
Human Population Density Influences Genetic Diversity of Two Rattus Species Worldwide: A Macrogenetic Approach

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Eligibility Criteria
2.2.1. Inclusion Criteria
2.2.2. Exclusion Criteria
2.3. Literature Search and Selection of Articles
2.4. Data Extraction
2.5. Risk of Bias across Studies
2.6. Definition of Predictors
3. Results
3.1. Systematic Review
3.2. Metrics of Genetic Diversity
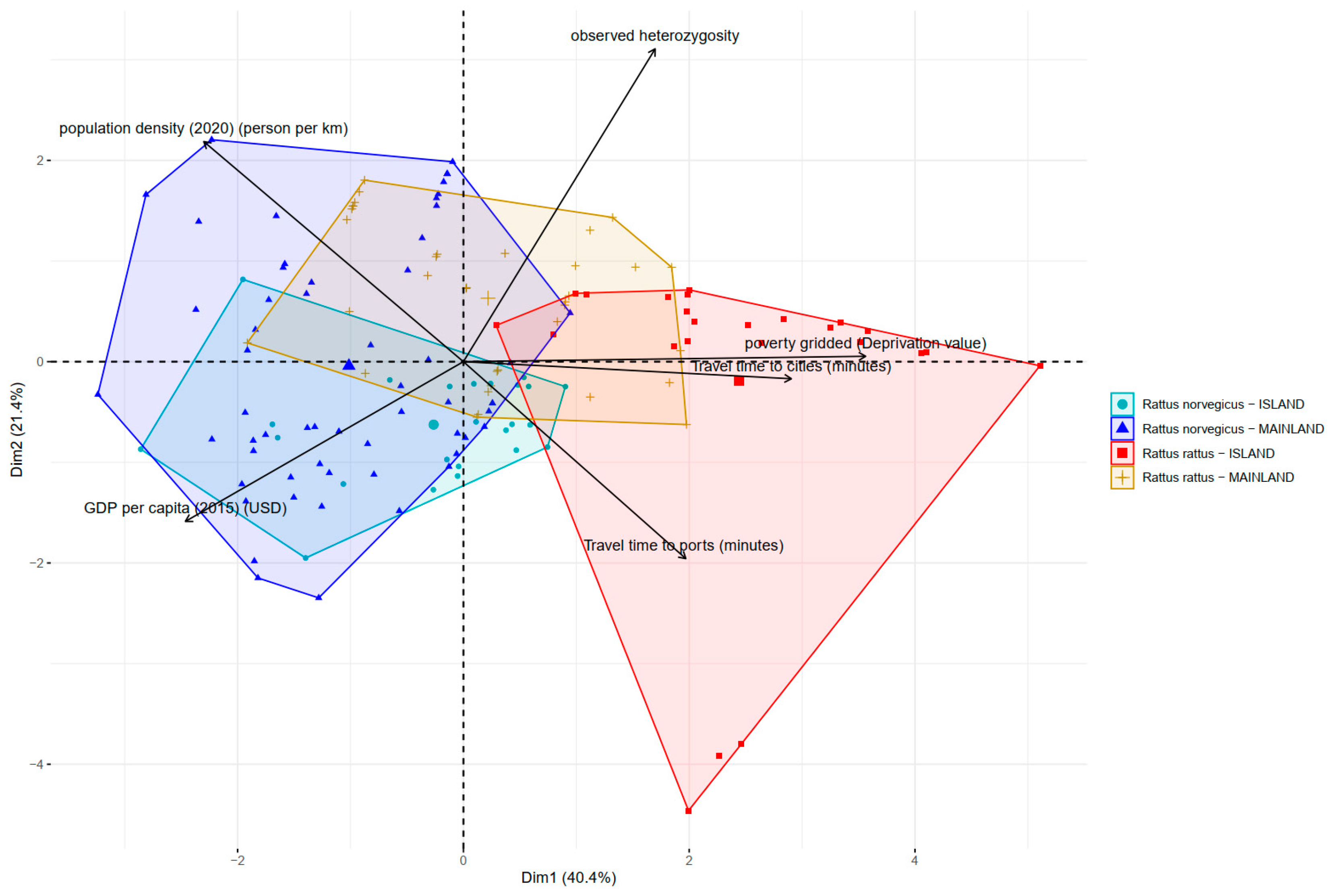
3.3. Predictors of Genetic Diversity
4. Discussion
5. Conclusions and Recommendations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Bank. Urban Development. Available online: https://www.worldbank.org/en/topic/urbandevelopment/overview (accessed on 3 May 2023).
- Cohen, B. Urbanization in developing countries: Current trends, future projections, and key challenges for sustainability. Technol. Soc. 2006, 28, 63–80. [Google Scholar] [CrossRef]
- Fusco, N.; Carlen, E.; Munshi-South, J. Urban landscape genetics: Are biologists keeping up with the pace of urbanization? Curr. Landsc. Ecol. Rep. 2021, 6, 35–45. [Google Scholar] [CrossRef]
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef] [PubMed]
- Banks, P.B.; Smith, H.M. The ecological impacts of commensal species: Black rats, Rattus rattus, at the urban–bushland interface. Wildl. Res. 2015, 42, 86–97. [Google Scholar] [CrossRef]
- Combs, M.; Byers, K.A.; Ghersi, B.M.; Blum, M.J.; Caccone, A.; Costa, F.; Himsworth, C.G.; Richardson, J.L.; Munshi-South, J. Urban rat races: Spatial population genomics of brown rats (Rattus norvegicus) compared across multiple cities. Proc. R. Soc. B Biol. Sci. 2018, 285, 20180245. [Google Scholar] [CrossRef] [Green Version]
- Doherty, T.S.; Glen, A.S.; Nimmo, D.G.; Ritchie, E.G.; Dickman, C.R. Invasive predators and global biodiversity loss. Proc. Natl. Acad. Sci. USA 2016, 113, 11261–11265. [Google Scholar] [CrossRef]
- Godwin, J.; Serr, M.; Barnhill-Dilling, S.K.; Blondel, D.V.; Brown, P.R.; Campbell, K.; Delborne, J.; Lloyd, A.L.; Oh, K.P.; Prowse, T.A.A.; et al. Rodent gene drives for conservation: Opportunities and data needs. Proc. R. Soc. B Biol. Sci. 2019, 286, 20191606. [Google Scholar] [CrossRef] [Green Version]
- Blasdell, K.R.; Morand, S.; Laurance, S.G.W.; Doggett, S.L.; Hahs, A.; Trinh, K.; Perera, D.; Firth, C. Rats and the city: Implications of urbanization on zoonotic disease risk in Southeast Asia. Proc. Natl. Acad. Sci. USA 2022, 119, e2112341119. [Google Scholar] [CrossRef]
- Morand, S.; Bordes, F.; Chen, H.W.; Claude, J.; Cosson, J.F.; Galan, M.; Czirjak, G.A.; Greenwood, A.D.; Latinne, A.; Michaux, J.; et al. Global parasite and Rattus rodent invasions: The consequences for rodent-borne diseases. Integr. Zool. 2015, 10, 409–423. [Google Scholar] [CrossRef]
- Fantle-Lepczyk, J.E.; Haubrock, P.J.; Kramer, A.M.; Cuthbert, R.N.; Turbelin, A.J.; Crystal-Ornelas, R.; Diagne, C.; Courchamp, F. Economic costs of biological invasions in the United States. Sci. Total Environ. 2022, 806, 151318. [Google Scholar] [CrossRef]
- Haubrock, P.J.; Turbelin, A.J.; Cuthbert, R.N.; Novoa, A.; Taylor, N.G.; Angulo, E.; Ballesteros-Mejia, L.; Bodey, T.W.; Capinha, C.; Diagne, C.; et al. Economic costs of invasive alien species across Europe. NeoBiota 2021, 67, 153–190. [Google Scholar] [CrossRef]
- Rissler, L.J. Union of phylogeography and landscape genetics. Proc. Natl. Acad. Sci. USA 2016, 113, 8079–8086. [Google Scholar] [CrossRef]
- Leigh, D.M.; van Rees, C.B.; Millette, K.L.; Breed, M.F.; Schmidt, C.; Bertola, L.D.; Hand, B.K.; Hunter, M.E.; Jensen, E.L.; Kershaw, F.; et al. Opportunities and challenges of macrogenetic studies. Nat. Rev. Genet. 2021, 22, 791–807. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Stewart, L.; Shekelle, P. All in the family: Systematic reviews, rapid reviews, scoping reviews, realist reviews, and more. Syst. Rev. 2015, 4, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haddaway, N.R.; Macura, B.; Whaley, P.; Pullin, A.S. ROSES RepOrting standards for Systematic Evidence Syntheses: Pro forma, flow-diagram and descriptive summary of the plan and conduct of environmental systematic reviews and systematic maps. Environ. Evid. 2018, 7, 7. [Google Scholar] [CrossRef]
- Sutherland, W.J.; Wordley, C.F. A fresh approach to evidence synthesis. Nature 2018, 558, 364–366. [Google Scholar] [CrossRef] [Green Version]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [Green Version]
- Combs, M.; Puckett, E.E.; Richardson, J.; Mims, D.; Munshi-South, J. Spatial population genomics of the brown rat (Rattus norvegicus) in New York City. Mol. Ecol. 2018, 27, 83–98. [Google Scholar] [CrossRef]
- Gardner-Santana, L.C.; Norris, D.E.; Fornadel, C.M.; Hinson, E.R.; Klein, S.L.; Glass, G.E. Commensal ecology, urban landscapes, and their influence on the genetic characteristics of city-dwelling Norway rats (Rattus norvegicus). Mol. Ecol. 2009, 18, 2766–2778. [Google Scholar] [CrossRef] [Green Version]
- Kajdacsi, B.; Costa, F.; Hyseni, C.; Porter, F.; Brown, J.; Rodrigues, G.; Farias, H.; Reis, M.G.; Childs, J.E.; Ko, A.I.; et al. Urban population genetics of slum-dwelling rats (Rattus norvegicus) in Salvador, Brazil. Mol. Ecol. 2013, 22, 5056–5070. [Google Scholar] [CrossRef]
- Kummu, M.; Taka, M.; Guillaume, J.H.A. Gridded global datasets for Gross Domestic Product and Human Development Index over 1990–2015. Sci. Data 2018, 5, 180004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Center for International Earth Science Information Network—CIESIN—Columbia University. Gridded Population of the World, Version 4 (GPWv4): Population Density, Revision 11. 2018. Available online: https://sedac.ciesin.columbia.edu/data/collection/gpw-v4/documentation (accessed on 18 May 2023).
- Center for International Earth Science Information Network—CIESIN—Columbia University. Global Gridded Relative Deprivation Index (GRDI), Version 1. 2022. Available online: https://sedac.ciesin.columbia.edu/data/set/povmap-grdi-v1 (accessed on 18 May 2023).
- Nelson, A.; Weiss, D.J.; van Etten, J.; Cattaneo, A.; McMenomy, T.S.; Koo, J. A suite of global accessibility indicators. Sci. Data 2019, 6, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillaumot, C.; Artois, J.; Saucde, T.; Demoustier, L.; Moreau, C.; Elaume, M.; Agera, A.; Danis, B. Broad-scale species distribution models applied to data-poor areas. Prog. Oceanogr. 2019, 175, 198–207. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, C.; Hoban, S.; Jetz, W. Conservation Macrogenetics. 2023. Available online: https://ecoevorxiv.org/repository/view/5317/ (accessed on 2 May 2023).
- Lack, J.B.; Hamilton, M.J.; Braun, J.K.; Mares, M.A.; van Den Bussche, R.A. Comparative phylogeography of invasive Rattus rattus and Rattus norvegicus in the U.S. reveals distinct colonization histories and dispersal. Biol. Invasions 2013, 15, 1067. [Google Scholar] [CrossRef]
- Brouat, C.; Rahelinirina, S.; Loiseau, A.; Rahalison, L.; Rajerison, M.; Laffly, D.; Handschumacher, P.; Duplantier, J.M. Plague circulation and population genetics of the reservoir Rattus rattus: The influence of topographic relief on the distribution of the disease within the Madagascan focus. PLoS Negl. Trop. Dis. 2013, 7, e2266. [Google Scholar] [CrossRef] [Green Version]
- Mangombi, J.B.; Brouat, C.; Loiseau, A.; Banga, O.; Leroy, E.M.; Bourgarel, M.; Duplantier, J.M. Urban population genetics of the invasive black rats in Franceville, Gabon. J. Zool. 2016, 299, 1–8. [Google Scholar] [CrossRef]
- Berthier, K.; Garba, M.; Leblois, R.; Navascues, M.; Tatard, C.; Gauthier, P.; Gagare, S.; Piry, S.; Brouat, C.; Dalecky, A.; et al. Black rat invasion of inland Sahel: Insights from interviews and population genetics in south-western Niger. Biol. J. Linn. Soc. 2016, 119, 748–765. [Google Scholar] [CrossRef] [Green Version]
- Millette, K.L.; Fugère, V.; Debyser, C.; Greiner, A.; Chain, F.J.J.; Gonzalez, A. No consistent effects of humans on animal genetic diversity worldwide. Ecol. Lett. 2020, 23, 55–67. [Google Scholar] [CrossRef]
- Johnson, M.T.; Munshi-South, J. Evolution of life in urban environments. Science 2017, 358, eaam8327. [Google Scholar] [CrossRef] [Green Version]
- Tucker, M.A.; Böhning-Gaese, K.; Fagan, W.F.; Fryxell, J.M.; van Moorter, B.; Alberts, S.C.; Ali, A.H.; Allen, A.M.; Attias, N.; Avgar, T.; et al. Moving in the Anthropocene: Global reductions in terrestrial mammalian movements. Science 2018, 359, 466–469. [Google Scholar] [CrossRef] [Green Version]
- Baig, M.; Sameera, F.; Atkulwar, A.; Searle, J. Genomic analysis reveals subdivision of black rats (Rattus rattus) in India, origin of the worldwide species spread. Genes 2022, 13, 267. [Google Scholar] [CrossRef] [PubMed]
- Konečný, A.; Estoup, A.; Duplantier, J.-M.; Bryja, J.; Bâ, K.; Galan, M.; Tatard, C.; Cosson, J.-F. Invasion genetics of the introduced black rat (Rattus rattus) in Senegal, West Africa. Mol. Ecol. 2013, 22, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Puckett, E.E.; Park, J.; Combs, M.; Blum, M.J.; Bryant, J.E.; Caccone, A.; Costa, F.; Deinum, E.E.; Esther, A.; Himsworth, C.G.; et al. Global population divergence and admixture of the brown rat (Rattus norvegicus). Proc. R. Soc. B Biol. Sci. 2016, 283, 20161762. [Google Scholar] [CrossRef]
- Haniza, M.Z.H.; Adams, S.; Jones, E.P.; MacNicoll, A.; Mallon, E.B.; Smith, R.H.; Lambert, M.S. Large-scale structure of brown rat (Rattus norvegicus) populations in England: Effects on rodenticide resistance. PeerJ 2015, 3, e1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conroy, C.J.; Rowe, K.C.; Rowe, K.M.C.; Kamath, P.L.; Aplin, K.P.; Hui, L.; James, D.K.; Moritz, C.; Patton, J.L. Cryptic genetic diversity in Rattus of the San Francisco Bay region, California. Biol. Invasions 2013, 15, 741–758. [Google Scholar] [CrossRef]
- Cheylan, G. Les mammifères des îles de Provence et de Méditerranée occidentale: Une exemple du peuplement insulaire non équilibré? Rev. D’ecologie 1984, 39, 37–54. [Google Scholar] [CrossRef]
- Sjodin, B.M.F.; Irvine, R.L.; Ford, A.T.; Howald, G.R.; Russello, M.A. Rattus population genomics across the Haida Gwaii archipelago provides a framework for guiding invasive species management. Evol. Appl. 2020, 13, 889–904. [Google Scholar] [CrossRef] [Green Version]
- Badou, S.; Gauthier, P.; Houéménou, G.; Loiseau, A.; Dossou, J.; Etougbétché, J.; Houéménou, H.; Agbangla, C.; Brouat, C.; Dobigny, G. Population genetic structure of blacks rats in an urban environment: A case study in Cotonou, Benin. Hystrix It. J. Mammal 2021, 32, 1825–5272. [Google Scholar] [CrossRef]
- Varudkar, A.; Ramakrishnan, U. Commensalism facilitates gene flow in mountains: A comparison between two Rattus species. Heredity 2015, 115, 253–261. [Google Scholar] [CrossRef] [Green Version]
- Willows-Munro, S.; Dowler, R.; Jarcho, M.R.; Phillips, R.B.; Snell, H.L.; Wilbert, T.R.; Edwards, C.W. Cryptic diversity in black rats Rattus rattus of the Galápagos islands, Ecuador. Ecol. Evol. 2016, 6, 3721–3733. [Google Scholar] [CrossRef] [Green Version]
- Chacón-Vargas, K.; García-Merchán, V.H.; Sanín, M.J. From keystone species to conservation: Conservation genetics of wax palm Ceroxylon quindiuense in the largest wild populations of Colombia and selected neighboring ex situ plant collections. Biodivers. Conserv. 2020, 29, 283–302. [Google Scholar] [CrossRef]
- García-Merchán, V.H.; Arenas-Calle, C.; Sosa-Arango, C.C.; Castaño-Castro, P.; Chacón-Vargas, K. Conservation genetics in the neotropical carnivorous mammals: A systematic review. In Molecular Ecology and Conservation Genetics of Neotropical Mammals; Nardelli, M., Túnez, J.I., Eds.; Springer Nature: Basel, Switzerland, 2021; pp. 277–295. [Google Scholar]
- MacroPopGen. MacroPopGen Database: Geo-Referenced Population-Specific Microsatellite Data across the American Continents. Available online: https://figshare.com/articles/dataset/MacroPopGen_Database_Geo-referenced_population-specific_microsatellite_data_across_the_American_continents/7207514/1 (accessed on 14 May 2023).
- Abdelkrim, J.; Pascal, M.; Samadi, S. Island colonization and founder effects: The invasion of the Guadeloupe islands by ship rats (Rattus rattus). Mol. Ecol. 2005, 14, 2923–2931. [Google Scholar] [CrossRef] [PubMed]
- Abdelkrim, J.; Pascal, M.; Calmet, C.; Samadi, S. Importance of Assessing Population Genetic Structure before Eradication of Invasive Species: Examples from Insular Norway Rat Populations. Conserv. Biol. 2005, 19, 1509–1518. [Google Scholar] [CrossRef]
- Abdelkrim, J.; Pascal, M.; Samadi, S. Genetic structure and functioning of alien ship rat populations from a Corsican micro-insular complex. Biol. Invasions 2008, 11, 473–482. [Google Scholar] [CrossRef]
- Abdelkrim, J.; Byrom, A.E.; Gemmell, N.J. Fine-scale genetic structure of mainland invasive Rattus rattus populations: Implications for restoration of forested conservation areas in New Zealand. Conserv. Genet. 2010, 11, 1953–1964. [Google Scholar] [CrossRef]
- Calmet, C.; Pascal, M.; Samadi, S. Is it worth eradicating the invasive pest Rattus norvegicus from Molène archipelago? Genetic structure as a decision-making tool. Biodivers. Conserv. 2001, 10, 911–928. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, L.; Teng, H.; Shi, C.; Liu, Q.; Zhang, J.; Zhang, Y. Population genomics reveal rapid genetic differentiation in a recently invasive population of Rattus norvegicus. Front. Zool. 2021, 18, 1–10. [Google Scholar] [CrossRef]
- Costa, F.; Richardson, J.L.; Dion, K.; Mariani, C.; Pertile, A.C.; Burak, M.K.; Childs, J.E.; Ko, A.I.; Caccone, A. Multiple Paternity in the Norway Rat, Rattus norvegicus, from Urban Slums in Salvador, Brazil. J. Hered. 2016, 107, 181–186. [Google Scholar] [CrossRef]
- Cramer, D.V.; Chakravarti, A.; Arenas, O.; Humprieres, J.; Mowery, P.A. Genetic Diversity within and between Natural Populations of Rattus norvegicus. J. Hered. 1988, 79, 319–324. [Google Scholar] [CrossRef]
- Desvars-Larrive, A.; Pascal, M.; Gasqui, P.; Cosson, J.-F.; Benoît, E.; Lattard, V.; Crespin, L.; Lorvelec, O.; Pisanu, B.; Teynié, A.; et al. Population genetics, community of parasites, and resistance to rodenticides in an urban brown rat (Rattus norvegicus) population. PLoS ONE 2017, 12, e0184015. [Google Scholar] [CrossRef] [Green Version]
- Gilabert, A.; Loiseau, A.; Duplantier, J.-M.; Rahelinirina, S.; Rahalison, L.; Chanteau, S.; Brouat, C. Genetic structure of black rat populations in a rural plague focus in Madagascar. Can. J. Zool. 2007, 85, 965–972. [Google Scholar] [CrossRef]
- Villafañe, I.E.G.; Guzmán, N.; Ortiz, N.; Gonzalez-Ittig, R.; León, V.A.; Busch, M. Connectivity among Norway rat subpopulations (Rattus norvegicus) at poultry farms in Exaltación de la Cruz, Buenos Aires, Argentina. Mastozoología Neotrop. 2019, 26, 359–367. [Google Scholar] [CrossRef]
- Natali, C.; Ciofi, C.; Sposimo, P.; Fratini, S. Population genetic analysis of invasive black rats: Defining eradication units in the Tuscan Archipelago National Park. Hystrix 2018, 29, 67–73. [Google Scholar] [CrossRef]
- Jing, M.; Chen, Y.; Yao, K.; Wang, Y.; Huang, L. Comparative phylogeography of two commensal rat species (Rattus tanezumi and Rattus norvegicus) in China: Insights from mitochondrial DNA, microsatellite, and 2b-RAD data. Ecol. Evol. 2022, 12, e9409. [Google Scholar] [CrossRef]
- Piertney, S.B.; Black, A.; Watt, L.; Christie, D.; Poncet, S.; Collins, M.A. Resolving patterns of population genetic and phylogeographic structure to inform control and eradication initiatives for brown rats Rattus norvegicus on South Georgia. J. Appl. Ecol. 2016, 53, 332–339. [Google Scholar] [CrossRef]
- Puckett, E.E.; Micci-Smith, O.; Munshi-South, J. Genomic analyses identify multiple Asian origins and deeply diverged mitochondrial clades in inbred brown rats ( Rattus norvegicus ). Evol. Appl. 2017, 11, 718–726. [Google Scholar] [CrossRef]
- Puckett, E.E.; Magnussen, E.; Khlyap, L.A.; Strand, T.M.; Lundkvist, Å.; Munshi-South, J. Genomic analyses reveal three independent introductions of the invasive brown rat (Rattus norvegicus) to the Faroe Islands. Heredity 2019, 124, 15–27. [Google Scholar] [CrossRef]
- Tabak, M.A.; Poncet, S.; Passfield, K.; Carling, M.D.; del Rio, C.M. The relationship between distance and genetic similarity among invasive rat populations in the Falkland Islands. Conserv. Genet. 2014, 16, 125–135. [Google Scholar] [CrossRef]
- Teng, H.; Zhang, Y.; Shi, C.; Mao, F.; Cai, W.; Lu, L.; Zhao, F.; Sun, Z.; Zhang, J. Population Genomics Reveals Speciation and Introgression between Brown Norway Rats and Their Sibling Species. Mol. Biol. Evol. 2017, 34, 2214–2228. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predictor | R. norvegicus | R. rattus |
|---|---|---|
| GDP_per_capita_2015 | −0.064 (0.567) | −0.108 (0.417) |
| gpw_v4_population_density_2020 | 0.385 (0) | −0.236 (0.072) |
| povmap.grdi.v1 | 0.162 (0.143) | 0.3 (0.021) |
| travel_time_to_cities_12_MOD | −0.004 (0.97) | 0.286 (0.028) |
| travel_time_to_ports_5_MOD | −0.058 (0.604) | −0.588 (0) |
| Predictor | R. norvegicus | R. rattus |
|---|---|---|
| GDP_per_capita_2015 | 0.743 | 0.697 |
| gpw_v4_population_density_2020 | 1 | 0.741 |
| povmap.grdi.v1 | 0.720 | 0.363 |
| travel_time_to_cities_12_MOD | 0.205 | 0.595 |
| travel_time_to_ports_5_MOD | 0.565 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sosa, C.C.; Arenas, C.; García-Merchán, V.H. Human Population Density Influences Genetic Diversity of Two Rattus Species Worldwide: A Macrogenetic Approach. Genes 2023, 14, 1442. https://doi.org/10.3390/genes14071442
Sosa CC, Arenas C, García-Merchán VH. Human Population Density Influences Genetic Diversity of Two Rattus Species Worldwide: A Macrogenetic Approach. Genes. 2023; 14(7):1442. https://doi.org/10.3390/genes14071442
Chicago/Turabian StyleSosa, Chrystian C., Catalina Arenas, and Víctor Hugo García-Merchán. 2023. "Human Population Density Influences Genetic Diversity of Two Rattus Species Worldwide: A Macrogenetic Approach" Genes 14, no. 7: 1442. https://doi.org/10.3390/genes14071442