Jing, J.; Yang, C.-L.; Hsu, R.-J. Estrogen Enhances the Expression of the Multidrug Transporter Gene ABCG2—Increasing Drug Resistance of Breast Cancer Cells through Estrogen Receptors. IJMS 2017, 18, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, X.; Wang, P.; Yang, T.; Li, G.; Teng, X.; Huang, W.; Yu, H. Identification of Key Modules and Genes Associated with Breast Cancer Prognosis Using WGCNA and ceRNA Network Analysis. Aging 2021, 13, 2519–2538. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.-S.; Zhao, Z.; Yang, Z.-N.; Xu, F.; Lu, H.-J.; Zhu, Z.-Y.; Shi, W.; Jiang, J.; Yao, P.-P.; Zhu, H.-P. Risk Factors and Preventions of Breast Cancer. Int. J. Biol. Sci. 2017, 13, 1387–1397. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Gao, H.; Zhang, X.; Liu, Q.; Chen, G. Mutational and Transcriptional Alterations and Clinicopathological Factors Predict the Prognosis of Stage I Hepatocellular Carcinoma: Prediction of Stage I HCC Prognosis. BMC Gastroenterol. 2022, 22, 427. [Google Scholar] [CrossRef]
- Yersal, O.; Barutca, S. Biological Subtypes of Breast Cancer: Prognostic and Therapeutic Implications. World J. Clin. Oncol. 2014, 5, 412–424. [Google Scholar] [CrossRef] [PubMed]
- Makki, J. Diversity of Breast Carcinoma: Histological Subtypes and Clinical Relevance. Clin. Med. Insights Pathol. 2015, 8, Cpath-S31563. [Google Scholar] [CrossRef] [Green Version]
- Scholl, A.R.; Flanagan, M.B. Educational Case: Invasive Ductal Carcinoma of the Breast. Acad. Pathol. 2020, 7, 2374289519897390. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene | Name | Protein | Locus | Reference |
|---|---|---|---|---|
| Mitochondrially encoded cytochrome c oxidase I | MT-CO1 | COX1 | m.5904-7445 | [25] |
| Mitochondrially encoded cytochrome c oxidase II | MT-CO2 | COX2 | m.7586-8269 | [25] |
| Mitochondrially encoded cytochrome c oxidase III | MT-CO3 | COX3 | m.9207-9990 | [25] |
| Cytochrome c oxidase subunit 4I1 | COX4I1 | COX4 | 16q24.1 | [26] |
| Cytochrome c oxidase subunit 4I2 | COX4I2 | COX4 | 20q11.21 | [26] |
| Cytochrome c oxidase subunit 5A | COX5A | COX5A | 15q24.2 | [25] |
| Cytochrome c oxidase subunit 5B | COX5B | COX5B | 2.11.2 | [27] |
| Cytochrome c oxidase subunit 6A1 | COX6A1 | COX6A1 or COX6A | 12q24.31 | [27] |
| Cytochrome c oxidase subunit 6A2 | COX6A2 | COX6A2 or COX6A | 16p11.2 | [27] |
| Cytochrome c oxidase subunit 6B1 | COX6B1 | COX6B1 or COX6B | 19q13.12 | [25] |
| Cytochrome c oxidase subunit 6B2 | COX6B2 | COX6B2 or COX6B | 19q13.42 | [25] |
| Cytochrome c oxidase subunit 6C | COX6C | COX6C | 8q22.2 | [25] |
| Cytochrome c oxidase subunit 7A1 | COX7A1 | COX7A | 19q13.12 | [26] |
| Cytochrome c oxidase subunit 7A2 | COX7A2 | COX7A2 or COX7A | 6q14.1 | [26] |
| Cytochrome c oxidase subunit 7B | COX7B | COX7B | Xq21.1 | [25] |
| Cytochrome c oxidase subunit 7B2 | COX7B2 | COX7B2 or COX7B | 4p12 | [25] |
| Cytochrome c oxidase subunit 7C | COX7C | COX7C | 5q14.3 | [25] |
| Cytochrome c oxidase subunit 8A | COX8A | COX8A or COX8 | 11q13.1 | [26] |
| Cytochrome c oxidase subunit 8C | COX8C | COX8C or COX8 | 14q32.12 | [26] |
| Cytochrome c oxidase subunit fa4 | NDUFA4 | NDUFA4 or COXFA4 | 7p21.3 | [28] |
| Material | Database | Samples | Reference |
|---|---|---|---|
| mtDNA | The Cancer Mitochondria Atlas | 216 | [32] |
| nDNA | Breast Invasive Carcinoma (British Columbia, Nature 2012) | 65 | [33] |
| Breast Invasive Carcinoma (Broad, Nature 2012) | 103 | [34] | |
| Breast Invasive Carcinoma (Sanger, Nature 2012) | 100 | [35] | |
| Metastatic Breast Cancer (INSERM, PLoS Med 2016) | 216 | [36] | |
| Breast Invasive Carcinoma (TCGA, PanCancer Atlas) | 1084 | [37,38,39,40,41,42,43,44,45,46,47] | |
| The Metastatic Breast Cancer Project (Provisional, December 2021) | 301 | [48,49,50] | |
| Proteogenomic landscape of breast cancer (CPTAC, Cell 2020) | 122 | [51] |
| Age by Sex | |||
|---|---|---|---|
| Median | Minimum | Maximum | |
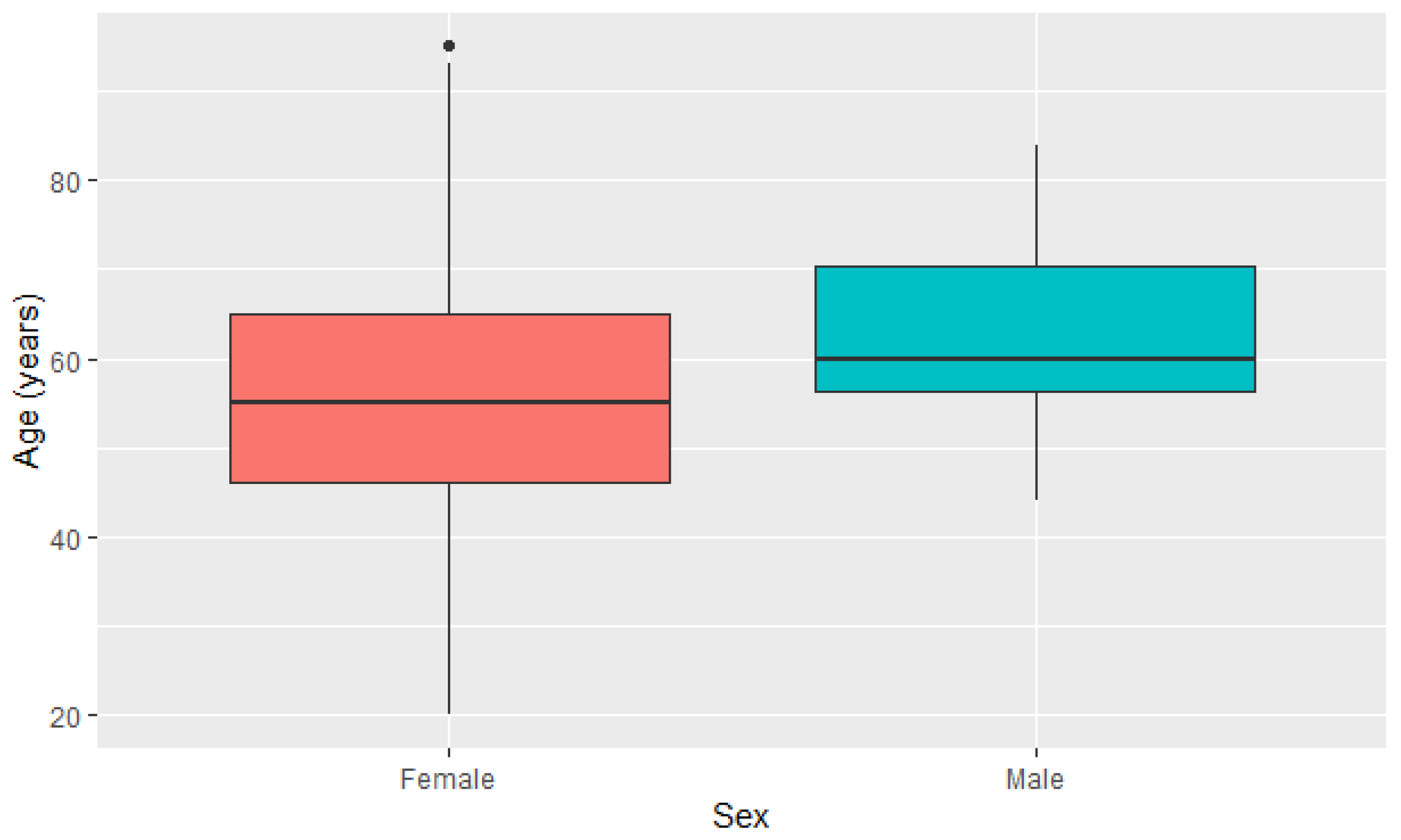
| Female (n = 1701) | 55.9 ± 13.694 | 20.0 | 95.0 |
| Male (n = 278) | 62.6 ± 12.391 | 44.0 | 84.0 |
| Frequency Distribution | |||
| Alteration | Sex | Frequency (%) | χ2 |
| With | Female | 28 (75.7) | p = 0.654 |
| Male | 0 (0.0) | ||
| ND | 9 (24.3) | ||
| Without | Female | 1673 (85.6) | |
| Male | 12 (0.6) | ||
| ND | 269 (13.8) | ||
| Alteration | Molecular Subtype | Frequency (%) | χ2 |
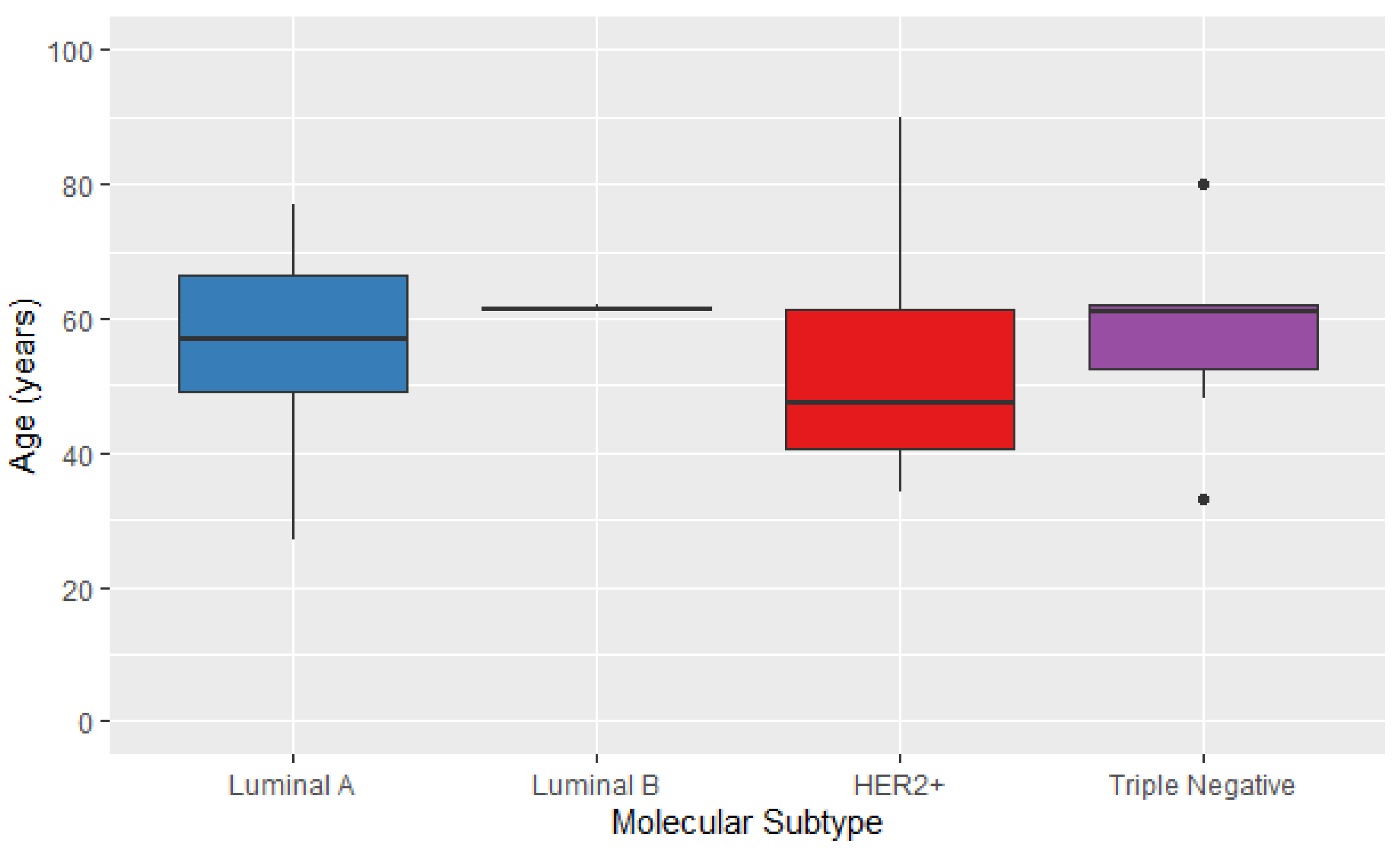
| With | Normal-like | 0 (0.0) | p = 0.108 |
| Luminal A | 14 (37.8) | ||
| Luminal B | 2 (5.4) | ||
| HER2 | 4 (10.8) | ||
| Basal-like | 8 (21.6) | ||
| ND | 9 (24.4) | ||
| Without | Normal-like | 75 (3.8) | |
| Luminal A | 804 (41.2) | ||
| Luminal B | 337 (17.3) | ||
| HER2 | 114 (5.8) | ||
| Basal-like | 288 (14.7) | ||
| ND | 336 (17.2) | ||
| Alteration | Cancer staging | Frequency (%) | χ2 |
| With | I | 5 (13.5) | p = 0.714 |
| II | 15 (40.5) | ||
| III | 5 (13.5) | ||
| IV | 2 (5.4) | ||
| ND | 10 (27.1) | ||
| Without | I | 235 (12.1) | |
| II | 840 (42.9) | ||
| III | 448 (22.9) | ||
| IV | 87 (4.5) | ||
| ND | 344 (17.6) | ||
| Alteration | Metastasis staging | Frequency (%) | χ2 |
| With | MX | 2 (5.4) | p = 0.535 |
| M0 | 18 (48.7) | ||
| CM0 (I+) | 0 (0.0) | ||
| M1 | 12 (32.4) | ||
| ND | 5 (13.5) | ||
| Without | MX | 160 (8.2) | |
| M0 | 1013 (51.8) | ||
| CM0 (I+) | 6 (0.4) | ||
| M1 | 427 (21.8) | ||
| ND | 348 (17.8) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Oliveira, R.C.; dos Reis, S.P.; Cavalcante, G.C. Mutations in Structural Genes of the Mitochondrial Complex IV May Influence Breast Cancer. Genes 2023, 14, 1465. https://doi.org/10.3390/genes14071465
de Oliveira RC, dos Reis SP, Cavalcante GC. Mutations in Structural Genes of the Mitochondrial Complex IV May Influence Breast Cancer. Genes. 2023; 14(7):1465. https://doi.org/10.3390/genes14071465
Chicago/Turabian Stylede Oliveira, Ricardo Cunha, Sávio Pinho dos Reis, and Giovanna C. Cavalcante. 2023. "Mutations in Structural Genes of the Mitochondrial Complex IV May Influence Breast Cancer" Genes 14, no. 7: 1465. https://doi.org/10.3390/genes14071465