
Insight into Antibiotic Synergy Combinations for Eliminating Colistin Heteroresistant Klebsiella pneumoniae
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain Collection and MIC Determination
2.2. Detection of Colistin Heteroresistant Strains by Population Analysis Profiling (PAP) Assays
2.3. Time–Kill Assays
2.4. Whole-Genome Sequencing and Analysis
3. Results
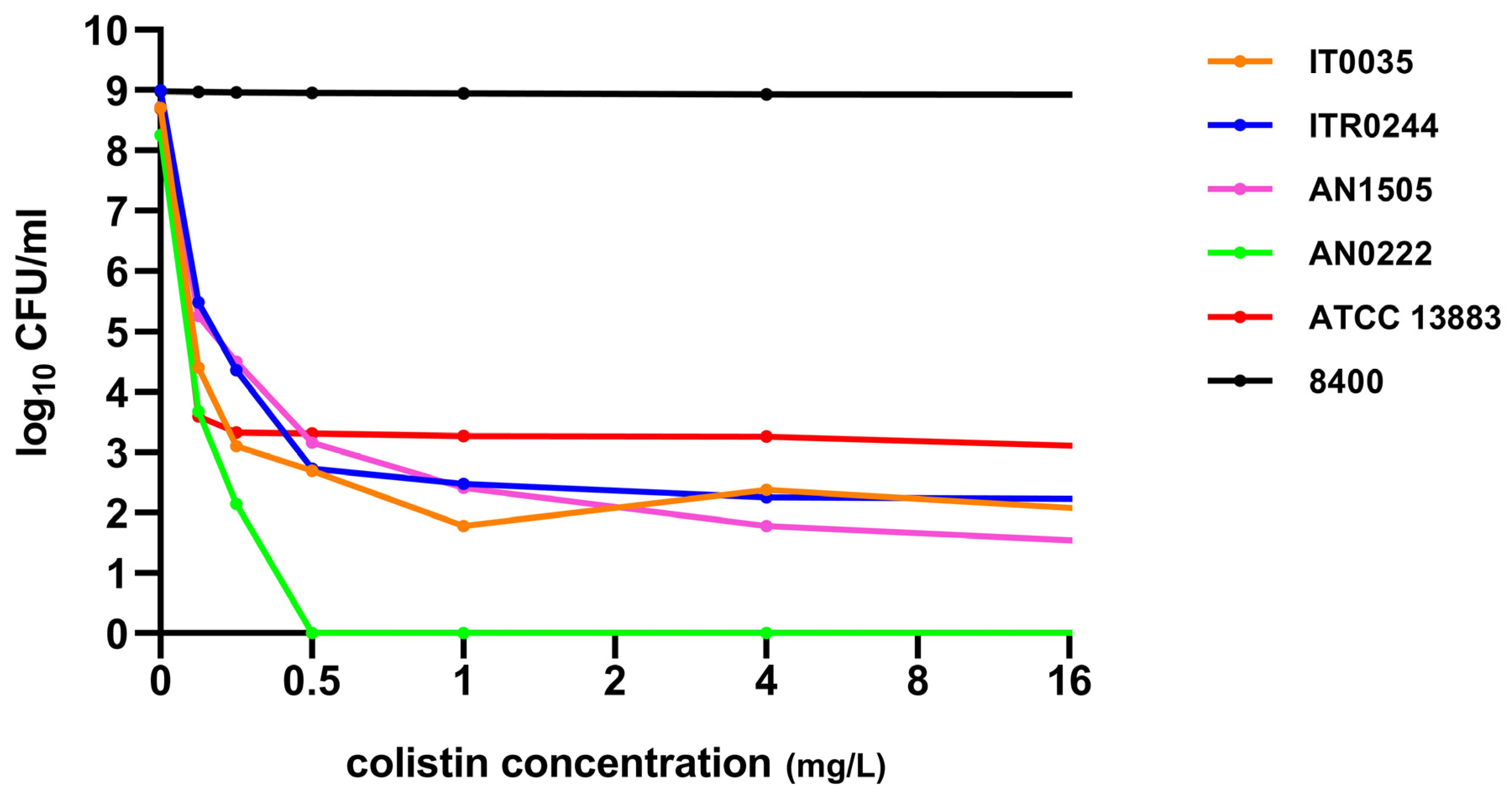
3.1. MIC Determination and Colistin Heteroresistance
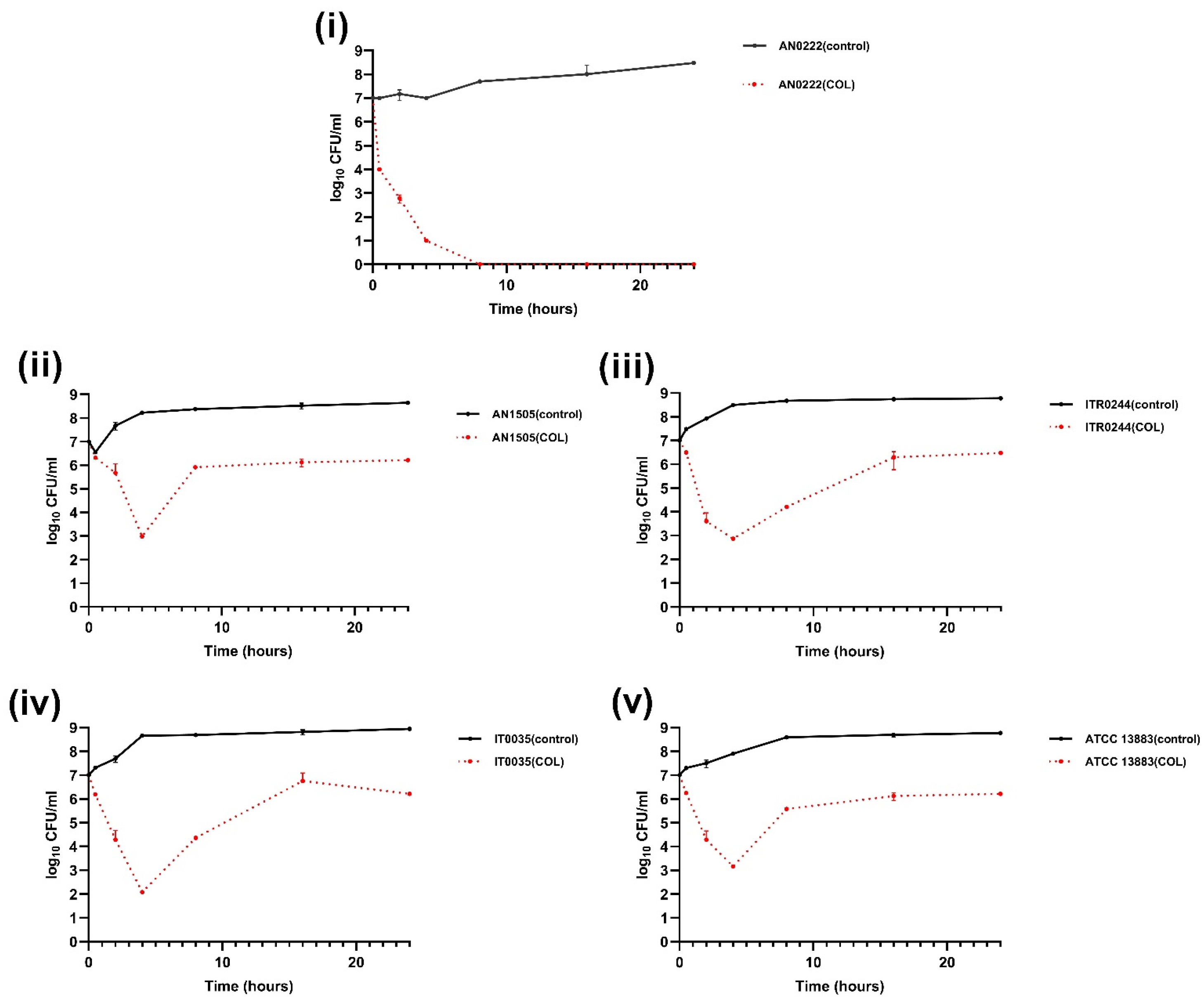
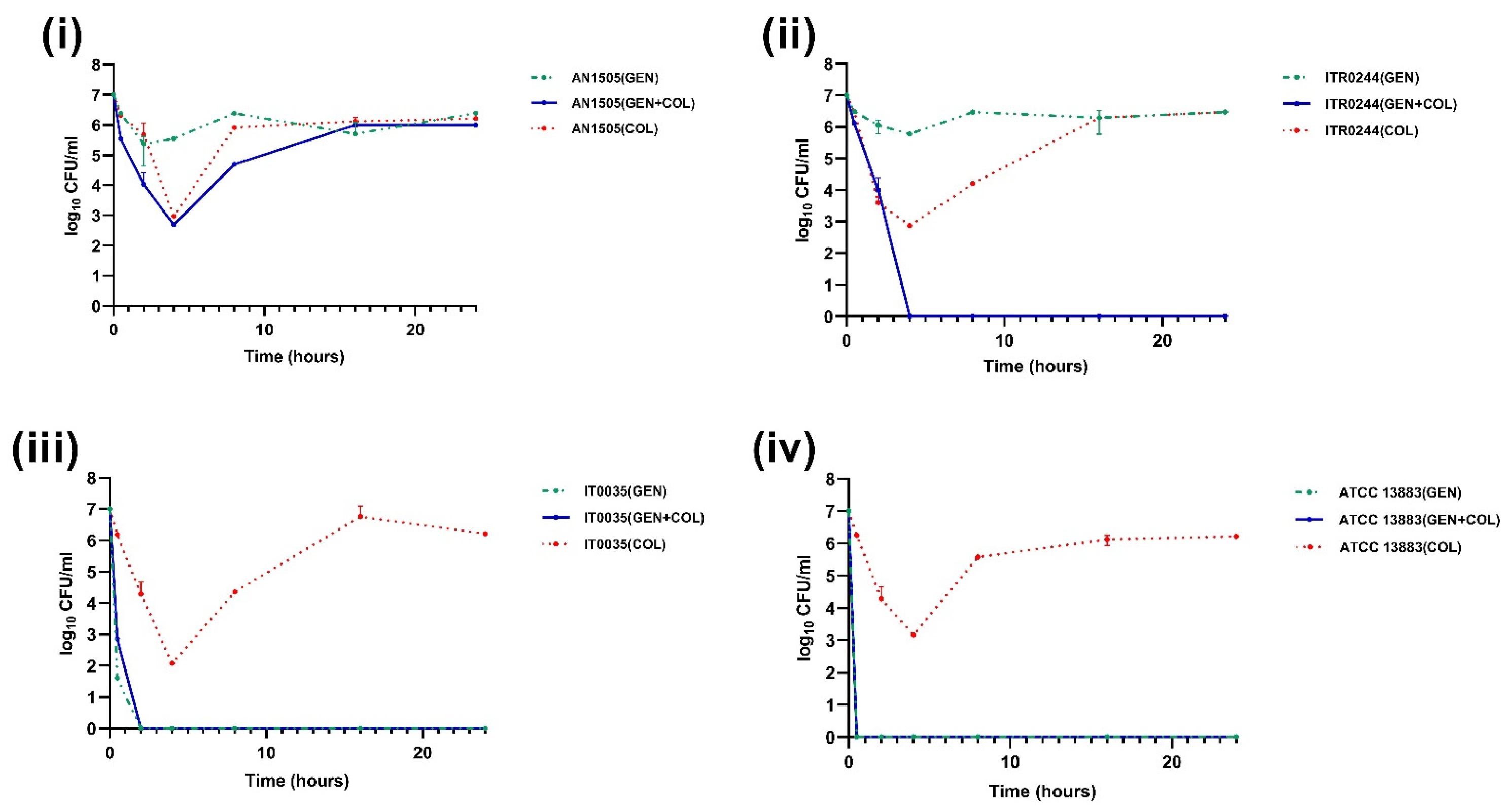
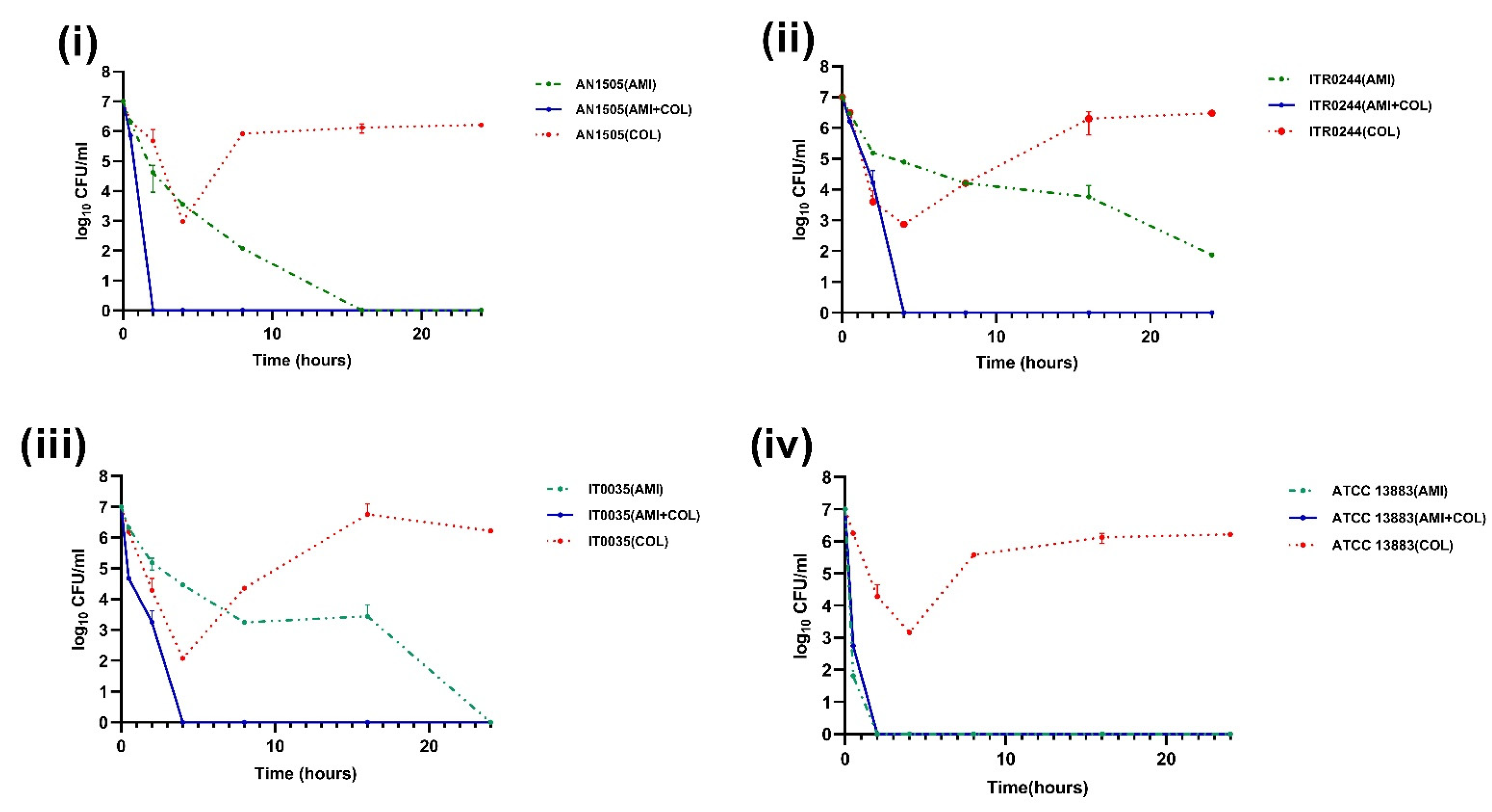
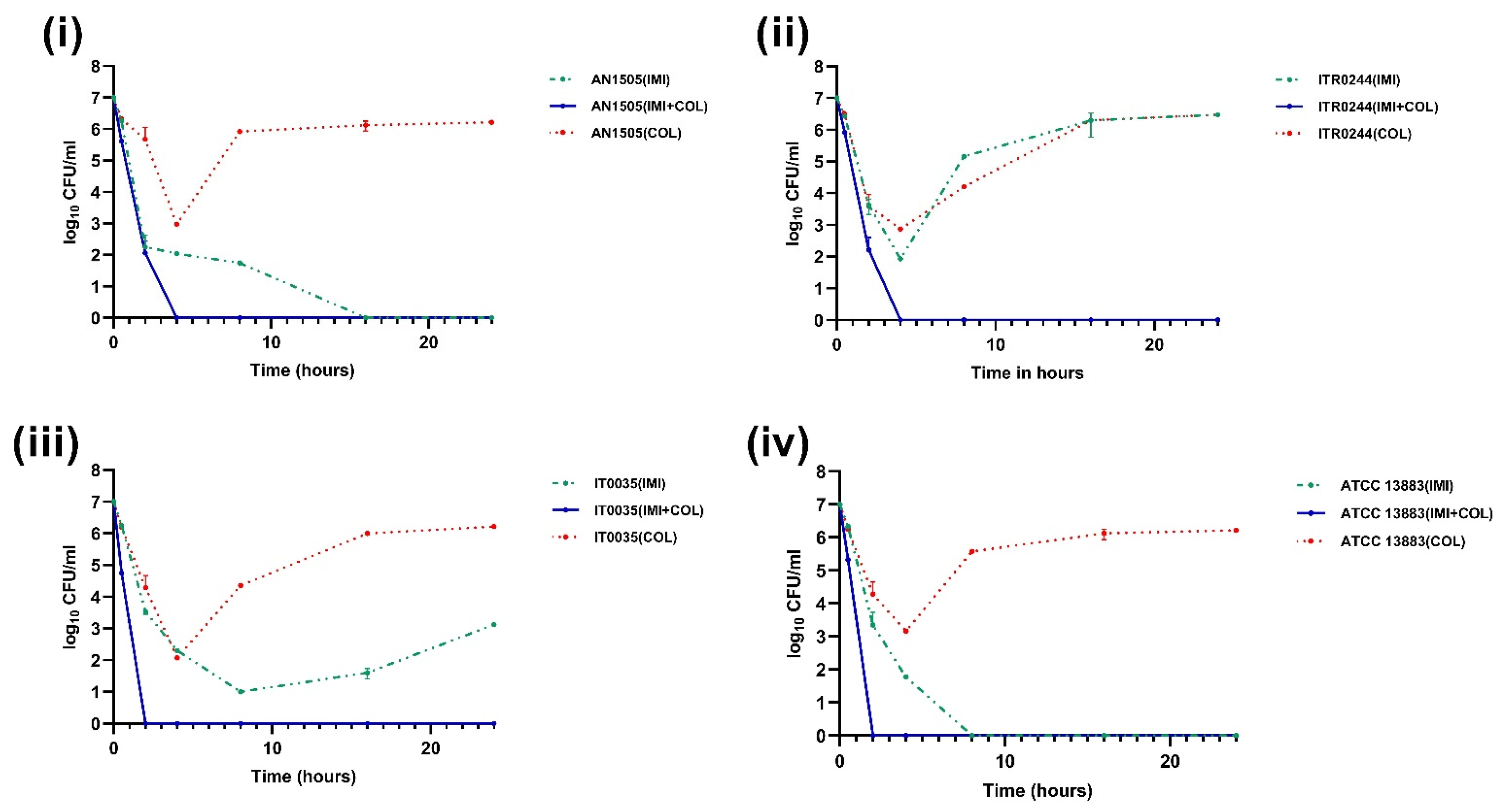
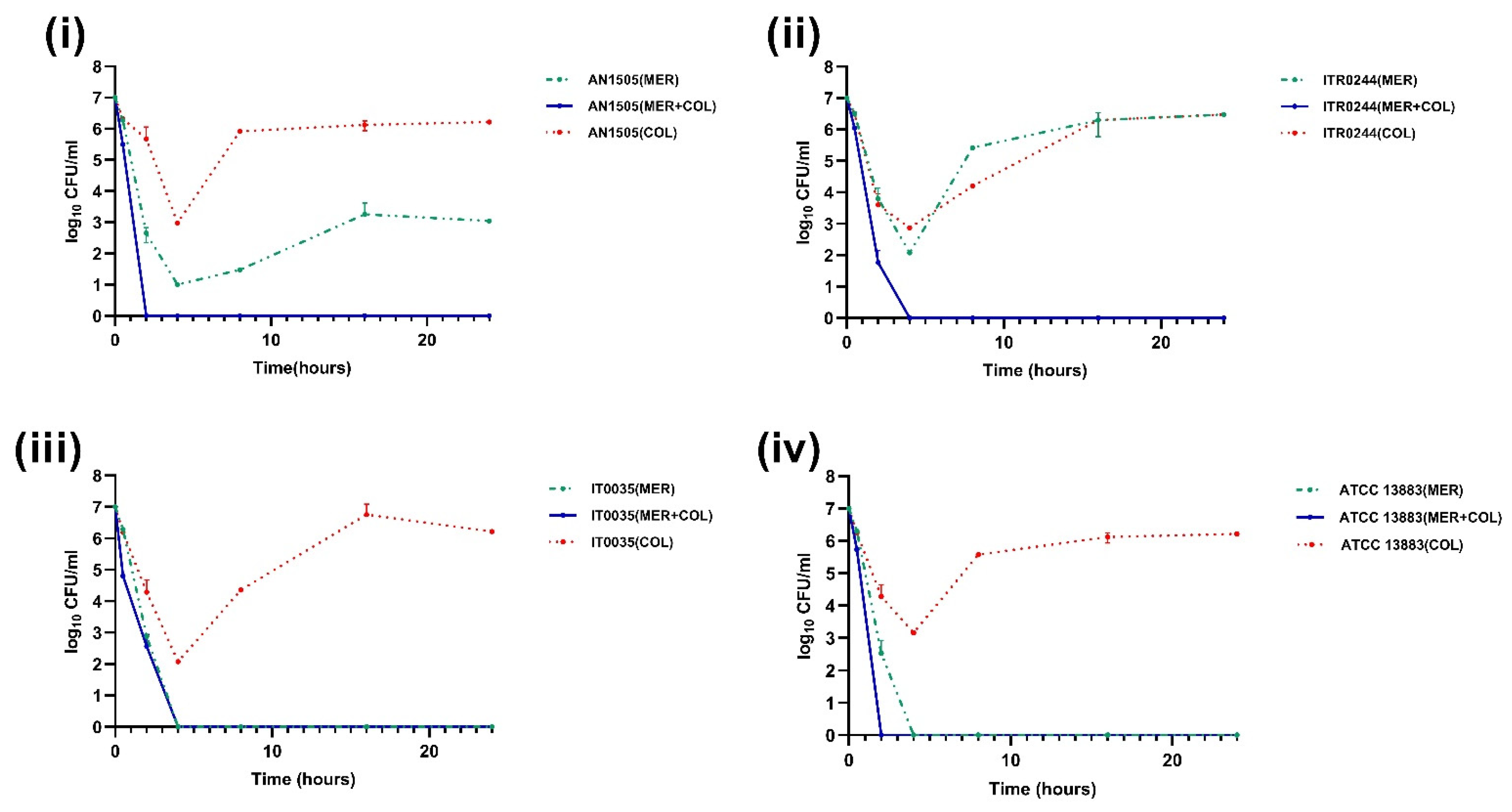
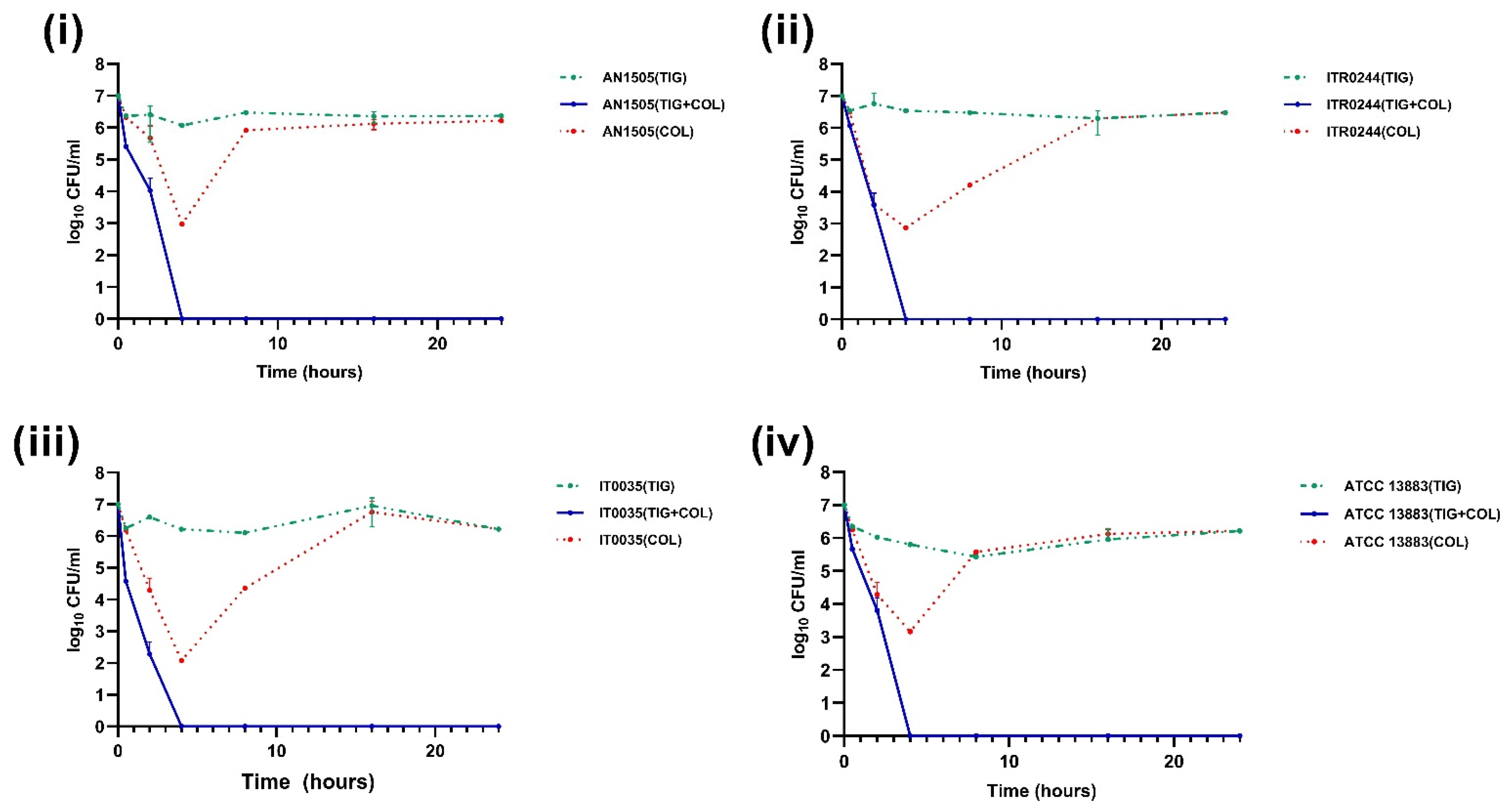
3.2. Time–Kill Curve Assays
3.3. Mutations Associated with Colistin Resistance Observed in the Heteroresistant Subpopulations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Effah, C.Y.; Sun, T.; Liu, S.; Wu, Y. Klebsiella pneumoniae: An increasing threat to public health. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 1. [Google Scholar] [CrossRef]
- Ustundag, G.; Oncel, E.K.; Sahin, A.; Keles, Y.E.; Aksay, A.K.; Ciftdogan, D.Y. Colistin Treatment for Multidrug-Resistant Gram-Negative Infections in Children: Caution Required for Nephrotoxicity. Sisli. Etfal Hastan Tip. Bul. 2022, 56, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Berglund, B. Acquired Resistance to Colistin via Chromosomal And Plasmid-Mediated Mechanisms in Klebsiella pneumoniae. Infect. Microbes Dis. 2019, 1, 10–19. [Google Scholar] [CrossRef]
- El-Sayed Ahmed, M.A.E.G.; Zhong, L.-L.; Shen, C.; Yang, Y.; Doi, Y.; Tian, G.-B. Colistin and its role in the Era of antibiotic resistance: An extended review (2000–2019). Emerg. Microbes Infect. 2020, 9, 868–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef] [Green Version]
- Band, V.I.; Weiss, D.S. Heteroresistance: A cause of unexplained antibiotic treatment failure? PLoS Pathog. 2019, 15, e1007726. [Google Scholar] [CrossRef] [Green Version]
- Jeannot, K.; Bolard, A.; Plésiat, P. Resistance to polymyxins in Gram-negative organisms. Int. J. Antimicrob. Agents 2017, 49, 526–535. [Google Scholar] [CrossRef]
- Umair, M.; Hassan, B.; Farzana, R.; Ali, Q.; Sands, K.; Mathias, J.; Afegbua, S.; Haque, M.N.; Walsh, T.R.; Mohsin, M. International manufacturing and trade in colistin, its implications in colistin resistance and One Health global policies: A microbiological, economic, and anthropological study. Lancet Microbe 2023, 4, e264–e276. [Google Scholar] [CrossRef]
- Aghapour, Z.; Gholizadeh, P.; Ganbarov, K.; Bialvaei, A.Z.; Mahmood, S.S.; Tanomand, A.; Yousefi, M.; Asgharzadeh, M.; Yousefi, B.; Kafil, H.S. Molecular mechanisms related to colistin resistance in Enterobacteriaceae. Infect. Drug Resist. 2019, 12, 965–975. [Google Scholar] [CrossRef] [Green Version]
- Lepe, J.; Martínez-Martínez, L. Resistance mechanisms in Gram-negative bacteria. Med. Intensiv. 2022, 46, 392–402. [Google Scholar] [CrossRef]
- Fordham, S.M.E.; Mantzouratou, A.; Sheridan, E. Prevalence of insertion sequence elements in plasmids relating to mgrB gene disruption causing colistin resistance in Klebsiella pneumoniae. Microbiologyopen 2022, 11, e1262. [Google Scholar] [CrossRef]
- Cannatelli, A.; Giani, T.; D’Andrea, M.M.; Di Pilato, V.; Arena, F.; Conte, V.; Tryfinopoulou, K.; Vatopoulos, A.; Rossolini, G.M. MgrB Inactivation Is a Common Mechanism of Colistin Resistance in KPC-Producing Klebsiella pneumoniae of Clinical Origin. Antimicrob. Agents Chemother. 2014, 58, 5696–5703. [Google Scholar] [CrossRef] [Green Version]
- Andersson, D.I.; Nicoloff, H.; Hjort, K. Mechanisms and clinical relevance of bacterial heteroresistance. Nat. Rev. Genet. 2019, 17, 479–496. [Google Scholar] [CrossRef] [PubMed]
- Ezadi, F.; Ardebili, A.; Mirnejad, R. Antimicrobial Susceptibility Testing for Polymyxins: Challenges, Issues, and Recommendations. J. Clin. Microbiol. 2019, 57, e01390-18. [Google Scholar] [CrossRef] [Green Version]
- Meletis, G.; Tzampaz, E.; Sianou, E.; Tzavaras, I.; Sofianou, D. Colistin heteroresistance in carbapenemase-producing Klebsiella pneumoniae. J. Antimicrob. Chemother. 2011, 66, 946–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roch, M.; Sierra, R.; Andrey, D.O. Antibiotic heteroresistance in ESKAPE pathogens, from bench to bedside. Clin. Microbiol. Infect. 2022, 29, 320–325. [Google Scholar] [CrossRef] [PubMed]
- El-Halfawy, O.M.; Valvano, M.A. Antimicrobial Heteroresistance: An Emerging Field in Need of Clarity. Clin. Microbiol. Rev. 2015, 28, 191–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landman, D.; Salamera, J.; Quale, J. Irreproducible and Uninterpretable Polymyxin B MICs for Enterobacter cloacae and Enterobacter aerogenes. J. Clin. Microbiol. 2013, 51, 4106–4111. [Google Scholar] [CrossRef] [Green Version]
- Ni, W.; Yang, D.; Guan, J.; Xi, W.; Zhou, D.; Zhao, L.; Cui, J.; Xu, Y.; Gao, Z.; Liu, Y. In vitro and in vivo synergistic effects of tigecycline combined with aminoglycosides on carbapenem-resistant Klebsiella pneumoniae. J. Antimicrob. Chemother. 2021, 76, 2097–2105. [Google Scholar] [CrossRef]
- Tian, Y.; Zhang, Q.; Wen, L.; Chen, J. Combined effect of Polymyxin B and Tigecycline to overcome Heteroresistance in Carbapenem-Resistant Klebsiella pneumoniae. Microbiol. Spectr. 2021, 9, e0015221. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ma, X.; Zhao, L.; He, Y.; Yu, W.; Fu, S.; Ni, W.; Gao, Z. Heteroresistance Is Associated With in vitro Regrowth During Colistin Treatment in Carbapenem-Resistant Klebsiella pneumoniae. Front. Microbiol. 2022, 13, 868991. [Google Scholar] [CrossRef] [PubMed]
- Band, V.I.; Hufnagel, D.A.; Jaggavarapu, S.; Sherman, E.X.; Wozniak, J.E.; Satola, S.W.; Farley, M.M.; Jacob, J.T.; Burd, E.M.; Weiss, D.S. Antibiotic combinations that exploit heteroresistance to multiple drugs effectively control infection. Nat. Microbiol. 2019, 4, 1627–1635. [Google Scholar] [CrossRef] [PubMed]
- Halaby, T.; Kucukkose, E.; Janssen, A.B.; Rogers, M.R.C.; Doorduijn, D.J.; van der Zanden, A.G.M.; al Naiemi, N.; Vandenbroucke-Grauls, C.M.J.E.; van Schaik, W. Genomic Characterization of Colistin Heteroresistance in Klebsiella pneumoniae during a Nosocomial Outbreak. Antimicrob. Agents Chemother. 2016, 60, 6837–6843. [Google Scholar] [CrossRef] [Green Version]
- Poudyal, A.; Howden, B.P.; Bell, J.M.; Gao, W.; Owen, R.J.; Turnidge, J.D.; Nation, R.L.; Li, J. In vitro pharmacodynamics of colistin against multidrug-resistant Klebsiella pneumoniae. J. Antimicrob. Chemother. 2008, 62, 1311–1318. [Google Scholar] [CrossRef]
- Benítez-Cano, A.; Luque, S.; Sorlí, L.; Carazo, J.; Ramos, I.; Campillo, N.; Curull, V.; Sánchez-Font, A.; Vilaplana, C.; Horcajada, J.P.; et al. Intrapulmonary concentrations of meropenem administered by continuous infusion in critically ill patients with nosocomial pneumonia: A randomized pharmacokinetic trial. Crit. Care 2020, 24, 55. [Google Scholar] [CrossRef] [Green Version]
- Erttmann, M.; Krausse, R.; Ullmann, U. Pharmacokinetics of imipenem in patients undergoing major colon surgery. Infection 1990, 18, 367–371. [Google Scholar] [CrossRef]
- Tsuji, B.T.; Pogue, J.M.; Zavascki, A.P.; Paul, M.; Daikos, G.L.; Forrest, A.; Giacobbe, D.R.; Viscoli, C.; Giamarellou, H.; Karaiskos, I.; et al. International Consensus Guidelines for the Optimal Use of the Polymyxins: Endorsed by the American College of Clinical Pharmacy (ACCP), European Society of Clinical Microbiology and Infectious Diseases (ESCMID), Infectious Diseases Society of America (IDSA), International Society for Anti-infective Pharmacology (ISAP), Society of Critical Care Medicine (SCCM), and Society of Infectious Diseases Pharmacists (SIDP). Pharmacotherapy 2019, 39, 10–39. [Google Scholar] [CrossRef] [Green Version]
- Cheong, H.S.; Kim, S.Y.; Wi, Y.M.; Peck, K.R.; Ko, K.S. Colistin Heteroresistance in Klebsiella Pneumoniae Isolates and Diverse Mutations of PmrAB and PhoPQ in Resistant Subpopulations. J. Clin. Med. 2019, 8, 1444. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.; Wi, Y.M.; Kim, J.M.; Kim, Y.-J.; Ko, K.S. Detection of colistin-resistant populations prior to antibiotic exposure in KPC-2-producing Klebsiella pneumoniae clinical isolates. J. Microbiol. 2021, 59, 590–597. [Google Scholar] [CrossRef]
- Morales-León, F.; Lima, C.A.; González-Rocha, G.; Opazo-Capurro, A.; Bello-Toledo, H. Colistin Heteroresistance among Extended Spectrum β-lactamases-Producing Klebsiella pneumoniae. Microorganisms 2020, 8, 1279. [Google Scholar] [CrossRef] [PubMed]
- Bardet, L.; Baron, S.; Leangapichart, T.; Okdah, L.; Diene, S.M.; Rolain, J.-M. Deciphering Heteroresistance to Colistin in a Klebsiella pneumoniae Isolate from Marseille, France. Antimicrob. Agents Chemother. 2017, 61, e00356-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mmatli, M.; Mbelle, N.M.; Maningi, N.E.; Osei Sekyere, J. Emerging Transcriptional and Genomic Mechanisms Mediating Carbapenem and Polymyxin Resistance in Enterobacteriaceae: A Systematic Review of Current Reports. mSystems 2020, 5, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Jayol, A.; Bontron, S.; Villegas, M.-V.; Ozdamar, M.; Türkoglu, S.; Nordmann, P. The mgrB gene as a key target for acquired resistance to colistin in Klebsiella pneumoniae. J. Antimicrob. Chemother. 2015, 70, 75–80. [Google Scholar] [CrossRef]
- Tamma, P.D.; Aitken, S.L.; A Bonomo, R.; Mathers, A.J.; van Duin, D.; Clancy, C.J. Infectious Diseases Society of America 2022 Guidance on the Treatment of Extended-Spectrum β-lactamase Producing Enterobacterales (ESBL-E), Carbapenem-Resistant Enterobacterales (CRE), and Pseudomonas aeruginosa with Difficult-to-Treat Resistance (DTR-P. aeruginosa). Clin. Infect. Dis. 2022, 75, 187–212. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | MLST | COL (mg/L) | GEN (mg/L) | AMI (mg/L) | IMI (mg/L) | MER (mg/L) | TIG (mg/L) | Resistome |
|---|---|---|---|---|---|---|---|---|
| (S ≤ 2; R > 2) | (S ≤ 2; R > 2) | (S ≤ 8; R > 8) | (S ≤ 2; R > 4) | (S ≤ 2; R > 8) | ECOFF * 2 mg/L | |||
| AN1505 | ST323 | 1 | 48 | 4 | 0.25 | 0.05 | 1.5 | aac(3)-IIa, aac(6′)-Ib-cr |
| ITR0244 | ST409 | 1 | 24 | 2 | 0.75 | 0.75 | 4 | aac(3)-IIa, blaOXA-48, aac(6′)-Ib-cr |
| IT0035 | ST15 | 0.5 | 0.25 | 2 | 0.25 | 0.05 | 1.5 | aac(6′)-Ib-cr |
| ATCC 13883 | ST3 | 1 | 0.25 | 1 | 0.75 | 0.03 | 0.75 | _ |
| Colistin-Resistant Subpopulation | Mutation | COL (mg/L) |
|---|---|---|
| AN1505-R1 | mgrB gene interrupted by IS903B of IS5 family at the 70th position | 32 |
| AN1505-R2 | mgrB gene interrupted by IS903B of IS5 family at the 70th position | 32 |
| ITR0244-R1 | mgrB gene interrupted by IS903B of IS5 family at the 117th position | 64 |
| ITR0244-R2 | mgrB promoter region disrupted by ISkpn34 of IS3 family IS elements | 64 |
| IT0035-R1 | mgrB gene interrupted by IS903B of IS5 family at the 70th position | 32 |
| IT0035-R2 | mgrB gene interrupted by ISKpn14 of IS1 family at 123rd position | 32 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajakani, S.G.; Xavier, B.B.; Sey, A.; Mariem, E.B.; Lammens, C.; Goossens, H.; Glupczynski, Y.; Malhotra-Kumar, S. Insight into Antibiotic Synergy Combinations for Eliminating Colistin Heteroresistant Klebsiella pneumoniae. Genes 2023, 14, 1426. https://doi.org/10.3390/genes14071426
Rajakani SG, Xavier BB, Sey A, Mariem EB, Lammens C, Goossens H, Glupczynski Y, Malhotra-Kumar S. Insight into Antibiotic Synergy Combinations for Eliminating Colistin Heteroresistant Klebsiella pneumoniae. Genes. 2023; 14(7):1426. https://doi.org/10.3390/genes14071426
Chicago/Turabian StyleRajakani, Sahaya Glingston, Basil Britto Xavier, Adwoa Sey, El Bounja Mariem, Christine Lammens, Herman Goossens, Youri Glupczynski, and Surbhi Malhotra-Kumar. 2023. "Insight into Antibiotic Synergy Combinations for Eliminating Colistin Heteroresistant Klebsiella pneumoniae" Genes 14, no. 7: 1426. https://doi.org/10.3390/genes14071426