
Whole Exome Sequencing to Find Candidate Variants for the Prediction of Kidney Transplantation Efficacy

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. Whole Exome Sequencing
2.3. Analysis of Variants
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Danovitch, G.M. Handbook of Kidney Transplantation. Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2009; ISBN 978-0781793742. [Google Scholar]
- Gwinner, W. Renal transplant rejection markers. World J. Urol. 2007, 25, 445–455. [Google Scholar] [CrossRef]
- Farid, W.R.; Pan, Q.; van der Meer, A.J.; de Ruiter, P.E.; Ramakrishnaiah, V.; de Jonge, J.; Kwekkeboom, J.; Janssen, H.L.; Metselaar, H.J.; Tilanus, H.W. Hepatocyte-derived microRNAs as serum biomarkers of hepatic injury and rejection after liver transplantation. Liver Transplant. 2012, 18, 290–297. [Google Scholar] [CrossRef]
- Fleissner, F.; Goerzig, Y.; Haverich, A.; Thum, T. Microvesicles as novel biomarkers and therapeutic targets in transplantation medicine. Am. J. Transplant. 2012, 12, 289–297. [Google Scholar] [CrossRef]
- Julich, H.; Willms, A.; Lukacs-Kornek, V.; Kornek, M. Extracellular vesicle profiling and their use as potential disease specific biomarker. Front. Immunol. 2014, 5, 413. [Google Scholar] [CrossRef] [Green Version]
- Pourmand, G.; Emamjomeh, B.; Shabestari, A.N.; Gooran, S.; Namdari, F.; Mehrsai, A.; Dehghani, S.; Soleimani, V.; Aghaii, M.; Bagherpour, F.; et al. The Predictive Value of Serum NGAL for the Diagnosis of Delayed Graft Function in Kidney Transplantation. Transl. Res. Urol. 2021, 3, 182–190. [Google Scholar] [CrossRef]
- Aghamir, S.M.K.; Jamali, R.; Tamehri Zadeh, S.S.; Khajavi, A.; Khatami, F.; Pourmand, G.; Dehghani, S. The Relationship between Graft Function and Liver Function after Kidney Transplantation: A Retrospective Cohort Study. Transl. Res. Urol. 2019, 1, 84–90. [Google Scholar] [CrossRef]
- Yang, J.Y.; Sarwal, M.M. Transplant genetics and genomics. Nat. Rev. Genet. 2017, 18, 309–326. [Google Scholar] [CrossRef]
- Sheldon, S.; Poulton, K. HLA typing and its influence on organ transplantation. Transplant. Immunol. 2006, 157–174. [Google Scholar] [CrossRef]
- Rohland, N.; Reich, D. Cost-effective, high-throughput DNA sequencing libraries for multiplexed target capture. Genome Res. 2012, 22, 939–946. [Google Scholar] [CrossRef] [Green Version]
- Kendig, K.I.; Baheti, S.; Bockol, M.A.; Drucker, T.M.; Hart, S.N.; Heldenbrand, J.R.; Hernaez, M.; Hudson, M.E.; Kalmbach, M.T.; Klee, E.W. Sentieon DNASeq variant calling workflow demonstrates strong computational performance and accuracy. Front. Genet. 2019, 10, 736. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Li, M.; Hakonarson, H. ANNOVAR: Functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010, 38, e164. [Google Scholar] [CrossRef]
- Karczewski, K.J.; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alföldi, J.; Wang, Q.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; Birnbaum, D.P. Variation across 141,456 human exomes and genomes reveals the spectrum of loss-of-function intolerance across human protein-coding genes. BioRxiv 2019, 531210. [Google Scholar]
- Li, Q.; Wang, K. InterVar: Clinical interpretation of genetic variants by the 2015 ACMG-AMP guidelines. Am. J. Hum. Genet. 2017, 100, 267–280. [Google Scholar] [CrossRef] [Green Version]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–423. [Google Scholar] [CrossRef] [Green Version]
- Tavtigian, S.V.; Greenblatt, M.S.; Harrison, S.M.; Nussbaum, R.L.; Prabhu, S.A.; Boucher, K.M.; Biesecker, L.G. Modeling the ACMG/AMP variant classification guidelines as a Bayesian classification framework. Genet. Med. 2018, 20, 1054–1060. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Emond, M.J.; Bamshad, M.J.; Barnes, K.C.; Rieder, M.J.; Nickerson, D.A.; Team, E.L.P.; Christiani, D.C.; Wurfel, M.M.; Lin, X. Optimal unified approach for rare-variant association testing with application to small-sample case-control whole-exome sequencing studies. Am. J. Hum. Genet. 2012, 91, 224–237. [Google Scholar] [CrossRef] [Green Version]
- Chadban, S.J.; Ahn, C.; Axelrod, D.A.; Foster, B.J.; Kasiske, B.L.; Kher, V.; Kumar, D.; Oberbauer, R.; Pascual, J.; Pilmore, H.L. KDIGO clinical practice guideline on the evaluation and management of candidates for kidney transplantation. Transplantation 2020, 104, S11–S103. [Google Scholar] [CrossRef]
- US Department of Health and Human Services. OPTN: Organ Procurement and Transplantation Network. Rep. Liver Transpl. Tex. Transpl. Cent. 2014. Available online: https://optn.transplant.hrsa.gov/data/view-data-reports/regional-data (accessed on 4 May 2016).
- Rahimzadeh, H.; Tamehri Zadeh, S.S.; Khajavi, A.; Saatchi, M.; Reis, L.O.; Guitynavard, F.; Dehghani, S.; Soleimani, V.; Aghamir, S.M.K. The Tsunami of COVID-19 Infection Among Kidney Transplant Recipients: A Single-Center Study from Iran. J. Epidemiol. Glob. Health 2021, 11, 389–396. [Google Scholar] [CrossRef]
- Aghamir, S.M.K. Stem Cells in Urology; Springer Nature: Tehran, Iran, 2020. [Google Scholar]
- Mann, N.; Braun, D.A.; Amann, K.; Tan, W.; Shril, S.; Connaughton, D.M.; Nakayama, M.; Schneider, R.; Kitzler, T.M.; van der Ven, A.T. Whole-exome sequencing enables a precision medicine approach for kidney transplant recipients. J. Am. Soc. Nephrol. 2019, 30, 201–215. [Google Scholar] [CrossRef] [Green Version]
- Mota-Zamorano, S.; Luna, E.; Garcia-Pino, G.; González, L.M.; Gervasini, G. Combined donor-recipient genotypes of leptin receptor and adiponectin gene polymorphisms affect the incidence of complications after renal transplantation. Mol. Genet. Metab. Rep. 2020, 25, 100648. [Google Scholar] [CrossRef]
- Gervasini, G.; Garcia-Pino, G.; Mota-Zamorano, S.; Luna, E.; Garcia-Cerrada, M.; Tormo, M.A.; Cubero, J.J. Association of polymorphisms in leptin and adiponectin genes with long-term outcomes in renal transplant recipients. Pharm. J. 2020, 20, 388–397. [Google Scholar] [CrossRef]
- Kang, E.S.; Magkos, F.; Kim, B.S.; Zhai, R.; Su, L.; Kim, Y.S.; Christiani, D.C.; Lee, H.C.; Mantzoros, C.S. Variants of the adiponectin and adiponectin receptor-1 genes and posttransplantation diabetes mellitus in renal allograft recipients. J. Clin. Endocrinol. Metab. 2012, 97, E129–E135. [Google Scholar] [CrossRef] [Green Version]
- Romanowski, M.; Dziedziejko, V.; Maciejewska-Karlowska, A.; Sawczuk, M.; Safranow, K.; Domanski, L.; Pawlik, A. Adiponectin and leptin gene polymorphisms in patients with post-transplant diabetes mellitus. Pharmacogenomics 2015, 16, 1243–1252. [Google Scholar] [CrossRef]
- Narouie, B.; Saatchi, M. Short term and Long term Survival Rate and Risk Factors of Graft Rejection after Deceased Donor Kidney Transplantation: A Systematic Review and Meta-Analysis. Transl. Res. Urol. 2021, 3, 95–114. [Google Scholar] [CrossRef]
- Arya, P.; Ahadi, Z.; Khajavi, A.; Pourhosein, E.; Dehghani, S.; Soleymani, V.; Zahmatkesh, P.; Keshavarz Pakseresht, B.; Mohammadi, A.; Tavoosian, A. The Effect of Pre-Transplant Psychosocial Problems on Kidney Transplantation Outcomes. Transl. Res. Urol. 2021, 3, 125–130. [Google Scholar] [CrossRef]
- Rahm, A.-K.; Wieder, T.; Gramlich, D.; Müller, M.E.; Wunsch, M.N.; El Tahry, F.A.; Heimberger, T.; Weis, T.; Most, P.; Katus, H.A. HDAC2-dependent remodeling of KCa2. 2 (KCNN2) and KCa2. 3 (KCNN3) K+ channels in atrial fibrillation with concomitant heart failure. Life Sci. 2021, 266, 118892. [Google Scholar] [CrossRef]
- Baheti, S.; Singh, P.; Zhang, Y.; Evans, J.; Jensen, M.D.; Somers, V.K.; Kocher, J.-P.A.; Sun, Z.; Chakkera, H.A. Adipose tissue DNA methylome changes in development of new-onset diabetes after kidney transplantation. Epigenomics 2017, 9, 1423–1435. [Google Scholar] [CrossRef]
- Lee, Y.S.; Saxena, V.; Bromberg, J.S.; Scalea, J.R. G-CSF promotes alloregulatory function of MDSCs through a c-Kit dependent mechanism. Cell. Immunol. 2021, 364, 104346. [Google Scholar] [CrossRef]
- Schnabel, R.B.; Kerr, K.F.; Lubitz, S.A.; Alkylbekova, E.L.; Marcus, G.M.; Sinner, M.F.; Magnani, J.W.; Wolf, P.A.; Deo, R.; Lloyd-Jones, D.M. Large-scale candidate gene analysis in whites and African Americans identifies IL6R polymorphism in relation to atrial fibrillation: The National Heart, Lung, and Blood Institute’s Candidate Gene Association Resource (CARe) project. Circ. Cardiovasc. Genet. 2011, 4, 557–564. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, H.; Higuchi, T.; Yamada, S.; Hirano, T.; Sato, Y.; Nishioka, Y.; Yoshioka, S.; Tatsumi, K.; Ueda, M.; Maeda, M. Human extravillous trophoblasts express laeverin, a novel protein that belongs to membrane-bound gluzincin metallopeptidases. Biochem. Biophys. Res. Commun. 2004, 313, 962–968. [Google Scholar] [CrossRef]
- Maruyama, M.; Hattori, A.; Goto, Y.; Ueda, M.; Maeda, M.; Fujiwara, H.; Tsujimoto, M. Laeverin/aminopeptidase Q, a novel bestatin-sensitive leucine aminopeptidase belonging to the M1 family of aminopeptidases. J. Biol. Chem. 2007, 282, 20088–20096. [Google Scholar] [CrossRef] [Green Version]
- Ng, D.; Johnston, J.J.; Teer, J.K.; Singh, L.N.; Peller, L.C.; Wynter, J.S.; Lewis, K.L.; Cooper, D.N.; Stenson, P.D.; Mullikin, J.C. Interpreting secondary cardiac disease variants in an exome cohort. Circ. Cardiovasc. Genet. 2013, 6, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Gendler, S.J.; Spicer, A.P. Epithelial mucin genes. Annu. Rev. Physiol. 1995, 57, 607–634. [Google Scholar] [CrossRef]
- Wasserberg, N.; Salgar, S.K.; Yang, D.; Ruiz, P.; Ho, S.B.; De Faria, W.L.; Santiago, S.F.; Gandia, C.E.; Miller, J.; Niv, Y. Expression of MUC2 and MUC4 proteins and cytokines: Early markers of intestinal graft rejection1, 2. Transplantation 2003, 75, 1249–1255. [Google Scholar] [CrossRef]
- Sun, L.; Zou, L.-X.; Wang, J.; Chen, T.; Han, Y.-C.; Zhu, D.-D.; Zhuo, S.-C. Mucin 4 gene silencing reduces oxidative stress and calcium oxalate crystal formation in renal tubular epithelial cells through the extracellular signal-regulated kinase signaling pathway in nephrolithiasis rat model. Kidney Blood Press. Res. 2018, 43, 820–835. [Google Scholar] [CrossRef]
- Sribudiani, Y.; Chauhan, R.; Petrova, L.; Harrison, C.; Wabbersen, T.; Brosens, E.; de Graaf, B.; Burzynski, G.; Brouwer, R.; van IJcken, W. Oligogenic inheritance in Hirschsprung disease: Implications of RET and Hedgehog signaling in ENS development. Genet. Funct. Stud. Hirschsprung Dis. 2016, 85. [Google Scholar]
- Coxam, B.; Padberg, Y.; Maier, K.; Jung, S.; Bartels-Klein, E.; Szymborska, A.; Finotto, L.; Schulte-Merker, S.; Gerhardt, H. Svep1 stabilizes developmental vascular anastomosis in reduced flow conditions. Development 2022, 149, dev199858. [Google Scholar] [CrossRef]
- Winkler, M.J.; Müller, P.; Sharifi, A.M.; Wobst, J.; Winter, H.; Miritsch, B.; Hinterdobler, J.; Schreiber, J.; Werner, J.; Stiller, B. Lack of SVEP1 Promotes Vascular Inflammation and Atherosclerotic Plaque Formation. Arterioscler. Thromb. Vasc. Biol. 2020, 40, A325. [Google Scholar]
- Lorenzo, P.; Aspberg, A.; Önnerfjord, P.; Bayliss, M.T.; Neame, P.J.; Heinegård, D. Identification and Characterization of Asporin: A novel member of the leucine-rich repeat protein family closely related to decorin and biglycan*. J. Biol. Chem. 2001, 276, 12201–12211. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wu, H.; Wang, L.; Zhang, H.; Lu, J.; Liang, Z.; Liu, T. Asporin promotes pancreatic cancer cell invasion and migration by regulating the epithelial-to-mesenchymal transition (EMT) through both autocrine and paracrine mechanisms. Cancer Lett. 2017, 398, 24–36. [Google Scholar] [CrossRef]
- Rochette, A.; Boufaied, N.; Scarlata, E.; Hamel, L.; Brimo, F.; Whitaker, H.C.; Ramos-Montoya, A.; Neal, D.E.; Dragomir, A.; Aprikian, A. Asporin is a stromally expressed marker associated with prostate cancer progression. Br. J. Cancer 2017, 116, 775–784. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Jing, X.; Cheng, X.; He, Y.; Hu, L.; Wu, H.; Ye, F.; Zhao, R. Asporin enhances colorectal cancer metastasis through activating the EGFR/src/cortactin signaling pathway. Oncotarget 2016, 7, 73402. [Google Scholar] [CrossRef] [Green Version]
- Yamaba, S.; Yamada, S.; Kajikawa, T.; Awata, T.; Sakashita, H.; Tsushima, K.; Fujihara, C.; Yanagita, M.; Murakami, S. PLAP-1/asporin regulates TLR2-and TLR4-induced inflammatory responses. J. Dent. Res. 2015, 94, 1706–1714. [Google Scholar] [CrossRef] [Green Version]
- Iida, A.; Kizawa, H.; Nakamura, Y.; Ikegawa, S. High-resolution SNP map of ASPN, a susceptibility gene for osteoarthritis. J. Hum. Genet. 2006, 51, 151–154. [Google Scholar] [CrossRef] [Green Version]
- Sakashita, H.; Yamada, S.; Kinoshita, M.; Kajikawa, T.; Iwayama, T.; Murakami, S. Mice lacking PLAP-1/asporin counteracts high fat diet-induced metabolic disorder and alveolar bone loss by controlling adipose tissue expansion. Sci. Rep. 2021, 11, 4970. [Google Scholar] [CrossRef]
- Chang, Y.; Shah, T.; Yang, J.; Min, D.I. Association of genetic polymorphisms of angiopoietin-like 4 with severity of posttransplant proteinuria in kidney allograft recipients. Transpl. Immunol. 2017, 40, 35–41. [Google Scholar] [CrossRef]
- Yildiz, E.; Bardak, H.; Gunay, M.; Bardak, Y.; Imamoglu, S.; Ozbas, H.; Bagci, O. Novel zinc finger protein gene 469 (ZNF469) variants in advanced keratoconus. Curr. Eye Res. 2017, 42, 1396–1400. [Google Scholar] [CrossRef]



{kind=link}
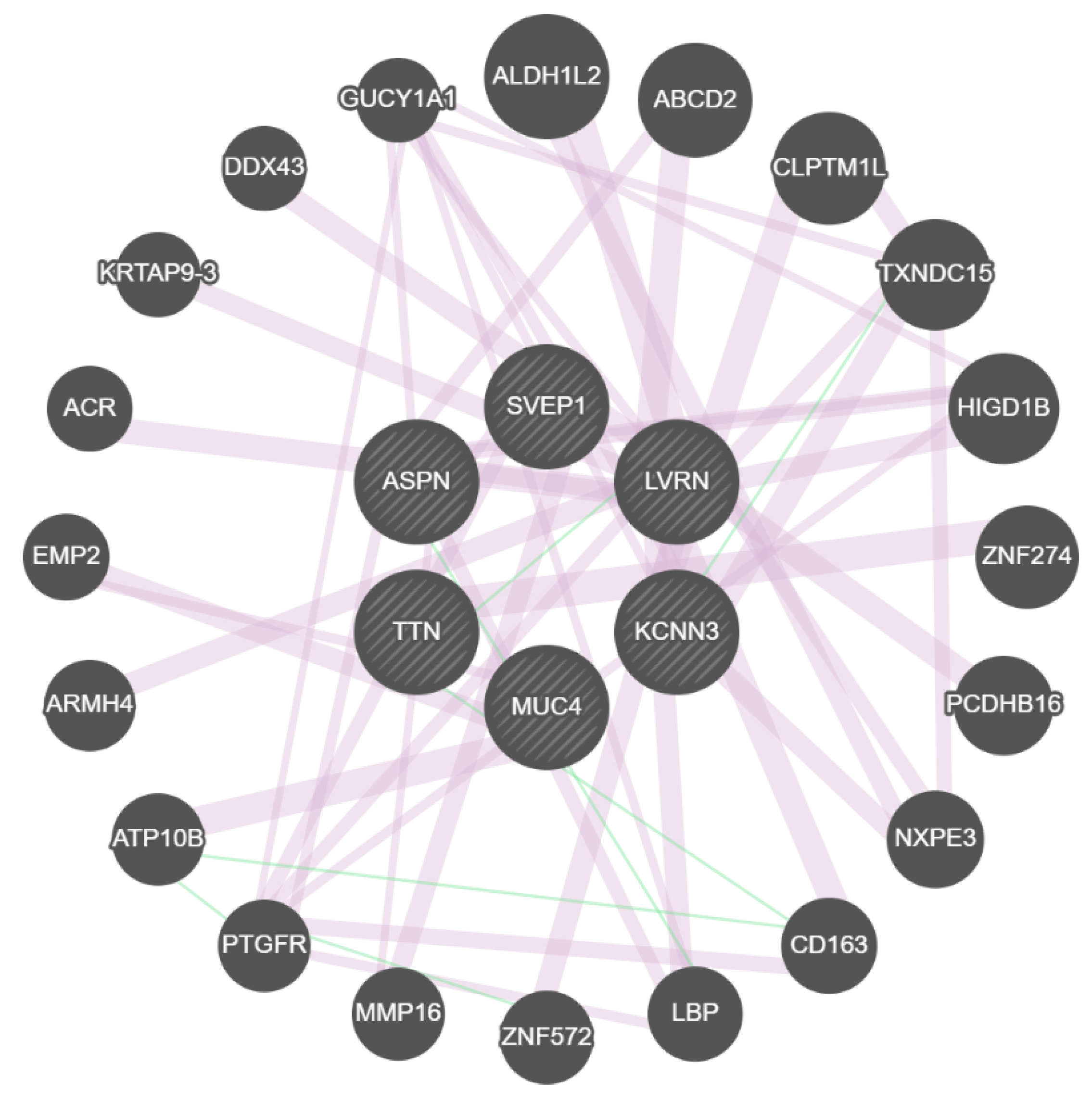{kind=link}
| Variable | Group | p-Value | |
|---|---|---|---|
| Rejecting | Non-Rejecting | ||
| Sex (male), n (%) | 4 (80%) | 4 (80%) | 1.000 |
| Education (BSc), n (%) | 1 (20%) | 3 (60%) | 0.524 |
| Age (year), median (IQR) | 37 (36–43) | 36 (34–42) | 0.459 |
| Creatinine, median (IQR) | 5 (3.6–6) | 1.3 (1.2–1.3) | 0.009 |
| Weight, median (IQR) | 76 (70–76) | 65 (65–87) | 0.750 |
| Height, median (IQR) | 170 (168–170) | 168 (168–168) | 0.142 |
| BMI, median (IQR) | 24.8 (24.0–26.3) | 23.9 (23.0–30.4) | 0.917 |
| DM (yes), n (%) | 2 (40%) | 1 (20%) | 0.500 |
| DM family history (yes), n (%) | 2 (40%) | 1 (20%) | 0.500 |
| Gene | Exonic/ Intronic | SNP | Pathogenicity | Cytoband | Start | End | REF Sequence | ALT Sequence | Depth | |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | LVRN | Exonic | rs564955632 | VUS | 5q23.1 | 115320380 | 115320380 | G | A | 78 |
| 2 | MUC4 | Exonic | rs529922492 | VUS | 3q29 | 195510751 | 195510751 | G | T | 455 |
| 3 | TTN | Intron | rs762675930 | VUS | 2q31.2 | 179611634 | 179611634 | C | T | 199 |
| 4 | TTN | Exonic | rs569593251 | VUS | 2q31.2 | 179406087 | 179406087 | G | A | 105 |
| 5 | TTN | Exonic | rs72648913 | Benign | 2q31.2 | 179610967 | 179610967 | C | T | 166 |
| 6 | SVEP1 | Exonic | rs192347509 | Likely Benign | 9q31.3 | 113170636 | 113170636 | C | T | 98 |
| 7 | SVEP1 | Exonic | rs548514380 | VUS | 9q31.3 | 113261413 | 113261413 | C | T | 146 |
| 8 | ASPN | Intronic | rs779232502 | VUS | 9q22.31 | 95236968 | 95236968 | A | G | 33 |
| 9 | KCNN3 | Exonic | rs3831942 | Likely Benign | 1q21.3 | 154842199 | 154842199 | - | GCTGCTGCTGCTGCT (insertion) | 80 |
| Gene | Exonic/Intronic | SNP | Pathogenicity | Cytoband | Start | End | REF Sequence | ALT Sequence | Depth | |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | ACSF3 | Splicing | - | - | 16q24.3 | 89167068 | 89167068 | - | GGAG | 44 |
| 2 | ANKRD36 | Exonic | rs71329611 | VUS | 2q11.2 | 97864333 | 97864334 | CC | AT | 36 |
| 3 | ANKRD36C | Exonic | rs78179792 | VUS | 2q11.1 | 96616504 | 96616504 | C | A | 103 |
| 4 | ANKRD36C | Exonic | rs78178577 | VUS | 2q11.1 | 96521640 | 96521640 | C | T | 211 |
| 5 | ANKRD36C | Exonic | rs76276218 | VUS | 2q11.1 | 96517474 | 96517474 | G | A | 130 |
| 6 | ANKRD36C | Exonic | rs200183690 | VUS | 2q11.1 | 96521090 | 96521090 | T | C | 108 |
| 7 | ANKRD36C | Exonic | rs77124870 | VUS | 2q11.1 | 96517471 | 96517471 | A | G | 128 |
| 8 | ATXN3 | Exonic;splicing | rs748879218 | Likely Pathogenic | 14q32.12 | 92537397 | 92537397 | T | - | 73 |
| 9 | CDC27 | Exonic | rs111227623 | VUS | 17q21.32 | 45216162 | 45216162 | A | C | 94 |
| 10 | CDC27 | Exonic | rs76836152 | VUS | 17q21.32 | 45216172 | 45216172 | A | G | 94 |
| 11 | CDC27 | Exonic | rs62075657 | VUS | 17q21.32 | 45216132 | 45216132 | T | G | 93 |
| 12 | CDC27 | Exonic | rs796538886 | VUS | 17q21.32 | 45219329 | 45219329 | A | C | 87 |
| 13 | CTDSP2 | Exonic | rs74343811 | VUS | 12q14.1 | 58220841 | 58220841 | C | T | 168 |
| 14 | CTDSP2 | Exonic | rs111346934 | VUS | 12q14.1 | 58220823 | 58220823 | C | T | 165 |
| 15 | CTDSP2 | Exonic | rs76940645 | VUS | 12q14.1 | 58220816 | 58220816 | A | G | 165 |
| 16 | CTDSP2 | Exonic | rs75591888 | VUS | 12q14.1 | 58220844 | 58220844 | C | T | 168 |
| 17 | CTDSP2 | Exonic | rs74554628 | VUS | 12q14.1 | 58240210 | 58240210 | G | T | 115 |
| 18 | DSPP | Exonic | - | - | 4q22.1 | 88537133 | 88537133 | G | A | 38 |
| 19 | FAM104B | Exonic | rs1047037 | Likely Benign | Xp11.21 | 55172687 | 55172687 | T | C | 213 |
| 20 | FRG1BP; FRG1DP | Exonic | rs867116961 | VUS | Xp11.21 | 29612099 | 29612099 | G | A | 151 |
| 21 | FRG1DP; FRG1BP | Exonic | rs1047037 | VUS | Xp11.21 | 29612103 | 29612103 | T | C | 203 |
| 22 | FRG2C | Exonic | rs2118760 | VUS | 3p12.3 | 75715099 | 75715099 | T | A | 235 |
| 23 | HLA-A | Exonic | - | - | 6p22.1 | 29911260 | 29911261 | AC | CG | 116 |
| 24 | HLA-B | Exonic | - | - | 6p21.33 | 31324887 | 31324888 | GG | CT | 73 |
| 25 | HLA-DQA1 | Exonic | rs386699859 | VUS | 6p21.32 | 33037639 | 33037640 | GC | AT | 38 |
| 26 | HLA-DQB1 | Exonic | - | - | 6p21.32 | 32632659 | 32632660 | CT | TG | 57 |
| 27 | HLA-DRB1 | Exonic | rs796196270 | VUS | 6p21.32 | 32551957 | 32551958 | GC | TT | 82 |
| 28 | HLA-DRB1 | Exonic | rs796846373 | VUS | 6p21.32 | 32551938 | 32551939 | GG | AT | 79 |
| 29 | HYDIN | Exonic | rs375727122 | - | 16q22.2 | 70954704 | 70954718 | GCGCTCCTTCTCCGT | - | 182 |
| 30 | HYDIN | Exonic | rs11337008 | Likely Pathogenic | 16q22.2 | 70896016 | 70896016 | A | - | 111 |
| 31 | IGSF3 | Exonic | rs75947003 | VUS | 1p13.1 | 117142700 | 117142700 | C | A | 107 |
| 32 | IGSF3 | Exonic | rs61786651 | VUS | 1p13.1 | 117156459 | 117156459 | C | T | 117 |
| 33 | KRTAP9-1 | Exonic | rs76389571 | VUS | 17q21.2 | 39346578 | 39346578 | C | G | 68 |
| 34 | MUC2 | Exonic | - | - | 11p15.5 | 1093483 | 1093484 | GT | AC | 302 |
| 35 | MUC3A | Exonic | rs113251740 | VUS | 2q21.2 | 100551133 | 100551133 | C | T | 513 |
| 36 | MUC3A | Exonic | rs72494466 | VUS | 7q22.1 | 100551122 | 100551122 | T | G | 448 |
| 37 | MUC3A | Exonic | rs760637480 | VUS | 7q22.1 | 100551094 | 100551094 | G | A | 423 |
| 38 | MUC3A | Exonic | rs771974573 | VUS | 7q22.1 | 100551092 | 100551092 | G | C | 158 |
| 39 | MUC3A | Exonic | rs749410668 | VUS | 7q22.1 | 100551082 | 100551082 | C | T | 170 |
| 40 | MUC3A | Exonic | rs141925032 | VUS | 7q22.1 | 100550704 | 100550704 | C | A | 139 |
| 41 | MUC4 | Exonic | rs773542127 | Likely Benign | 3q29 | 195510254 | 195510254 | C | G | 160 |
| 42 | MUC6 | Exonic | rs374545453 | VUS | 11p15.5 | 1017239 | 1017240 | CG | TA | 263 |
| 43 | OR2L8 | Exonic | rs34851853 | VUS | 1q44 | 248112762 | 248112763 | TG | CA | 78 |
| 44 | PCM1 | Exonic | rs754721723 | VUS | 8p22 | 17796382 | 17796383 | AC | GT | 69 |
| 45 | PLIN4 | Exonic | - | - | 19p13.3 | 4513143 | 4513144 | CA | TG | 107 |
| 46 | PSG3 | Exonic | rs34721205 | VUS | 19q13.2 | 43243217 | 43243218 | AA | GG | 85 |
| 47 | TEX11 | Exonic | rs386825673 | VUS | Xq13.1 | 69749852 | 69749853 | AT | GA | 43 |
| 48 | ZNF717 | Exonic | rs796081257 | VUS | 3p12.3 | rs796081257 | rs796081257 | CA | GG | 144 |
| 49 | PRB3 | Exonic | - | - | 12p13.2 | 11420495 | 11420496 | GG | AA | 53 |
| 50 | RP1L1 | Exonic | rs747592079 | VUS | 8p23.1 | 10466686 | 10466686 | G | A | 103 |
| 51 | SLC35G4 | Exonic | rs386801281 | VUS | 18p11.21 | 11609903 | 11609904 | CA | GG | 150 |
| 52 | ZNF705E | Exonic | - | - | 1q13.4 | 71527895 | 71527896 | CG | TA | 162 |
| 53 | ZNF717 | Exonic | rs77322475 | Likely Benign | 3p12.3 | 75787159 | 75787159 | C | T | 325 |
| 54 | ZNF806 | Exonic | rs11491243 | VUS | 2q21.2 | 133075612 | 133075612 | C | T | 181 |
| 55 | ZNF806 | Exonic | rs113311843 | Likely Pathogenic | 2q21.2 | 133075904 | 133075904 | - | A | 154 |
| 56 | ZNF806 | Exonic | rs111405036 | Likely Pathogenic | 2q21.2 | 133076118 | 133076118 | A | - | 146 |
| 57 | ZNF806 | Exonic | rs111944984 | Likely Pathogenic | 2q21.2 | 133075479 | 133075479 | C | - | 221 |
| 58 | ARHGEF26 | Exonic | rs386667246 | VUS | 3q25.2 | 153839959 | 153839960 | CT | TC | 87 |
| 59 | ARMC9 | Exonic | rs386656198 | Likely Benign | 2q37.1 | 232087474 | 232087475 | AT | GA | 66 |
| 60 | MYG1, MYG1-AS1, PFDN5 | Exonic | rs71453838 | VUS | 12q13.13 | 53693532 | 53693533 | AA | GC | 43 |
| 61 | DUSP5 | Exonic | rs35834951 | VUS | 10q25.2 | 112266822 | 112266823 | GC | AT | 62 |
| 62 | FCGBP | Exonic | rs796880559 | VUS | 19q13.2 | 40368498 | 40368499 | AA | CC | 71 |
| 63 | GTF2IRD2; GTF2IRD2B | Exonic | rs370642824 | VUS | 7q11.23 | 74558397 | 74558397 | C | A | 24 |
| 64 | HLA-C | Exonic | rs796075923 | VUS | 6p21.33 | 31238009 | 31238010 | TT | CG | 31 |
| 65 | IP6K3 | Exonic | rs34332988 | VUS | 6p21.33 | 33690796 | 33690797 | CA | TG | 87 |
| 66 | KLRC3 | Exonic | rs796361824 | VUS | 12p13.2 | 10573094 | 10573095 | CA | GG | 16 |
| 67 | KRTAP10-6 | Exonic | - | - | 21q22.3 | 46012181 | 46012182 | CG | TA | 49 |
| 68 | MPP2 | Exonic | rs70964679 | VUS | 17q21.31 | 41960633 | 41960634 | CG | GC | 83 |
| 69 | MUC12 | Exonic | rs763405288 | Likely Benign | 7q22.1 | 100639418 | 100639419 | CG | AA | 50 |
| 70 | OR2L8 | Exonic | rs34851853 | VUS | 1q44 | 248112762 | 248112763 | TG | CA | 102 |
| 71 | OR2T35 | Exonic | - | - | 1q44 | 248801610 | 248801611 | GC | AT | 35 |
| OR9G1; OR9G9 | Exonic | rs71458233 | VUS | 11q12.1 | 56468047 | 56468048 | AC | GT | 278 | |
| 72 | PCM1 | Exonic | rs754721723 | VUS | 8p22 | 17796382 | 17796383 | AC | GT | 71 |
| 73 | PSG3 | Exonic | rs34721205 | VUS | 19q13.2 | 43243217 | 43243218 | AA | GG | 98 |
| 74 | RGPD5 | Exonic | - | - | 2q13 | 110593536 | 110593536 | C | A | 14 |
| 75 | RNF212 | Exonic | - | - | 4p16.3 | 1087329 | 1087329 | - | TGGAGCCAGCCAT | 44 |
| 76 | SCARF2 | Exonic | rs70944210 | VUS | 22q11.21 | 20779946 | 20779947 | CG | GC | 37 |
| 77 | SPIB | Exonic | rs113934432 | VUS | 19q13.33 | 50926264 | 50926265 | TG | CC | 40 |
| 78 | TCF15 | Exonic | rs71212728 | VUS | 20p13 | 590542 | 590543 | CG | GC | 55 |
| 79 | TRIM50 | Exonic | rs71517080 | VUS | 7q11.23 | 72738762 | 72738763 | CA | TG | 65 |
| 80 | VCX2 | Exonic | Xp22.31 | 8138170 | 8138171 | CT | GC | 19 | ||
| 81 | PCDHA9 | Exonic | rs35021536 | VUS | 5q31.3 | 140230370 | 140230371 | AA | CC | 115 |
| 82 | MUC5B | Exonic | - | - | 11p15.5 | 1258240 | 1258241 | CA | TG | 158 |
| 83 | OR4C3 | Exonic | rs386753295 | VUS | 11p11.2 | 48346961 | 48346962 | AA | TG | 174 |
| 84 | KRTAP12-2 | Exonic | rs35163632 | VUS | 21q22.3 | 46086757 | 46086758 | GC | AT | 83 |
| 85 | PIGZ | Exonic | rs71611508 | VUS | 3q29 | 196674972 | 196674973 | CT | TC | 69 |
| 86 | PRIM2 | Exonic | - | - | 6p11.2 | 57512796 | 57512796 | - | CA | 86 |
| Patient Code | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
| Mediation | B1 100 | Vit B6 | Rocatrol | Rocatrol | Vit B6 | Rocatrol | Metoral | Metoral | Rocatrol | Nistatine |
| Metoral | Amilodip | Amilodip | Amilodip | Prazosin | Calcium D | A.S.A | A.S.A | Calcium D | A.S.A | |
| Amilodip | Ranitiine | NEFROVIT | NEFROVIT | Nistatine | VALCYTE | Vit B6 | Vit B6 | Carvedilol | Vit B6 | |
| Nistatine | Metoral | Allopurinol | Metoral | Rocatrol | Vit B6 | Pantamine | Pantamine | Cinacalcet | Carvedilol | |
| Rocatrol | Nistatine | Tacrolimus | Nistatine | Amilodip | Amilodip | Pantaprozol | Pantaprozol | Diltiazem | Ranitiine | |
| Sevelamer | Pantaprozol | Losartan | Pantaprozol | Calcium D | Nistatine | CellCept | CellCept | Losartan | Rocatrol | |
| VALCYTE | VALCYTE | Omeprazole | VALCYTE | VALCYTE | Allopurinol | Clotrimazole | Clotrimazole | Metformin | VALCYTE | |
| Levofloxacin | Insulin | Atorvastatin | Insulin | Pantaprozol | Isoniazid | Folic Acid | Folic Acid | Pantamine | Furosemide | |
| Pantaprozol | Allopurinol | CellCept | Allopurinol | Atorvastatin | Pantaprozol | Prednisolone | Prednisolone | Pantaprozol | Metformin | |
| Clotrimazole | CellCept | Co-trimoxazole | CellCept | CellCept | Atorvastatin | Rocatrol | Rocatrol | Amlopidine | Pantaprozol | |
| NEFROVIT | Clotrimazole | Folic Acid | Clotrimazole | Clotrimazole | CellCept | Persulfate | Tamsulosin | |||
| Prednisolone | Folic Acid | Hydrochlorothiazide | Folic Acid | Folic Acid | Clotrimazole | NEFROVIT | CellCept | |||
| Sandimmune | Prednisolone | Prednisolone | Prednisolone | Prednisolone | Folic Acid | Omeprazole | Clotrimazole | |||
| Rocaltrol | Sandimmune | Rocaltrol | Prednisolone | Atorvastatin | Folic Acid | |||||
| Sandimmune | Sandimmune | Sandimmune | CellCept | Prednisolone | ||||||
| Clotrimazole | ||||||||||
| Prednisolone | ||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aghamir, S.M.K.; Roudgari, H.; Heidari, H.; Salimi Asl, M.; Jafari Abarghan, Y.; Soleimani, V.; Mashhadi, R.; Khatami, F. Whole Exome Sequencing to Find Candidate Variants for the Prediction of Kidney Transplantation Efficacy. Genes 2023, 14, 1251. https://doi.org/10.3390/genes14061251
Aghamir SMK, Roudgari H, Heidari H, Salimi Asl M, Jafari Abarghan Y, Soleimani V, Mashhadi R, Khatami F. Whole Exome Sequencing to Find Candidate Variants for the Prediction of Kidney Transplantation Efficacy. Genes. 2023; 14(6):1251. https://doi.org/10.3390/genes14061251
Chicago/Turabian StyleAghamir, Seyed Mohammad Kazem, Hassan Roudgari, Hassan Heidari, Mohammad Salimi Asl, Yousef Jafari Abarghan, Venous Soleimani, Rahil Mashhadi, and Fatemeh Khatami. 2023. "Whole Exome Sequencing to Find Candidate Variants for the Prediction of Kidney Transplantation Efficacy" Genes 14, no. 6: 1251. https://doi.org/10.3390/genes14061251