
DNA Methyltransferase Genes Are Associated with Oral Mucositis and Creatinine Levels in Oncopediatric Patients
 , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics and Study Design
2.2. Sample Collection and DNA Extraction
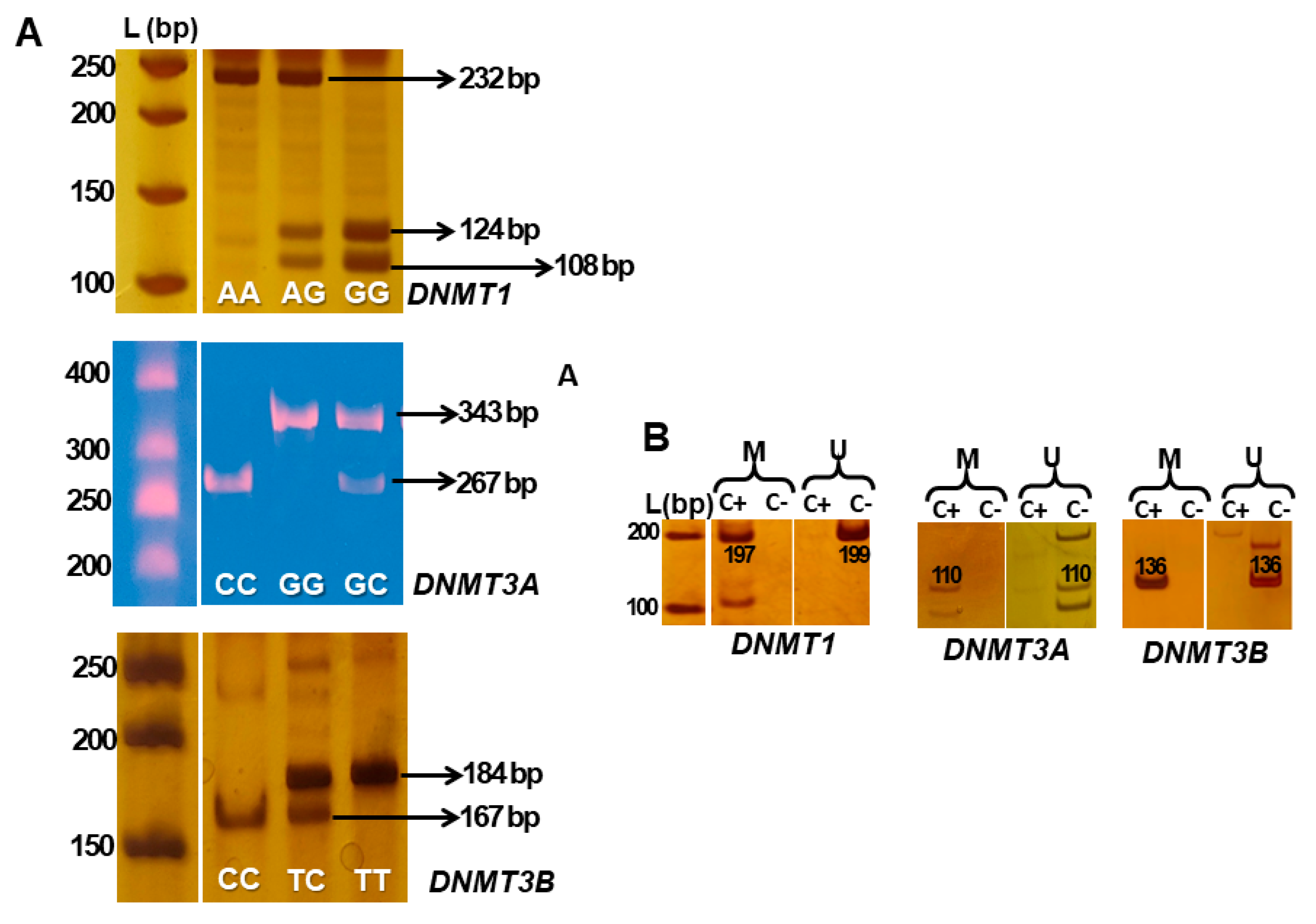
2.3. Polymorphism Analysis
2.4. DNA Methylation Analysis
2.5. Analysis of Genotypes, DNA Methylation Profiles, Blood Cell Counts, and Biochemical Parameters
2.6. Statistical Analysis
3. Results
3.1. Demographic and Clinical Data
3.2. Analysis of Polymorphisms
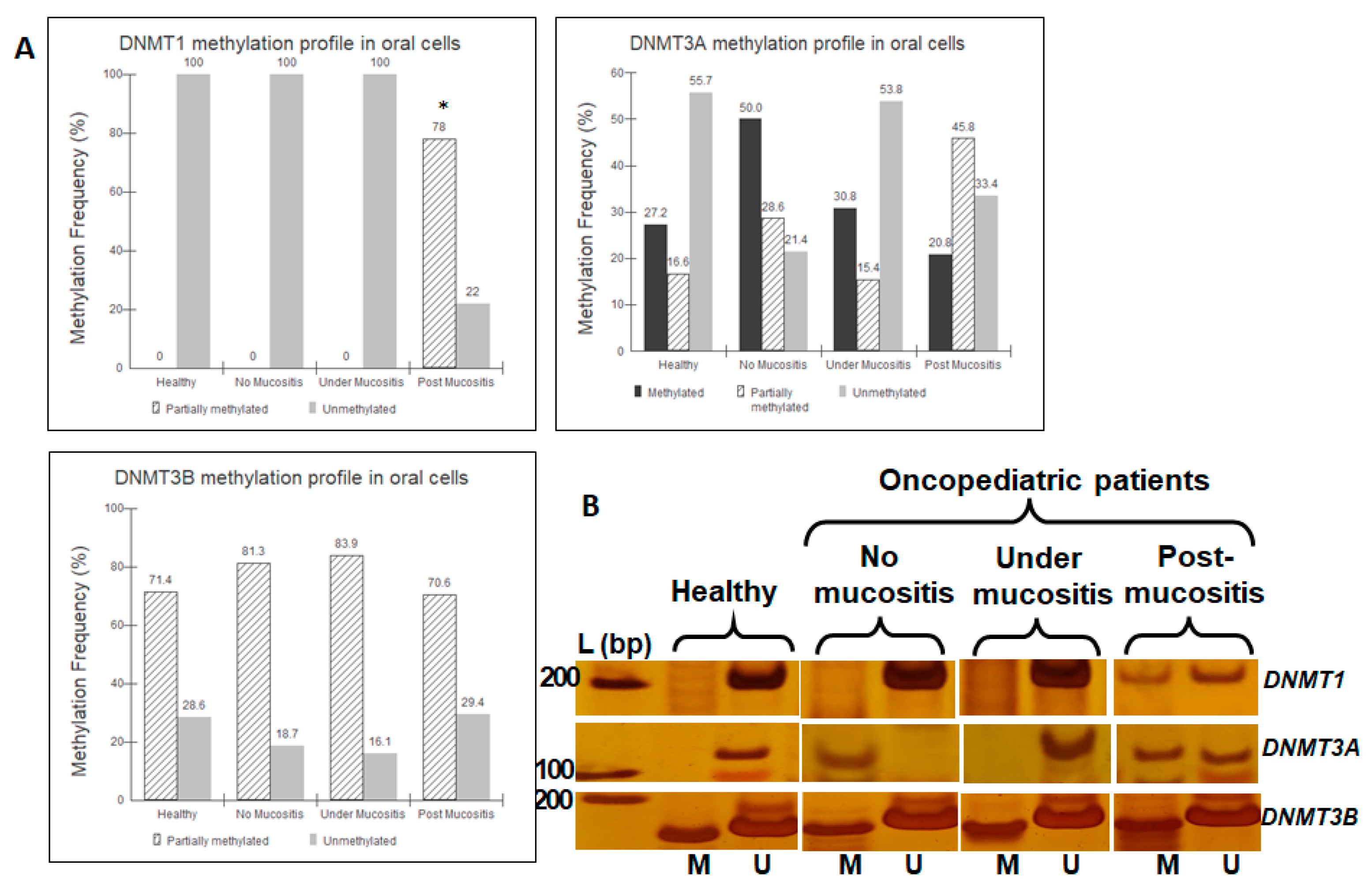
3.3. DNA Methylation Analysis
3.4. Analysis of Genotypes, DNA Methylation Profiles, Blood Cell Counts, and Biochemical Parameters
4. Discussion
4.1. Polymorphisms
4.2. DNA Methylation
4.3. Genotype, DNA Methylation Profile, Blood Cell Count, and Biochemical Parameters
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mattei, A.L.; Bailly, N.; Meissner, A. DNA methylation: A historical perspective. Trends Genet. 2022, 38, 676–707. [Google Scholar] [CrossRef] [PubMed]
- Jeltsch, A.; Broche, J.; Bashtrykov, P. Molecular Processes Connecting DNA Methylation Patterns with DNA Methyltransferases and Histone Modifications in Mammalian Genomes. Genes 2018, 11, 566. [Google Scholar] [CrossRef] [PubMed]
- Leng, S.; Stidley, C.A.; Bernauer, A.M.; Picchi, M.A.; Sheng, X.; Frasco, M.A.; Van Den Berg, D.; Gilliland, F.D.; Crowell, R.E.; Belinsky, S.A. Haplotypes of DNMT1 and DNMT3B are associated with mutagen sensitivity induced by benzo[a]pyrene diol epoxide among smokers. Carcinogenesis 2008, 29, 1380–1385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lu, Q.; Chang, C. Epigenetics in Health and Disease. Adv. Exp. Med. Biol. 2020, 1253, 3–55. [Google Scholar] [CrossRef]
- Murphy, T.M.; Mullins, N.; Ryan, M.; Foster, T.; Kelly, C.; McClelland, R.; O’Grady, J.; Corcoran, E.; Brady, J.; Reilly, M.; et al. Genetic variation in DNMT3B and increased global DNA methylation is associated with suicide attempts in psychiatric patients. Genes Brain Behav. 2013, 12, 125–132. [Google Scholar] [CrossRef]
- Saradalekshmi, K.R.; Neetha, N.V.; Sathyan, S.; Nair, I.V.; Nair, C.M.; Banerjee, M. DNA methyl transferase (DNMT) gene polymorphisms could be a primary event in epigenetic susceptibility to schizophrenia. PLoS ONE 2014, 9, e98182. [Google Scholar] [CrossRef]
- Li, H.; Liu, J.W.; Sun, L.P.; Yuan, Y. A Meta-Analysis of the Association between DNMT1 Polymorphisms and Cancer Risk. BioMed Res. Int. 2017, 2017, 3971259. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Zhou, J.D.; Yang, D.Q.; He, P.F.; Yao, D.M.; Qian, Z. Intragenic hypomethylation of DNMT3A in patients with myelodysplastic syndrome. Clin. Chem. Lab. Med. 2018, 56, 485–491. [Google Scholar] [CrossRef]
- Omidali, M.; Jabbara, N.; Tehrani, G.A. Relationship between promoter hypermethylation of DNMT3A and DNMT3B genes and endometrial cancer. J. Epigenetics 2019, 1, 19–23. [Google Scholar] [CrossRef]
- Fonseca-Silva, T.; Oliveira, M.V.; Fraga, C.A.; Farias, L.C.; Gomes, E.P.; Barros, L.O.; Roy, A.; Santiago Gomez, R.; De Paula, A.M.B.; Guimarães, A.L.S. DNMT3B (C46359T) polymorphisms and immunoexpression of DNMT3b and DNMT1 proteins in oral lichen planus. Pathobiology 2012, 79, 18–23. [Google Scholar] [CrossRef]
- Viana-Filho, J.M.C.; Coêlho, M.C.; Ribeiro, I.L.A.; Persuhn, D.C.; Valença, A.M.G.; Oliveira, N.F.P. ABCG2 polymorphism, age and leukocyte count may contribute to oral mucositis in oncopediatric patients. Braz. Dent. J. 2021, 32, 14–26. [Google Scholar] [CrossRef]
- Sonis, S.T.; Elting, L.S.; Keefe, D.; Peterson, D.E.; Schubert, M.; Hauer-Jensen, M.; Bekele, B.N.; Raber-Durlacher, J.; Donnelly, J.P.; Rubenstein, E.B.; et al. Perspectives on cancer therapy-induced mucosal injury: Pathogenesis, measurement, epidemiology, and consequences for patients. Cancer 2004, 100, 1995–2025. [Google Scholar] [CrossRef]
- Morais, E.F.; Lira, J.A.; Macedo, R.A.; Santos, K.S.; Elias, C.T.; Morais, M.L.S.A. Oral manifestations resulting from chemotherapy in children with acute lymphoblastic leukemia. Braz. J. Otorhinolaryngol. 2014, 80, 78–85. [Google Scholar] [CrossRef]
- Moe, P.J.; Holen, A. High-dose methotrexate in childhood all. Pediatr. Hematol. Oncol. J. 2000, 17, 615–622. [Google Scholar] [CrossRef]
- Taylor, Z.L.; Vang, J.; Lopez-Lopez, E.; Oosterom, N.; Mikkelsen, T.; Ramsey, L.B. Systematic Review of Pharmacogenetic Factors That Influence High-Dose Methotrexate Pharmacokinetics in Pediatric Malignancies. Cancers 2021, 13, 2837. [Google Scholar] [CrossRef]
- Otmani, N.; Hattad, S. Clinical Outcome in Children with Chemotherapy-Induced Mucositis. Semin. Oncol. Nurs. 2021, 37, 151160. [Google Scholar] [CrossRef]
- Coêlho, M.C.; Viana-Filho, J.M.C.; Souza, B.F.; Valença, A.M.G.; Persuhn, D.C.; Oliveira, N.F.P. Genetic polymorphisms of genes involved in oxidative stress and inflammatory management in oncopediatric patients with chemo-induced oral mucositis. J. Appl. Oral. Sci. 2022, 30, e20210490. [Google Scholar] [CrossRef]
- Viana-Filho, J.M.C.; de Souza, B.F.; de Castro Coêlho, M.; Valença, A.M.G.; Persuhn, D.C.; de Oliveira, N.F.P. Polymorphism but not methylation status in the VDR gene contributes to oral mucositis in children. Oral Dis. 2022; Online ahead of print. [Google Scholar] [CrossRef]
- Oosterom, N.; Griffioen, P.H.; den Hoed, M.A.H.; Pieters, R.; de Jonge, R.; Tissing, W.J.E.; van den Heuvel-Eibrink, M.M.; Heil, S.G. Global methylation in relation to methotrexate-induced oral mucositis in children with acute lymphoblastic leukemia. PLoS ONE 2018, 13, e0199574. [Google Scholar] [CrossRef]
- Ferguson, L.R.; De Caterina, R.; Görman, U.; Allayee, H.; Kohlmeier, M.; Prasad, C.; Choi, M.S.; Curi, R.; De Luis, D.A.; Gil, Á.; et al. Guide and Position of the International Society of Nutrigenetics/Nutrigenomics on Personalised Nutrition: Part 1-Fields of Precision Nutrition. J. Nutrigenet. Nutr. 2016, 9, 12–27. [Google Scholar] [CrossRef]
- Cheng, K.K.; Chang, A.M.; Yuen, M.P. Prevention of oral mucositis in paediatric patients treated with chemotherapy; a randomised crossover trial comparing two protocols of oral care. Eur. J. Cancer 2004, 40, 1208–1216. [Google Scholar] [CrossRef]
- Aidar, M.; Line, S.R. A simple and cost-effective protocol for DNA isolation from buccal epithelial cells. Braz. Dent. J. 2007, 18, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Karpinski, P.; Myszka, A.; Ramsey, D.; Misiak, B.; Gil, J.; Laczmanska, I.; Grzebieniak, Z.; Sebzda, T.; Smigiel, R.; Stembalska, A.; et al. Polymorphisms in methyl-group metabolism genes and risk of sporadic colorectal cancer with relation to the CpG island methylator phenotype. Cancer Epidemiol. 2010, 34, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.A.; Azova, M.M.; Ramazanova, F.U.; Gigani, O.B. DNMT1 and DNMT3A Gene Polymorphisms and Early Pregnancy Loss. Russ. J. Genet. 2020, 56, 379–382. [Google Scholar] [CrossRef]
- Dou, C.Y.; Fan, Y.C.; Cao, C.J.; Yang, Y.; Wang, K. Sera DNA Methylation of CDH1, DNMT3b and ESR1 Promoters as Biomarker for the Early Diagnosis of Hepatitis B Virus-Related Hepatocellular Carcinoma. Dig. Dis. Sci. 2016, 61, 1130–1138. [Google Scholar] [CrossRef] [PubMed]
- Zebardast, S.; Sahmani, M.; Mohammadi, S.; Foroughi, F.; Dehghani Fard, A.; Mohammadi, Z.; Khojastepour, S.; Azad, M. The Gene Expression Profile and DNA Methylation Pattern of CDH1 and DNMT1 Genes in Acute Promyelocytic Leukemia (APL). Rep. Biochem. Mol. Biol. 2020, 8, 454–457. [Google Scholar] [PubMed]
- Xu, Z.; Taylor, J.A. SNPinfo: Integrating GWAS and candidate gene information into functional SNP selection for genetic association studies. Nucleic Acids Res. 2009, 37, W600–W605. [Google Scholar] [CrossRef]
- Mostowska, A.; Sajdak, S.; Pawlik, P.; Lianeri, M.; Jagodzinski, P.P. DNMT1, DNMT3A and DNMT3B gene variants in relation to ovarian cancer risk in the Polish population. Mol. Biol. Rep. 2013, 40, 4893–4899. [Google Scholar] [CrossRef]
- Veena, K.V.; Siddamalla, S.; Deenadayal, M.; Shivaji, S.; Bhanoori, M. DNMT1 and DNMT3B gene variants and their association with endometriosis in South Indian women. Mol. Biol. Rep. 2020, 49, 321–329. [Google Scholar] [CrossRef]
- Barišić, A.; Kolak, M.; Peterlin, A.; Tul, N.; Gašparović Krpina, M.; Ostojić, S.; Peterlin, B.; Pereza, N. DNMT3B rs1569686 and rs2424913 gene polymorphisms are associated with positive family history of preterm birth and smoking status. Croat. Med. J. 2020, 61, 8–17. [Google Scholar] [CrossRef]
- Yuan, X.Q.; Zhang, D.Y.; Yan, H.; Yang, Y.L.; Zhu, K.W.; Chen, Y.H.; Li, X.; Yin, J.Y.; Li, X.L.; Zeng, H.; et al. Evaluation of DNMT3A genetic polymorphisms as outcome predictors in AML patients. Oncotarget 2016, 7, 60555–60574. [Google Scholar] [CrossRef]
- Guo, H.; Cheng, J.; Sheng, R.; Cai, W.; Xin, Y.; Ding, E.; Zhang, H.; Zhu, B. Association of DNMT3a’s single nucleotide polymorphisms with susceptibility to noise-induced hearing loss in Chinese workers. Wei Sheng Yan Jiu 2018, 47, 638–643. [Google Scholar] [PubMed]
- Szczepańska, M.; Mostowska, A.; Wirstlein, P.; Malejczyk, J.; Płoski, R.; Skrzypczak, J.; Jagodziński, P.P. Polymorphic variants of DNMT3A and the risk of endometriosis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 166, 81–85. [Google Scholar] [CrossRef]
- Lee, S.J.; Jeon, H.S.; Jang, J.S.; Park, S.H.; Lee, G.Y.; Lee, B.H.; Kim, C.H.; Kang, Y.M.; Lee, W.K.; Kam, S.; et al. DNMT3B polymorphisms and risk of primary lung cancer. Carcinogenesis 2005, 26, 403–409. [Google Scholar] [CrossRef]
- De Castro, T.B.; Rodrigues-Fleming, G.H.; Oliveira-Cucolo, J.G.; Silva, J.N.G.D.; Silva, F.P.; Raposo, L.S.; Maniglia, J.V.; Pavarino, É.C.; Arantes, L.M.R.B.; Galbiatti-Dias, A.L.S.; et al. Gene Polymorphisms Involved in Folate Metabolism and DNA Methylation with the Risk of Head and Neck Cancer. Asian Pac. J. Cancer Prev. 2020, 21, 3751–3759. [Google Scholar] [CrossRef]
- Akiyama, N.; Yamamoto-Fukuda, T.; Yoshikawa, M.; Kojima, H. Regulation of DNA methylation levels in the process of oral mucosal regeneration in a rat oral ulcer model. Histol. Histopathol. 2010, 35, 247–256. [Google Scholar] [CrossRef]
- Cai, L.; Zhan, M.; Li, Q.; Li, D.; Xu, Q. DNA methyltransferase DNMT1 inhibits lipopolysaccharide-induced inflammatory response in human dental pulp cells involving the methylation changes of IL-6 and TRAF6. Mol. Med. Rep. 2020, 21, 959–968. [Google Scholar] [CrossRef]
- Viana-Filho, J.M.C.; Coêlho, M.C.; de Queiroz Neto, J.N.; de Souza, B.F.; Valença, A.M.G.; de Oliveira, N.F.P. TNF-α promoter hypomethylation is associated with occurrence of oral mucositis in oncopediatric patients treated with methotrexate. Braz. Oral Res. 2023, submitted.
- Bozack, A.K.; Cardenas, A.; Geldhof, J.; Quamruzzaman, Q.; Rahman, M.; Mostofa, G.; Christiani, D.C.; Kile, M.L. Cord blood DNA methylation of DNMT3A mediates the association between in utero arsenic exposure and birth outcomes: Results from a prospective birth cohort in Bangladesh. Environ. Res. 2020, 183, 109134. [Google Scholar] [CrossRef]
- Choi, S.J.; Jung, S.W.; Huh, S.; Chung, Y.S.; Cho, H.; Kang, H. Alteration of DNA Methylation in Gastric Cancer with Chemotherapy. J. Microbiol. Biotechnol. 2017, 27, 1367–1378. [Google Scholar] [CrossRef]
- Santana Bezerra, H.; Severo de Assis, C.; Dos Santos Nunes, M.K.; Wanderley de Queiroga Evangelista, I.; Modesto Filho, J.; Alves Pegado Gomes, C.N.; Ferreira do Nascimento, R.A.; Pordeus Luna, R.C.; de Carvalho Costa, M.J.; de Oliveira, N.F.P.; et al. The MTHFR promoter hypermethylation pattern associated with the A1298C polymorphism influences lipid parameters and glycemic control in diabetic patients. Diabetol. Metab. Syndr. 2019, 11, 4. [Google Scholar] [CrossRef]
- Bednarska-Makaruk, M.; Graban, A.; Sobczyńska-Malefora, A.; Harrington, D.J.; Mitchell, M.; Voong, K.; Dai, L.; Łojkowska, W.; Bochyńska, A.; Ryglewicz, D.; et al. Homocysteine metabolism and the associations of global DNA methylation with selected gene polymorphisms and nutritional factors in patients with dementia. Exp. Gerontol. 2016, 81, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Damascena, L.C.L.; de Lucena, N.N.N.; Ribeiro, I.L.A.; Pereira, T.L.; Lima-Filho, L.M.A.; Valença, A.M.G. Severe Oral Mucositis in Pediatric Cancer Patients: Survival Analysis and Predictive Factors. Int. J. Environ. Res. Public. Health 2020, 17, 1235. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.L.; Zhao, F.Y.; Song, H.; Shen, D.Y.; Xu, X.J. Methotrexate Associated Renal Impairment Is Related to Delayed Elimination of High-Dose Methotrexate. Sci. World. J. 2015, 2015, 751703. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Analysis | Primers (5′-3′) | Annealing (°C) | Product (bp) | |
|---|---|---|---|---|
| Polymorphism | RE-RS SNP Location | |||
| DNMT1 rs2228611 (G>A) | F: TATGTTGTCCAGGCTCGTCTC R: GTACTGTAAGCACGGTCACCTG | 55 | BsmaI GTCTC(1/5)^ exon 17 | AA: 232, 28 AG: 232, 108, 124, 28 GG: 108, 124, 28 |
| DNMT3A rs7590760 (C>G) | F: TGCTGTGCCTACTCCAAACA R: GCCATGAATGTCCAGAAGGT | 62.6 | RsaIGT^AC Intron 6 | CC: 267, 76 CG: 267, 76, 343 GG: 343 |
| DNMT3B rs6087990 (T>C) | F: GAAAAAGGCCCCAGAAGGC R: GGCGGGGACGAGGGAAATTT | 66 | BanI G^GYRCC promoter | TT: 184 TC: 184,167, 17 CC: 167,17 |
| DNA methylation | CpG location | |||
| DNMT1 | M F: TTAGTAAATCGTGGAGTTTGGAC M R: AACGATAAACGAAAACGACG U F: AGTAAATTGTGGAGTTTGGAT U R: AAAAAACAATAAACAAAAACAACATCT | 50 55 | promoter | 197 199 |
| DNMT3A | M F: GGGAGGTATTTGATATCGGTTC M R: AAACTTCGATAACTCCACCTAACG U F: GGGGAGGTATTTGATATTGGTTT U R: AACTTCAATAACTCCACCTAACACT | 51 56 | exon 1 | 110 110 |
| DNMT3B | M F: GATGATAGGTAGGGGTATCGC M R: CGAACGAAAACACACACACA U F: GTAGGATGATAGGTAGGGGTATTGT U R: ACCCAAACAAAAACACACACAC | 59 63 | promoter | 136 136 |
| G1 No Mucositis during Chemotherapy (n = 16) | G2 Mucositis during Chemotherapy (n = 86) | G3 Mild/Moderate Mucositis (n = 23) | G4 Severe Mucositis (n = 26) | |
|---|---|---|---|---|
| Age (years) Mean (±SD) | 10.1 (±3.6) | 10.4 (±4.9) | 8.7 (±4.8) | 10.8 (±4.6) |
| Sex—n (%) | ||||
| Girls | 11 (68.8) | 32 (37.2) | 10 (43.5) | 12 (46.0) |
| Boys | 05 (31.3) | 54 (62.7) * | 13 (56.5) | 14 (54.0) |
| Cancer—n (%) | ||||
| ALL | 11 (68.8) | 65 (75.5) | 20 (87.0) | 19 (73.0) |
| AML | 02 (12.5) | 10 (11.6) | 02 (9.0) | 02 (8.0) |
| APL | 01 (6.3) | 02 (2.3) | - | 02 (8.0) |
| CML | 01 (6.3) | 01 (1.2) | - | - |
| HL | 01 (6.3) | 01 (1.2) | 01 (4.0) | - |
| NHL | - | 07 (8.2) | - | 03 (12.0) |
| Healthy (n = 21) | No Mucositis (n = 16) | Under Mucositis (n = 17) | Post-Mucositis (n = 31) | |
|---|---|---|---|---|
| Age (years) Mean (±SD) | 10.2 (3.2) | 10.1 (3.6) | 10 (4.3) | 12.7 (4.7) |
| Sex—n (%) | ||||
| Girls | 14 (66.7) | 11 (68.8) | 7 (41.2) | 14 (45.2) |
| Boys | 7 (33.3) | 5 (31.3) | 10 (58.8) | 17 (54.8) |
| Cancer—n (%) | ||||
| ALL | - | 11 (68.8) | 12 (70.6) | 25 (80.6) |
| AML | - | 02 (12.5) | 02 (11.8) | 02 (6.5) |
| APL | - | 01 (6.3) | - | 02 (6.5) |
| CML | - | 01 (6.3) | - | 01 (3.2) |
| HL | - | 01 (6.3) | - | - |
| NHL | - | - | 03 (17.6) | 01 (3.2) |
| Without cancer | 21 (100) | - | - | - |
| SNP | No Mucositis during Chemotherapy 1 (n = 16) | Mucositis during Chemotherapy 2 (n = 86) | p-Valor (1 and 2) | Mild/Moderate Mucositis 3 (n = 23) | Severe Mucositis 4 (n = 26) | p-Valor (3 and 4) |
|---|---|---|---|---|---|---|
| DNMT1 (A > G) | 0.541 € | 0.458 € | ||||
| n (%) | ||||||
| AA | 06 (37.5) | 26 (30.2) | 05 (21.7) | 10 (38.5) | ||
| AG | 09 (56.3) | 44 (51.2) | 13 (56.6) | 11 (42.3) | ||
| GG | 01 (6.3) | 16 (18.6) | 05 (21.7) | 05 (19.2) | ||
| A | 21 (65.6) | 96 (55.9) | 0.302 | 23 (50.0) | 31 (59.6) | 0.339 ¥ |
| G | 11 (34.4) | 76 (44.1) | 23 (50.0) | 21 (40.4) | ||
| HWE (p) | 0.323 | 0.729 | - | 0.531 | 0.536 | - |
| DNMT3A (G > C) | 0.251 € | 0.691 € | ||||
| n (%) | ||||||
| GG | 07 (43.8) | 23 (26.7) | 07 (30.4) | 08 (30.8) | ||
| GC | 08 (50.0) | 45 (52.3) | 13 (56.5) | 12 (46.2) | ||
| CC | 01 (06.3) | 18 (20.9) | 03 (13.0) | 06 (23.1) | ||
| G | 22 (71.0) | 91 (52.9) | 0.062 ¥ | 27 (58.6) | 28 (53.8) | 0.629 ¥ |
| C | 9 (29.0) | 81 (47.1) | 19 (41.3) | 24 (46.2) | ||
| HWE (p) | 0.512 | 0.642 | - | 0.426 | 0.715 | - |
| DNMT3B (T > C) | 0.610 € | 0.676 € | ||||
| n (%) | ||||||
| TT | 03 (18.9) | 26 (30.2) | 06 (26.1) | 10 (38.5) | ||
| TC | 08 (50.0) | 39 (45.3) | 10 (43.5) | 09 (34.6) | ||
| CC | 05 (31.3) | 21 (24.4) | 07 (30.4) | 07 (26.9) | ||
| T | 14 (43.7) | 91 (52.9) | 0.341 ¥ | 22 (47.8) | 29 (55.8) | 0.432 ¥ |
| C | 18 (56.3) | 81 (47.1) | 24 (52.2) | 23 (44.2) | ||
| HWE (p) | 0.949 | 0.404 | - | 0.536 | 0.128 | - |
| DNMT1 Genotype | Methylated | Partially Methylated (n = 24) | Unmethylated (n = 40) | ||||||
| --- | AA (n = 8) | AG + GG (n = 16) | p | AA (n = 12) | AG + GG (n = 28) | p | |||
| Hemoglobin | 9.063 ± 2.536 | 9.200 ± 1.666 | 0.874 | 9.342 ± 1.498 | 9.721 ± 2.527 | 0.632 | |||
| Leukocytes | 3850 (1100–6400) | 2450 (100–17,500) | 0.444 | 3400 (100–7700) | 3900 (900–42,400) | 0.244 | |||
| Platelets | 129,000 (54,000–302,000) | 134,500 (23,000–533,000) | 0.653 | 140,500 (27,000–415,000) | 169,000 (28,000–561,000) | 0.384 | |||
| Urea | 17.5 (9.00–33.0) | 19.0 (9.00- 38.0) | 0.581 | 16.5 (10.0–61.0) | 19.0 (5.10–69.0) | 0.545 | |||
| Creatinine | 0.471 ± 0.140 | 0.512 ± 0.145 | 0.513 | 0.454 ± 0.134 | 0.502 ± 0.172 | 0.391 | |||
| DNMT3A Genotype | Methylated (n = 16) | Partially Methylated (n = 17) | Unmethylated (n = 18) | ||||||
| GG + GC (n = 13) | CC (n = 3) | p | GG + GC (n = 12) | CC (n = 5) | p | GG + GC (n = 14) | CC (n = 4) | p | |
| Hemoglobin | 9.238 ± 1.319 | 10.867 ± 0.473 | 0.058 | 10.600 ± 1.923 | 9.360 ± 1.996 | 0.249 | 9.000 ± 2.042 | 7.550 ± 3.044 | 0.275 |
| Leukocytes | 2900 (1100–42,400) | 2200 (1400–3200) | 0.239 | 3950 (100–8000) | 6300 (900–7100) | 0.712 | 3250 (100–22300) | 1900 (1000–5300) | 0.426 |
| Platelets | 145,000 (23,000–471,000) | 175,000 (56,000–242,000) | 1.000 | 161,500 (41,000–380,000) | 152,000 (28,000–302,000) | 0.460 | 118,500 (35,000–561,000) | 143,500 (103,000–283,000) | 0.721 |
| Urea | 17.0 (9.00–35.0) | 16.0 (12.0–16.0) | 0.417 | 22.0 (13.0–69.0) | 11.0 (10.0–50.0) | 0.268 | 15.0 (5.10–38.0) | 19.5 (11.0–31.0) | 0.557 |
| Creatinine | 0.472 ± 0.112 | 0.670 ± 0.171 | 0.024 * | 0.482 ± 0.156 | 0.480 ± 0.175 | 0.977 | 0.451 ± 0.136 | 0.565 ± 0.139 | 0.162 |
| DNMT3B Genotype | Methylated | Partially Methylated (n = 50) | Unmethylated (n = 13) | ||||||
| --- | TT + TC (n = 39) | CC (n = 11) | p | TT + TC (n = 8) | CC (n = 5) | p | |||
| Hemoglobin | 9.230 ± 2.092 | 10.036 ± 1.871 | 0.253 | 9.550 ± 2.7449 | 9.600 ± 2.440 | 0.974 | |||
| Leukocytes | 3000 (100–42,400) | 3200 (1300–6600) | 0.882 | 2000 (300–7700) | 5000 (100–17,500) | 0.509 | |||
| Platelets | 154,000 (23,000–561,000) | 175,000 (36,000–415,000) | 0.267 | 98,500 (27,000–296,000) | 110,000 (35,000–533,000) | 0.524 | |||
| Urea | 19.0 (5.10–69.0) | 16.0 (10.0–35.0) | 0.766 | 19.0 (9.00–45.0) | 16.0 (11.0–61.0) | 0.713 | |||
| Creatinine | 0.504 ± 0.170 | 0.482 ± 0.107 | 0.688 | 0.395 ± 0.0961 | 0.576 ± 0.107 | 0.009 * | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Souza, B.F.; Viana Filho, J.M.C.; de Queiroz Neto, J.N.; Coêlho, M.d.C.; Valença, A.M.G.; Persuhn, D.C.; de Oliveira, N.F.P. DNA Methyltransferase Genes Are Associated with Oral Mucositis and Creatinine Levels in Oncopediatric Patients. Genes 2023, 14, 1136. https://doi.org/10.3390/genes14061136
de Souza BF, Viana Filho JMC, de Queiroz Neto JN, Coêlho MdC, Valença AMG, Persuhn DC, de Oliveira NFP. DNA Methyltransferase Genes Are Associated with Oral Mucositis and Creatinine Levels in Oncopediatric Patients. Genes. 2023; 14(6):1136. https://doi.org/10.3390/genes14061136
Chicago/Turabian Stylede Souza, Beatriz Fernandes, José Maria Chagas Viana Filho, José Nunes de Queiroz Neto, Marina de Castro Coêlho, Ana Maria Gondim Valença, Darlene Camati Persuhn, and Naila Francis Paulo de Oliveira. 2023. "DNA Methyltransferase Genes Are Associated with Oral Mucositis and Creatinine Levels in Oncopediatric Patients" Genes 14, no. 6: 1136. https://doi.org/10.3390/genes14061136