
Coronin 1C, Regulated by Multiple microRNAs, Facilitates Cancer Cell Aggressiveness in Pancreatic Ductal Adenocarcinoma

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
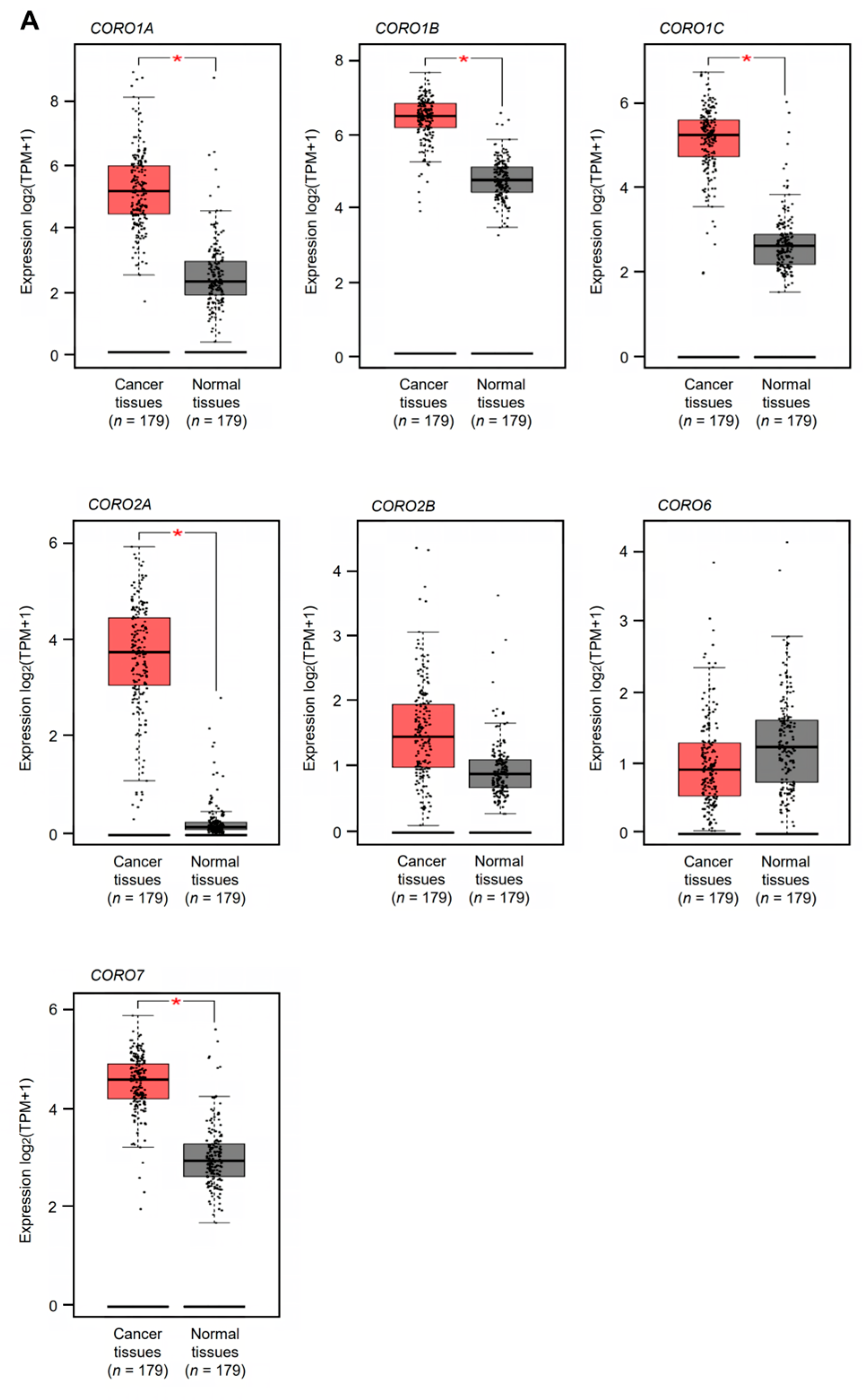
2.1. Analysis of the Expression and Clinical Significance of Coronin Genes in PDAC
2.2. Gene Set Enrichment Analysis (GSEA)
2.3. Human PDAC Cell Lines
2.4. Transfection of Small Interfering RNAs (siRNAs) and miRNAs into PDAC Cells and Quantitative Reverse-Transcription PCR (qRT-PCR)
2.5. Functional Assays (Cell Proliferation, Migration, and Invasion) in PDAC Cells
2.6. Identification of the miRNAs Regulating CORO1C Expression in PDAC Cells
2.7. Western Blotting
2.8. Statistical Analysis
3. Results
3.1. Expression and Clinical Significance of Coronin Family Members in Patients with PDAC
3.2. Molecular Pathways Associated with High CORO1C Expression in PDAC Cells
3.3. Effects of CORO1C Knockdown on PDAC Cell Proliferation, Migration, and Invasion
3.4. Identification of miRNAs That Regulate CORO1C Expression in PDAC Cells
3.5. Tumor-Suppressive Function of miR-26a-5p and miR-29c-3p in PDAC Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA A Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- McGuigan, A.; Kelly, P.; Turkington, R.C.; Jones, C.; Coleman, H.G.; McCain, R.S. Pancreatic cancer: A review of clinical diagnosis, epidemiology, treatment and outcomes. World J. Gastroenterol. 2018, 24, 4846–4861. [Google Scholar] [CrossRef] [PubMed]
- Mizrahi, J.D.; Surana, R.; Valle, J.W.; Shroff, R.T. Pancreatic cancer. Lancet 2020, 395, 2008–2020. [Google Scholar] [CrossRef] [PubMed]
- Cancer Statistics. Cancer Information Service, National Cancer Center, Japan (Vital Statistics of Japan, Ministry of Health, Labour and Welfare). Available online: https://ganjoho.jp/reg_stat/statistics/data/dl/index.html#mortality (accessed on 7 March 2023).
- Park, W.; Chawla, A.; O’Reilly, E.M. Pancreatic cancer: A review. JAMA 2021, 326, 851–862. [Google Scholar] [CrossRef]
- Rémond, M.S.; Pellat, A.; Brezault, C.; Dhooge, M.; Coriat, R. Are targeted therapies or immunotherapies effective in metastatic pancreatic adenocarcinoma? ESMO Open 2022, 7, 100638. [Google Scholar] [CrossRef]
- Qian, Y.; Gong, Y.; Fan, Z.; Luo, G.; Huang, Q.; Deng, S.; Cheng, H.; Jin, K.; Ni, Q.; Yu, X.; et al. Molecular alterations and targeted therapy in pancreatic ductal adenocarcinoma. J. Hematol. Oncol. 2020, 13, 130. [Google Scholar] [CrossRef]
- Cicenas, J.; Kvederaviciute, K.; Meskinyte, I.; Meskinyte-Kausiliene, E.; Skeberdyte, A.; Cicenas, J. KRAS, TP53, CDKN2A, SMAD4, BRCA1, and BRCA2 mutations in pancreatic cancer. Cancers 2017, 9, 42. [Google Scholar] [CrossRef]
- Pishvaian, M.J.; Blais, E.M.; Brody, J.R.; Lyons, E.; DeArbeloa, P.; Hendifar, A.; Mikhail, S.; Chung, V.; Sahai, V.; Sohal, D.P.S.; et al. Overall survival in patients with pancreatic cancer receiving matched therapies following molecular profiling: A retrospective analysis of the Know Your Tumor registry trial. Lancet Oncol. 2020, 21, 508–518. [Google Scholar] [CrossRef]
- Tempero, M.A. NCCN guidelines updates: Pancreatic cancer. J. Natl. Compr. Cancer Netw. JNCCN 2019, 17, 603–605. [Google Scholar]
- Jones, R.P.; Psarelli, E.E.; Jackson, R.; Ghaneh, P.; Halloran, C.M.; Palmer, D.H.; Campbell, F.; Valle, J.W.; Faluyi, O.; O’Reilly, D.A.; et al. Patterns of recurrence after resection of pancreatic ductal adenocarcinoma: A secondary analysis of the ESPAC-4 randomized adjuvant chemotherapy trial. JAMA Surg. 2019, 154, 1038–1048. [Google Scholar] [CrossRef]
- Winkler, J.; Abisoye-Ogunniyan, A.; Metcalf, K.J.; Werb, Z. Concepts of extracellular matrix remodelling in tumour progression and metastasis. Nat. Commun. 2020, 11, 5120. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Li, Y.; Ge, H.; Ghadban, T.; Reeh, M.; Güngör, C. The extracellular matrix: A key accomplice of cancer stem cell migration, metastasis formation, and drug resistance in PDAC. Cancers 2022, 14, 3998. [Google Scholar] [CrossRef]
- Gross, S.R. Actin binding proteins: Their ups and downs in metastatic life. Cell Adhes. Migr. 2013, 7, 199–213. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.T.; Creed, S.J.; Bear, J.E. Unraveling the enigma: Progress towards understanding the coronin family of actin regulators. Trends Cell Biol. 2011, 21, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Uetrecht, A.C.; Bear, J.E. Coronins: The return of the crown. Trends Cell Biol. 2006, 16, 421–426. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef]
- Kong, Y.W.; Ferland-McCollough, D.; Jackson, T.J.; Bushell, M. MicroRNAs in cancer management. Lancet Oncol. 2012, 13, e249–e258. [Google Scholar] [CrossRef]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef]
- Mootha, V.K.; Lindgren, C.M.; Eriksson, K.F.; Subramanian, A.; Sihag, S.; Lehar, J.; Puigserver, P.; Carlsson, E.; Ridderstråle, M.; Laurila, E.; et al. PGC-1alpha-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 2003, 34, 267–273. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Liberzon, A.; Subramanian, A.; Pinchback, R.; Thorvaldsdóttir, H.; Tamayo, P.; Mesirov, J.P. Molecular signatures database (MSigDB) 3.0. Bioinformatics 2011, 27, 1739–1740. [Google Scholar] [CrossRef] [PubMed]
- Idichi, T.; Seki, N.; Kurahara, H.; Fukuhisa, H.; Toda, H.; Shimonosono, M.; Yamada, Y.; Arai, T.; Kita, Y.; Kijima, Y.; et al. Involvement of anti-tumor miR-124-3p and its targets in the pathogenesis of pancreatic ductal adenocarcinoma: Direct regulation of ITGA3 and ITGB1 by miR-124-3p. Oncotarget 2018, 9, 28849–28865. [Google Scholar] [CrossRef] [PubMed]
- Hozaka, Y.; Kita, Y.; Yasudome, R.; Tanaka, T.; Wada, M.; Idichi, T.; Tanabe, K.; Asai, S.; Moriya, S.; Toda, H.; et al. RNA-sequencing based microRNA expression signature of colorectal cancer: The impact of oncogenic targets regulated by miR-490-3p. Int. J. Mol. Sci. 2021, 22, 9876. [Google Scholar] [CrossRef] [PubMed]
- Yasudome, R.; Seki, N.; Asai, S.; Goto, Y.; Kita, Y.; Hozaka, Y.; Wada, M.; Tanabe, K.; Idichi, T.; Mori, S.; et al. Molecular pathogenesis of colorectal cancer: Impact of oncogenic targets regulated by tumor suppressive miR-139-3p. Int. J. Mol. Sci. 2022, 23, 11616. [Google Scholar] [CrossRef] [PubMed]
- Yonemori, K.; Seki, N.; Idichi, T.; Kurahara, H.; Osako, Y.; Koshizuka, K.; Arai, T.; Okato, A.; Kita, Y.; Arigami, T.; et al. The microRNA expression signature of pancreatic ductal adenocarcinoma by RNA sequencing: Anti-tumour functions of the microRNA-216 cluster. Oncotarget 2017, 8, 70097–70115. [Google Scholar] [CrossRef] [PubMed]
- Fukuhisa, H.; Seki, N.; Idichi, T.; Kurahara, H.; Yamada, Y.; Toda, H.; Kita, Y.; Kawasaki, Y.; Tanoue, K.; Mataki, Y.; et al. Gene regulation by antitumor miR-130b-5p in pancreatic ductal adenocarcinoma: The clinical significance of oncogenic EPS8. J. Hum. Genet. 2019, 64, 521–534. [Google Scholar] [CrossRef]
- Idichi, T.; Seki, N.; Kurahara, H.; Fukuhisa, H.; Toda, H.; Shimonosono, M.; Okato, A.; Arai, T.; Kita, Y.; Mataki, Y.; et al. Molecular pathogenesis of pancreatic ductal adenocarcinoma: Impact of passenger strand of pre-miR-148a on gene regulation. Cancer Sci. 2018, 109, 2013–2026. [Google Scholar] [CrossRef]
- Idichi, T.; Seki, N.; Kurahara, H.; Yonemori, K.; Osako, Y.; Arai, T.; Okato, A.; Kita, Y.; Arigami, T.; Mataki, Y.; et al. Regulation of actin-binding protein ANLN by antitumor miR-217 inhibits cancer cell aggressiveness in pancreatic ductal adenocarcinoma. Oncotarget 2017, 8, 53180–53193. [Google Scholar] [CrossRef]
- Thal, D.; Xavier, C.P.; Rosentreter, A.; Linder, S.; Friedrichs, B.; Waha, A.; Pietsch, T.; Stumpf, M.; Noegel, A.; Clemen, C. Expression of coronin-3 (coronin-1C) in diffuse gliomas is related to malignancy. J. Pathol. 2008, 214, 415–424. [Google Scholar] [CrossRef]
- Han, S.; Ding, X.; Wang, S.; Xu, L.; Li, W.; Sun, W. MiR-133a-3p regulates hepatocellular carcinoma progression through targeting CORO1C. Cancer Manag. Res. 2020, 12, 8685–8693. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tsouko, E.; Jonsson, P.; Bergh, J.; Hartman, J.; Aydogdu, E.; Williams, C. MiR-206 inhibits cell migration through direct targeting of the actin-binding protein coronin 1C in triple-negative breast cancer. Mol. Oncol. 2014, 8, 1690–1702. [Google Scholar] [CrossRef]
- Lim, J.P.; Shyamasundar, S.; Gunaratne, J.; Scully, O.J.; Matsumoto, K.; Bay, B.H. YBX1 gene silencing inhibits migratory and invasive potential via CORO1C in breast cancer in vitro. BMC Cancer 2017, 17, 201. [Google Scholar] [CrossRef] [PubMed]
- Castagnino, A.; Castro-Castro, A.; Irondelle, M.; Guichard, A.; Lodillinsky, C.; Fuhrmann, L.; Vacher, S.; Aguera-Gonzalez, S.; Zagryazhskaya-Masson, A.; Romao, M.; et al. Coronin 1C promotes triple-negative breast cancer invasiveness through regulation of MT1-MMP traffic and invadopodia function. Oncogene 2018, 37, 6425–6441. [Google Scholar] [CrossRef]
- Fujii, K.; Miyata, Y.; Takahashi, I.; Koizumi, H.; Saji, H.; Hoshikawa, M.; Takagi, M.; Nishimura, T.; Nakamura, H. Differential proteomic analysis between small cell lung carcinoma (SCLC) and pulmonary carcinoid tumors reveals molecular signatures for malignancy in lung cancer. Proteom. Clin. Appl. 2018, 12, e1800015. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Wang, X.; Wu, Z.; Tan, S.; Zhu, T.; Ding, K. CORO1C expression is associated with poor survival rates in gastric cancer and promotes metastasis in vitro. FEBS Open Bio 2019, 9, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Jia, L.; Sun, Y.; Li, C.; Zhang, L.; Wang, X.; Chen, H. CORO1C is associated with poor prognosis and promotes metastasis through PI3K/AKT pathway in colorectal cancer. Front. Mol. Biosci 2021, 8, 682594. [Google Scholar] [CrossRef]
- Greco, L.; Rubbino, F.; Laghi, L. Epithelial to mesenchymal transition as mechanism of progression of pancreatic cancer: From mice to men. Cancers 2022, 14, 5797. [Google Scholar] [CrossRef]
- Joshi, V.B.; Gutierrez Ruiz, O.L.; Razidlo, G.L. The cell biology of metastatic invasion in pancreatic cancer: Updates and Mechanistic Insights. Cancers 2023, 15, 2169. [Google Scholar] [CrossRef]
- Zheng, X.; Carstens, J.L.; Kim, J.; Scheible, M.; Kaye, J.; Sugimoto, H.; Wu, C.C.; LeBleu, V.S.; Kalluri, R. Epithelial-to-mesenchymal transition is dispensable for metastasis but induces chemoresistance in pancreatic cancer. Nature 2015, 527, 525–530. [Google Scholar] [CrossRef]
- Alzahrani, A.S. PI3K/Akt/mTOR inhibitors in cancer: At the bench and bedside. Semin. Cancer Biol. 2019, 59, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, M.; Moosavi, F.; Martini, M.; Giovannetti, E.; Firuzi, O. Prospects of targeting PI3K/AKT/mTOR pathway in pancreatic cancer. Crit. Rev. Oncol./Hematol. 2022, 176, 103749. [Google Scholar] [CrossRef] [PubMed]
- Asati, V.; Mahapatra, D.K.; Bharti, S.K. PI3K/Akt/mTOR and Ras/Raf/MEK/ERK signaling pathways inhibitors as anticancer agents: Structural and pharmacological perspectives. Eur. J. Med. Chem. 2016, 109, 314–341. [Google Scholar] [CrossRef] [PubMed]
- Stanciu, S.; Ionita-Radu, F.; Stefani, C.; Miricescu, D.; Stanescu, S., II; Greabu, M.; Ripszky Totan, A.; Jinga, M. Targeting PI3K/AKT/mTOR signaling pathway in pancreatic cancer: From molecular to clinical aspects. Int. J. Mol. Sci. 2022, 23, 10132. [Google Scholar] [CrossRef]
- Yonemori, K.; Seki, N.; Kurahara, H.; Osako, Y.; Idichi, T.; Arai, T.; Koshizuka, K.; Kita, Y.; Maemura, K.; Natsugoe, S. ZFP36L2 promotes cancer cell aggressiveness and is regulated by antitumor microRNA-375 in pancreatic ductal adenocarcinoma. Cancer Sci. 2017, 108, 124–135. [Google Scholar] [CrossRef]
- Khalid, M.; Idichi, T.; Seki, N.; Wada, M.; Yamada, Y.; Fukuhisa, H.; Toda, H.; Kita, Y.; Kawasaki, Y.; Tanoue, K.; et al. Gene Regulation by antitumor miR-204-5p in pancreatic ductal adenocarcinoma: The clinical significance of direct RACGAP1 regulation. Cancers 2019, 11, 327. [Google Scholar] [CrossRef]
- Shimomura, H.; Okada, R.; Tanaka, T.; Hozaka, Y.; Wada, M.; Moriya, S.; Idichi, T.; Kita, Y.; Kurahara, H.; Ohtsuka, T.; et al. Role of miR-30a-3p regulation of oncogenic targets in pancreatic ductal adenocarcinoma pathogenesis. Int. J. Mol. Sci. 2020, 21, 6459. [Google Scholar] [CrossRef]
- Tanaka, T.; Okada, R.; Hozaka, Y.; Wada, M.; Moriya, S.; Satake, S.; Idichi, T.; Kurahara, H.; Ohtsuka, T.; Seki, N. Molecular pathogenesis of pancreatic ductal adenocarcinoma: Impact of miR-30c-5p and miR-30c-2-3p regulation on oncogenic genes. Cancers 2020, 12, 2731. [Google Scholar] [CrossRef]
- Nepal, P.; Hozaka, Y.; Tanaka, T.; Wada, M.; Asai, S.; Minemura, C.; Idichi, T.; Arigami, T.; Kurahara, H.; Seki, N.; et al. Impact of oncogenic targets controlled by tumor-suppressive miR-30a-5p in pancreatic ductal adenocarcinoma. Anticancer Res. 2021, 41, 4821–4836. [Google Scholar] [CrossRef]
- Deng, J.; He, M.; Chen, L.; Chen, C.; Zheng, J.; Cai, Z. The loss of miR-26a-mediated post-transcriptional regulation of cyclin E2 in pancreatic cancer cell proliferation and decreased patient survival. PLoS ONE 2013, 8, e76450. [Google Scholar] [CrossRef]
- Wang, L.; Li, M.; Chen, F. MicroRNA-26a represses pancreatic cancer cell malignant behaviors by targeting E2F7. Discov. Oncol. 2021, 12, 55. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, T.; Xue, W.; Fang, Y.; Chen, X.; Xu, L.; Zhang, L.; Guan, K.; Pan, J.; Zheng, L.; et al. ARNTL2 promotes pancreatic ductal adenocarcinoma progression through TGF/BETA pathway and is regulated by miR-26a-5p. Cell Death Dis. 2020, 11, 692. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Hu, C.; Cao, H.; Wu, X.; Wang, R.; Lu, H.; Li, H.; Chen, H. MicroRNA-29c Increases the chemosensitivity of pancreatic cancer cells by inhibiting USP22 mediated autophagy. Cell Physiol. Biochem. 2018, 47, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Hu, J.; Sun, W.; Li, S.; Deng, S.; Li, M. MiR-29c inhibits cell growth, invasion, and migration of pancreatic cancer by targeting ITGB1. OncoTargets Ther. 2016, 9, 99–109. [Google Scholar] [CrossRef]
- Si, H.; Zhang, N.; Shi, C.; Luo, Z.; Hou, S. Tumor-suppressive miR-29c binds to MAPK1 inhibiting the ERK/MAPK pathway in pancreatic cancer. Clin. Transl. Oncol. 2022, 25, 803–816. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Kawagoe, K.; Wada, M.; Idichi, T.; Okada, R.; Yamada, Y.; Moriya, S.; Okubo, K.; Matsushita, D.; Arigami, T.; Kurahara, H.; et al. Regulation of aberrantly expressed SERPINH1 by antitumor miR-148a-5p inhibits cancer cell aggressiveness in gastric cancer. J. Hum. Genet. 2020, 65, 647–656. [Google Scholar] [CrossRef]
- Wada, M.; Goto, Y.; Tanaka, T.; Okada, R.; Moriya, S.; Idichi, T.; Noda, M.; Sasaki, K.; Kita, Y.; Kurahara, H.; et al. RNA sequencing-based microRNA expression signature in esophageal squamous cell carcinoma: Oncogenic targets by antitumor miR-143-5p and miR-143-3p regulation. J. Hum. Genet. 2020, 65, 1019–1034. [Google Scholar] [CrossRef]
- Shinden, Y.; Hirashima, T.; Nohata, N.; Toda, H.; Okada, R.; Asai, S.; Tanaka, T.; Hozaka, Y.; Ohtsuka, T.; Kijima, Y.; et al. Molecular pathogenesis of breast cancer: Impact of miR-99a-5p and miR-99a-3p regulation on oncogenic genes. J. Hum. Genet. 2021, 66, 519–534. [Google Scholar] [CrossRef]
- Zhou, S.; Zhu, C.; Pang, Q.; Liu, H.C. MicroRNA-217: A regulator of human cancer. Biomed. Pharm. 2021, 133, 110943. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | miRbase Accession | Chromosome | FC (log2) | p Value | FDR |
|---|---|---|---|---|---|
| hsa-miR-217 | MIMAT0000274 | 2 | −3.1333 | 0.003394591 | 0.460235049 |
| hsa-miR-216a-3p | MIMAT0022844 | 2 | −2.5792 | 0.000762794 | 0.172184571 |
| hsa-miR-129-1-3p | MIMAT0004548 | 7 | −2.4045 | 0.001131503 | 0.208196466 |
| hsa-miR-148a-5p | MIMAT0004549 | 7 | −2.3043 | 0.000857068 | 0.172184571 |
| hsa-miR-211-5p | MIMAT0000268 | 15 | −2.1027 | 0.008394937 | 0.554496371 |
| hsa-miR-129-2-3p | MIMAT0004605 | 11 | −2.0092 | 0.008992112 | 0.564967844 |
| hsa-miR-2114-3p | MIMAT0011157 | X | −1.9620 | 0.025664949 | 0.920037824 |
| hsa-miR-7-2-3p | MIMAT0004554 | 15 | −1.9294 | 0.118440035 | 1 |
| hsa-miR-4780 | MIMAT0019939 | 2 | −1.7904 | 0.063787284 | 1 |
| hsa-miR-129-5p | MIMAT0000242 | 7 | −1.6670 | 0.063243326 | 1 |
| hsa-miR-204-5p | MIMAT0000265 | 9 | −1.6320 | 0.028968095 | 0.969114438 |
| hsa-miR-130b-5p | MIMAT0004680 | 22 | −1.5926 | 0.006911516 | 0.502959718 |
| hsa-miR-19a-3p | MIMAT0000073 | 13 | −1.3565 | 0.072163518 | 1 |
| hsa-miR-7855-5p | MIMAT0030430 | 14 | −1.3371 | 0.219871323 | 1 |
| hsa-miR-135a-5p | MIMAT0000428 | 3 | −1.3206 | 0.049728049 | 1 |
| hsa-miR-4507 | MIMAT0019044 | 14 | −1.3176 | 0.284165056 | 1 |
| hsa-miR-16-1-3p | MIMAT0004489 | 13 | −1.3047 | 0.297303611 | 1 |
| hsa-miR-4732-5p | MIMAT0019855 | 17 | −1.3026 | 0.178680845 | 1 |
| hsa-miR-30c-2-3p | MIMAT0004550 | 6 | −1.2993 | 0.006492318 | 0.502959718 |
| hsa-miR-576-5p | MIMAT0003241 | 4 | −1.2962 | 0.036429558 | 1 |
| hsa-miR-3938 | MIMAT0018353 | 3 | −1.2899 | 0.295549865 | 1 |
| hsa-miR-5589-3p | MIMAT0022298 | 19 | −1.2730 | 0.305462044 | 1 |
| hsa-miR-323a-3p | MIMAT0000755 | 14 | −1.2519 | 0.067319367 | 1 |
| hsa-miR-5189-5p | MIMAT0021120 | 16 | −1.1895 | 0.302501869 | 1 |
| hsa-miR-3133 | MIMAT0014998 | 2 | −1.1883 | 0.346269763 | 1 |
| hsa-miR-9-3p | MIMAT0000442 | 1 | −1.1722 | 0.265080141 | 1 |
| hsa-miR-3178 | MIMAT0015055 | 16 | −1.1722 | 0.265080141 | 1 |
| hsa-miR-494-3p | MIMAT0002816 | 14 | −1.1329 | 0.069170156 | 1 |
| hsa-miR-382-3p | MIMAT0022697 | 14 | −1.1112 | 0.186877213 | 1 |
| hsa-miR-19b-3p | MIMAT0000074 | 13 | −1.0940 | 0.073080409 | 1 |
| hsa-miR-4772-3p | MIMAT0019927 | 2 | −1.0569 | 0.343904631 | 1 |
| hsa-miR-5193 | MIMAT0021124 | 3 | −1.0533 | 0.289583569 | 1 |
| hsa-miR-29c-3p | MIMAT0000681 | 1 | −1.0248 | 0.041438418 | 1 |
| hsa-miR-7152-5p | MIMAT0028214 | 10 | −1.0203 | 0.309763874 | 1 |
| hsa-miR-26a-5p | MIMAT0000082 | 3 | −1.0153 | 0.005038089 | 0.502959718 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fukuda, K.; Seki, N.; Yasudome, R.; Mitsueda, R.; Asai, S.; Kato, M.; Idichi, T.; Kurahara, H.; Ohtsuka, T. Coronin 1C, Regulated by Multiple microRNAs, Facilitates Cancer Cell Aggressiveness in Pancreatic Ductal Adenocarcinoma. Genes 2023, 14, 995. https://doi.org/10.3390/genes14050995
Fukuda K, Seki N, Yasudome R, Mitsueda R, Asai S, Kato M, Idichi T, Kurahara H, Ohtsuka T. Coronin 1C, Regulated by Multiple microRNAs, Facilitates Cancer Cell Aggressiveness in Pancreatic Ductal Adenocarcinoma. Genes. 2023; 14(5):995. https://doi.org/10.3390/genes14050995
Chicago/Turabian StyleFukuda, Kosuke, Naohiko Seki, Ryutaro Yasudome, Reiko Mitsueda, Shunichi Asai, Mayuko Kato, Tetsuya Idichi, Hiroshi Kurahara, and Takao Ohtsuka. 2023. "Coronin 1C, Regulated by Multiple microRNAs, Facilitates Cancer Cell Aggressiveness in Pancreatic Ductal Adenocarcinoma" Genes 14, no. 5: 995. https://doi.org/10.3390/genes14050995