


Genome-Wide Characterization and Sequence Polymorphism Analyses of Glycine max Fibrillin (FBN) Revealed Its Role in Response to Drought Condition

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequence Identification
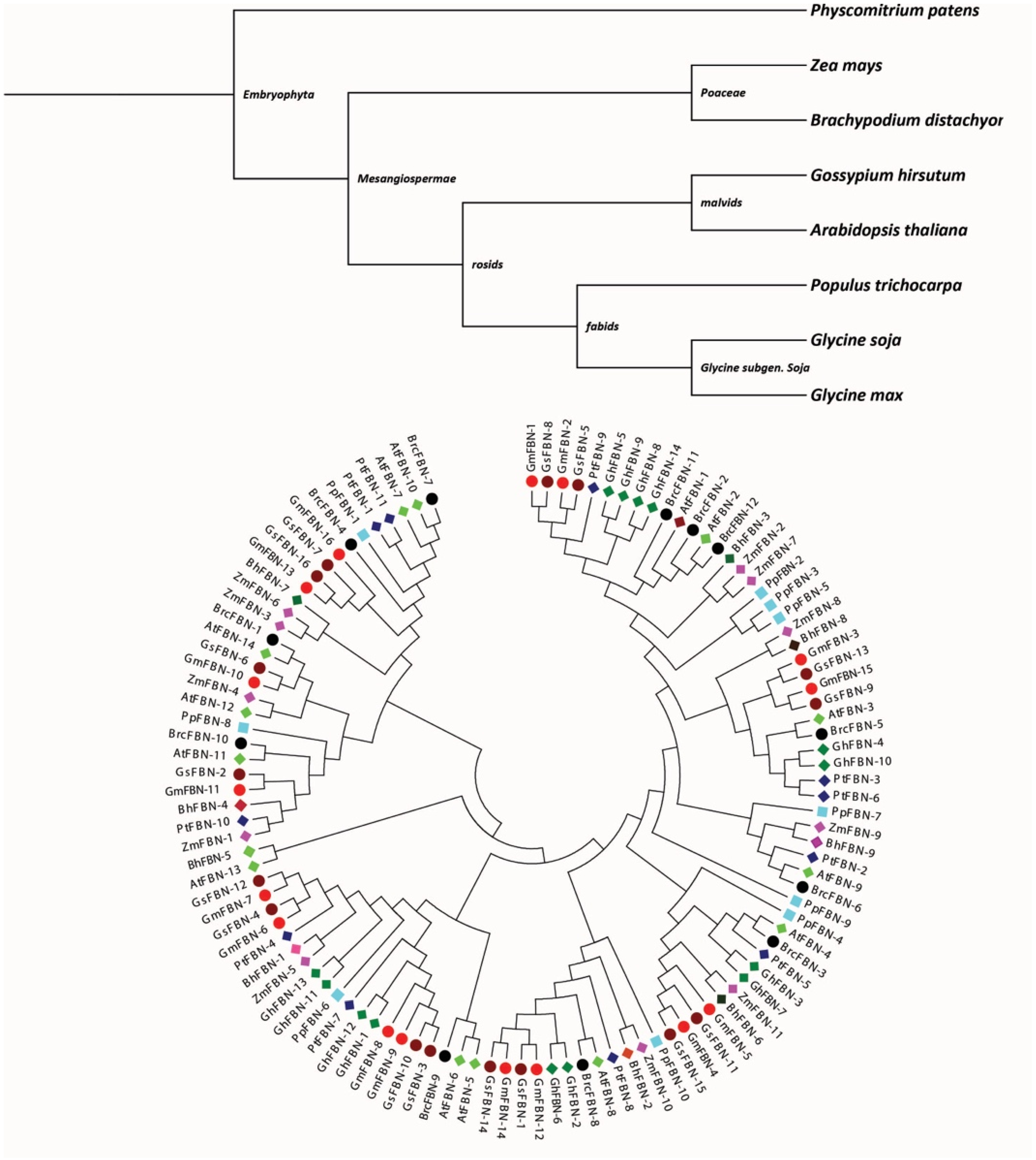
2.2. Phylogenetic Analysis of FBN
2.3. Gene Structure, Protein Motif and Cis-Acting Element Analysis
2.4. In-Silico Prediction and Physiochemical Properties of FBN in G. max
2.5. Sequence Logos of FBN Genes
2.6. Chromosomal Distribution and Synteny analyses of GmFBN
2.7. In-Silico Gene Expression Analysis of GmFBN
2.8. Soybean Plant Material and Phenotyping
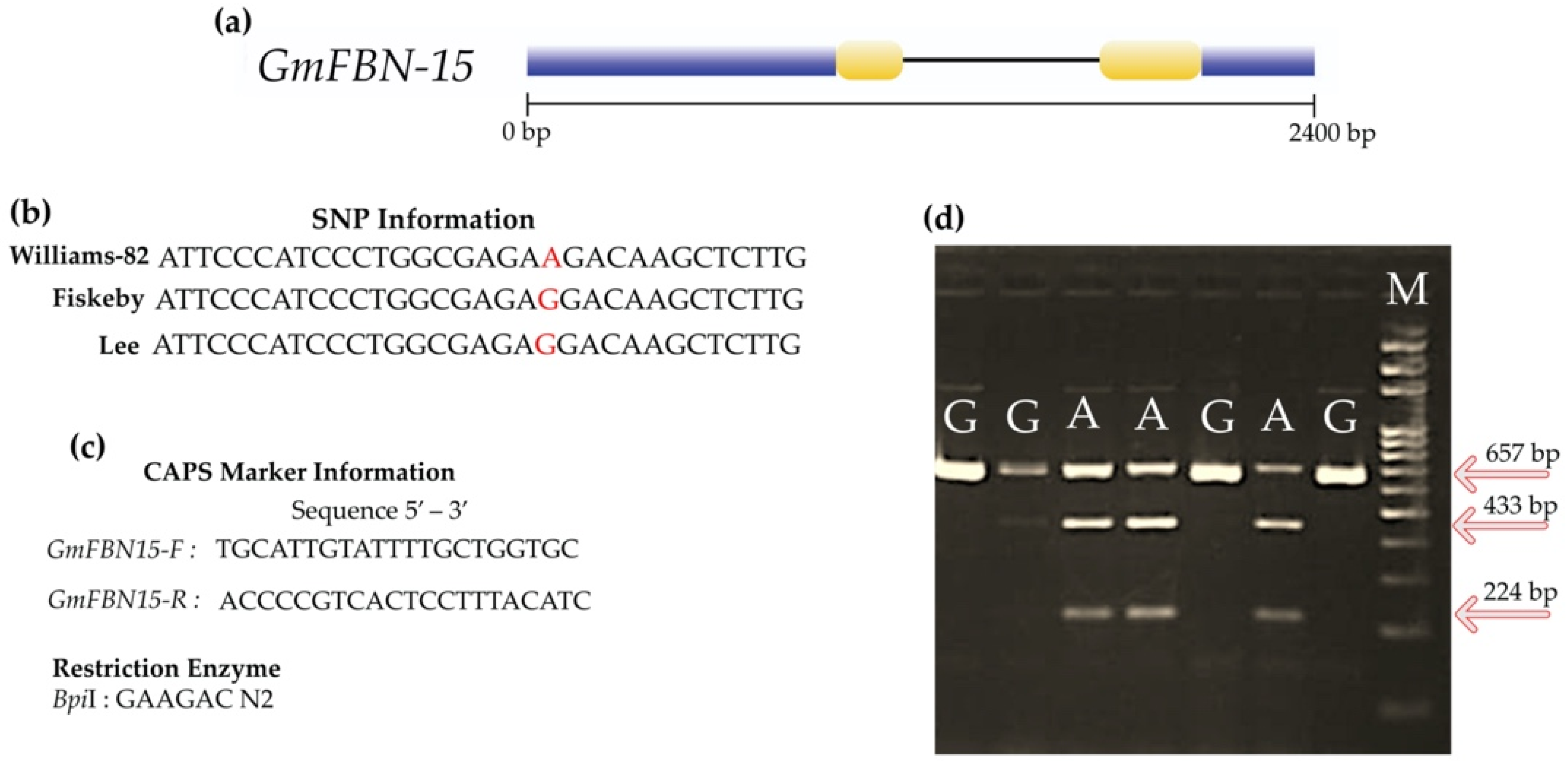
2.9. Development of Single Nucleotide Polymorphism Based Cleaved Amplified Polymorphic Sequence (CAPS) Marker for GmFBN-15
2.10. Statistical Analyses
3. Results
3.1. Identification of FBNs Gene Family Members in Different Species
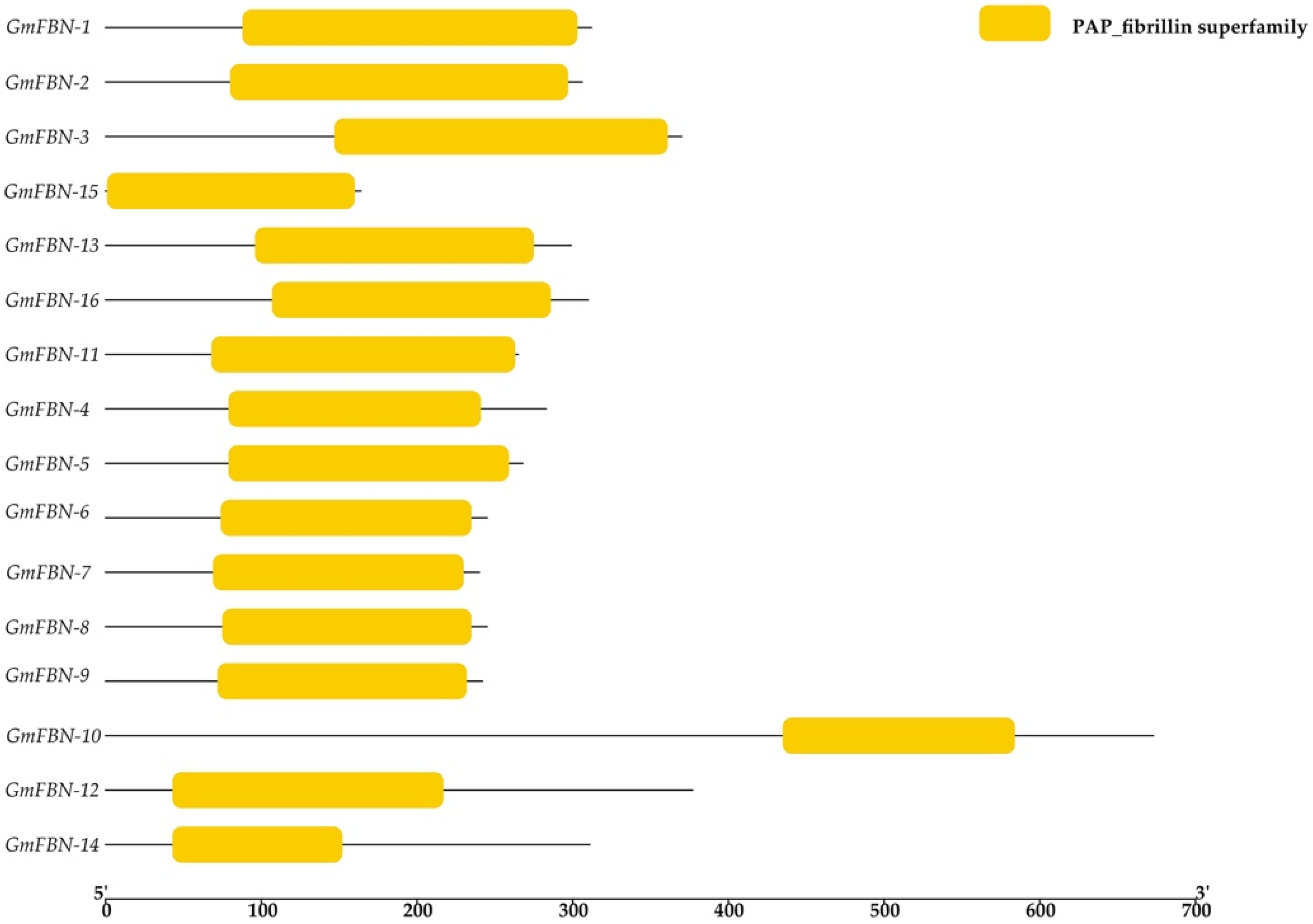
3.2. Conserved Domain, Gene Structure, Protein Motif Analysis and Cis-Acting Elements
3.3. Phylogenetic Analysis of FBNs Gene Family
3.4. In-Silico Prediction and Physiochemical Properties of FBN in G. max
3.5. Chromosomal Distribution, Gene Duplication and Synteny Analysis
3.6. In-Silico Gene Expression Analysis GmFBN
3.7. Development of SNP Based Marker Using CAPS Technique and Marker Trait Association Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nisar, T.; Tahir, M.H.N.; Iqbal, S.; Sajjad, M.; Nadeem, M.A.; Qanmber, G.; Baig, A.; Khan, Z.; Zhao, Z.; Geng, Z.; et al. Genome-wide characterization and sequence polymorphism analyses of cysteine-rich poly comb-like protein in Glycine max. Front. Plant Sci. 2022, 13, 996265. [Google Scholar] [CrossRef] [PubMed]
- Fatima, C.; Tahir, M.H.N.; Ikram, M.; Khan, Z.; Sajjad, M.; Qanmber, G.; Darwish, E.; Geng, Z.; Xiangkuo, G.; Rehman, S.U. Characterization of Histone H3 gene family reveals that GmHH3-3 is associated with higher seed weight in Glycine max. Front. Genet. 2022, 13, 949027. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Zhou, G.; Ma, J.; Jiang, W.; Jin, L.G.; Zhang, Z. De novo assembly of soybean wild relatives for pan-genome analysis of diversity and agronomic traits. Nat. Biotechnol. 2014, 32, 1045–1052. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.H.; Lee, Y.; Kim, H.U. Fibrillin 5 is essential for plastoquinone-9 biosynthesis by binding to solanesyl diphosphas in Arabidopsis. Plant Cell 2015, 27, 2956–2971. [Google Scholar] [PubMed]
- Deruere, J.; Romer, S.; D’harlingue, A.; Backhaus, R.A.; Kuntz, M.; Camara, B. Fibril assembly and carotenoid overaccumulation in chromoplasts: A model for supramolecular lipoprotein structures. Plant Cell 1994, 6, 119–133. [Google Scholar]
- Pozueta-Romero, J.; Rafia, F.; Houlne, G.; Cheniclet, C.; Carde, J.P.; Schantz, M.L.; Schantz, R. A ubiquitous plant housekeeping gene, PAP, encodes a major protein component of bell pepper chromoplasts. Plant Physiol. 1997, 115, 1185–1194. [Google Scholar] [CrossRef]
- Ting, J.T.; Wu, S.S.; Ratnayake, C.; Huang, A.H. Constituents of the tapetosomes and elaioplasts in Brassica campestris tapetum and their degradation and retention during microsporogenesis. Plant J. 1998, 16, 541–551. [Google Scholar] [CrossRef]
- Kim, E.H.; Lee, D.W.; Lee, K.R.; Jung, S.J.; Jeon, J.S.; Kim, H.U. Conserved function of fibrillin 5 in the plastoquinone pathway in Arabidopsis and rice. Front. Plant Sci. 2017, 8, 1197. [Google Scholar] [CrossRef]
- Kim, I.; Lee, S.C.; Kim, E.H.; Song, K.; Yang, T.J.; Kim, H.U. Genome-wide identification and expression analyses of the fibrillin family genes suggest their involvement in photoprotection in cucumber. Plants 2018, 7, 50. [Google Scholar] [CrossRef]
- Jiang, Y.; Hu, H.; Ma, Y.; Zhou, J. Genome-wide identification and characterization of the fibrillin gene family in Triticum aestivm L. PeerJ 2020, 8, e9225. [Google Scholar] [CrossRef]
- Sun, H.; Ren, M.; Zhang, J. Genome-wide identification and expression analyses of fibrillin (FBN) gene family in tomato(Solanum lycopersicum L.). PeerJ 2022, 10, e13414. [Google Scholar] [CrossRef]
- Pandey, A.; Sharma, P.; Mishra, D.; Dey, S.; Malviya, R.; Gayen, D. Genome-wide identification of fibrillin gene family in chickpea (Cicer arietium L.) and its response to drought stress. Intl. J. Biol. Macromol. 2023, 15, 123757. [Google Scholar] [CrossRef]
- Singh, D.K.; McNellis, T.W. Fibrillin protein function the tip of the iceberg? Trends Plant Sci. 2011, 16, 432–441. [Google Scholar] [CrossRef]
- Gasteiger, E.A.; Gattiker, C.; Hoogland, I.; Ivanyi, R.D.; Appel, A.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Briesemeister, S.; Rahnenfuhrer, J.; Kohlbacher, O. YLoc-an interpretable web server for predicting subcellular localization. Nucleic Acids Res. 2010, 38, 497–502. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van De Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE base of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Crooks, G.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef]
- Kumar, A.; Sandhu, N.; Kumruthi, P.; Singh, J.; Kaur, S.; Chhuneja, P. Genome-wide identification and in silico analysis of NPF, NRT2, CLC and SLAC1/SLAH nitrate transporters in hexaploid wheat (Triticum aestivum). Sci. Rep. 2022, 12, 11227. [Google Scholar] [CrossRef]
- Mayrose, I.; Doron-Faigenboim, A.; Bacharach, E.; Pupko, T. Towards realistic codon models: Among site variability and dependency of synonymous and non-synonymous rates. Bioinformatics 2007, 23, i319–i327. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Gong, Q.; Qin, W.; Yang, Z.; Cheng, Y.; Lu, L.; Ge, X.; Zhang, C.; Wu, Z.; Li, F. Genome-wide analysis of WOX genes in upland cotton and their expression pattern under different stresses. BMC Plant Biol. 2017, 17, 113. [Google Scholar] [CrossRef] [PubMed]
- Zafer, Z.M.; Tahir, M.H.N.; Bakhtavar, M.A.; Darwish, E.; Khan, M.A.; Khan, Z.; Fatima, C.; Aftab, A.; Rahman, S.U.; Rehman, S.U. Drought susceptibility index; a preferred criterion in screening for tolerance in soybean. J. Bioresour. Manag. 2023, 10, 8. [Google Scholar]
- Ali, Q.; Salisu, I.B.; Raza, A.; Shahid, A.A.; Rao, A.Q.; Husnain, T.A. Modified protocol for rapid DNA isolation from cotton (Gossypium spp.). MethodsX 2019, 6, 259–264. [Google Scholar] [CrossRef]
- Wei, W.; Liang, D.W.; Bian, X.H.; Shen, M.; Xiao, J.H.; Zhang, W.K.; Ma, B.; Lin, Q.; Lv, J.; Cen, S.Y.; et al. GmWRKY54 improves drought tolerance through activating genes in abscisic acid and Ca2+ signaling pathways in transgenic soybean. Plant J. 2019, 100, 384–398. [Google Scholar] [CrossRef]
- Rincker, K.; Nelson, R.; Specht, J.; Cart, T.; Cianzio, S.R.; Casteel, S.; Conley, S.; Chen, P.; Davis, V.; Fox, C. Genetic improvement of US soybean in maturity groups, II, III and IV. Crop Sci. 2014, 54, 1419–1432. [Google Scholar] [CrossRef]
- Rasheed, A.; Hao, Y.; Xia, X.; Khan, A.; Xu, Y.; Varshney, R.K.; He, Z. Crop breeding chips and genotyping platforms: Progress, challenges, and perspectives. Mol. Plant 2017, 10, 1047–1064. [Google Scholar] [CrossRef]
- Majeed, U.; Darwish, E.; Rehman, S.U.; Zhang, X. Kompetitive allele specific PCR (KASP) a singleplex genotyping platform and its application. J. Agric. Sci. 2018, 11, 11–20. [Google Scholar] [CrossRef]
- Ur Rehman, S.; Sher, M.A.; Saddique, M.A.B.; Ali, Z.; Khan, M.A.; Mao, X.; Irshad, A.; Sajjad, M.; Ikram, R.M.; Naeem, M.; et al. Development and exploitation of KASP assays for genes underpinning drought tolerance among wheat cultivars from Pakistan. Front. Genet. 2021, 12, 684702. [Google Scholar] [CrossRef]
- Yadav, S.; Kushwaha, H.R.; Kumar, K.; Verma, P.K. Comparative structural modelling of monothiol GRX from chickpea: Insight in iron-sulfur cluster assembly. Int. J. Biol. Macromol. 2012, 51, 266–273. [Google Scholar] [CrossRef]
- Bao, J.S.; Corke, E.H.; Sun, E.M. Microsatellites, single nucleotide polymorphisms and a sequence tagged site in starch-synthesizing genes in relation to starch physicochemical properties in non-waxy rice (Oryza sativa L.). Theor. Appl. Genet. 2006, 113, 1185–1196. [Google Scholar] [CrossRef]
- He, Y.; Han, Y.; Jiang, L.; Xu, C.; Lu, J.; Xu, M. Functional analysis of starch-synthesis genes in determining rice eating and cooking qualities. Mol. Breed 2006, 18, 277–290. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene 1 ID | Gene 2 ID | Ka/Ks | Time (MYA) |
|---|---|---|---|
| GmFBN-1 | GmFBN-2 | 0.129409128 | 1 |
| GmFBN-3 | GmFBN-15 | 0.234590794 | 2 |
| GmFBN-12 | GmFBN-14 | 0.172072626 | 1 |
| GmFBN-13 | GmFBN-16 | 0.13874396 | 1 |
| GmFBN-4 | GmFBN-5 | 0.504124568 | 6 |
| GmFBN-10 | GmFBN-7 | 0.482073521 | 3 |
| GmFBN-8 | GmFBN-9 | 0.437348305 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zafer, M.Z.; Tahir, M.H.N.; Khan, Z.; Sajjad, M.; Gao, X.; Bakhtavar, M.A.; Waheed, U.; Siddique, M.; Geng, Z.; Ur Rehman, S. Genome-Wide Characterization and Sequence Polymorphism Analyses of Glycine max Fibrillin (FBN) Revealed Its Role in Response to Drought Condition. Genes 2023, 14, 1188. https://doi.org/10.3390/genes14061188
Zafer MZ, Tahir MHN, Khan Z, Sajjad M, Gao X, Bakhtavar MA, Waheed U, Siddique M, Geng Z, Ur Rehman S. Genome-Wide Characterization and Sequence Polymorphism Analyses of Glycine max Fibrillin (FBN) Revealed Its Role in Response to Drought Condition. Genes. 2023; 14(6):1188. https://doi.org/10.3390/genes14061188
Chicago/Turabian StyleZafer, Muhammad Zeshan, Muhammad Hammad Nadeem Tahir, Zulqurnain Khan, Muhammad Sajjad, Xiangkuo Gao, Muhammad Amir Bakhtavar, Ummara Waheed, Maria Siddique, Zhide Geng, and Shoaib Ur Rehman. 2023. "Genome-Wide Characterization and Sequence Polymorphism Analyses of Glycine max Fibrillin (FBN) Revealed Its Role in Response to Drought Condition" Genes 14, no. 6: 1188. https://doi.org/10.3390/genes14061188