
TNFα Causes a Shift in Gene Expression of TNFRSF1A and TNFRSF1B Isoforms

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Signal Peptide-Positive Protein Coding Isoform Selection via InterPro
2.2. Primer Design
2.3. Cell Line Culturing with TNFα
2.4. Total RNA Extraction
2.5. Reverse Transcription
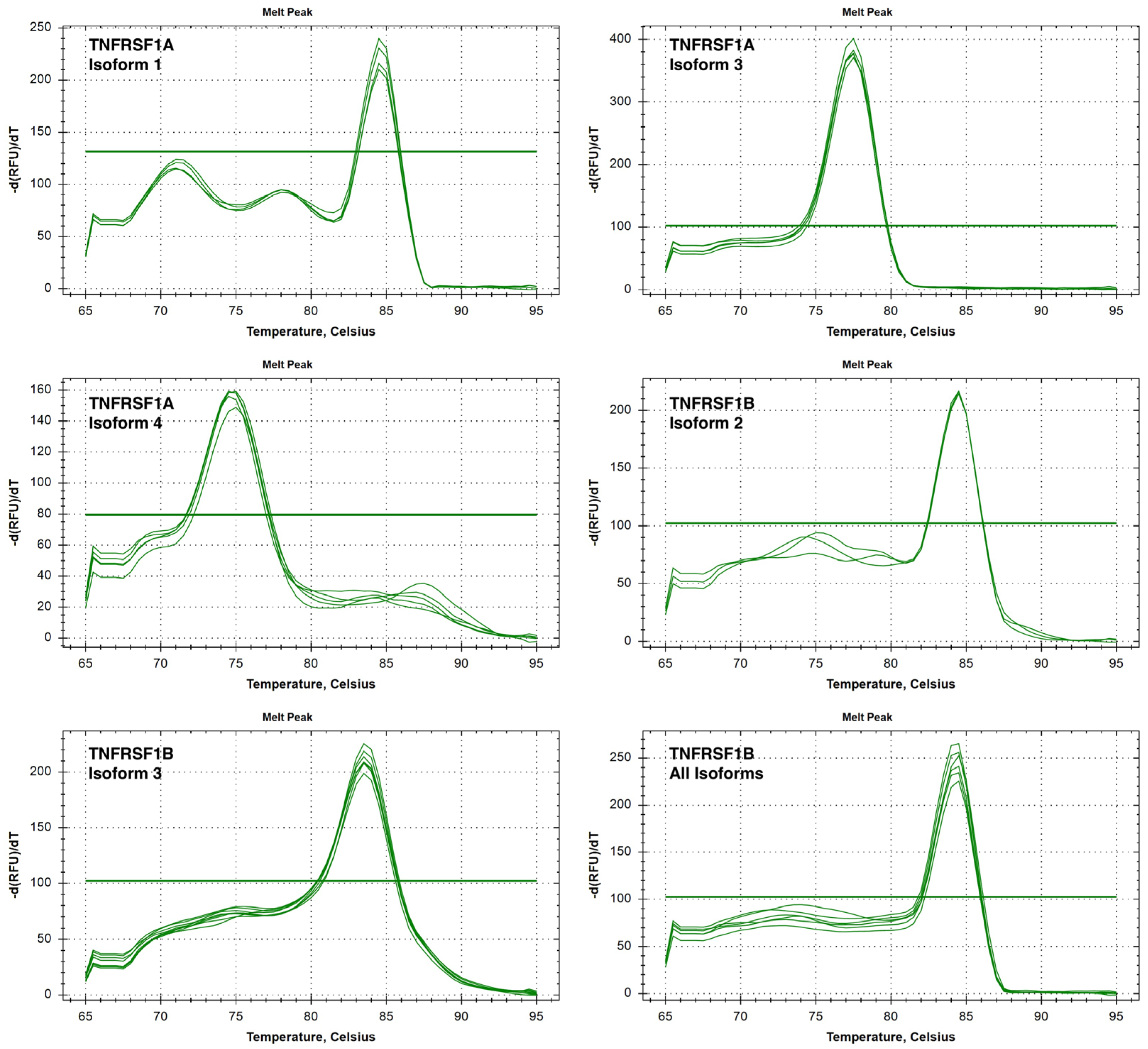
2.6. PCR and Melt Curve Analysis
2.7. Relative Isoform Expression Analysis
2.8. Differential Isoform Expression Testing
3. Results
3.1. Analysis of Isoforms of TNFRSF1A and TNFRSF1B Genes for the Presence of Signal Peptide in Their Amino Acid Sequence
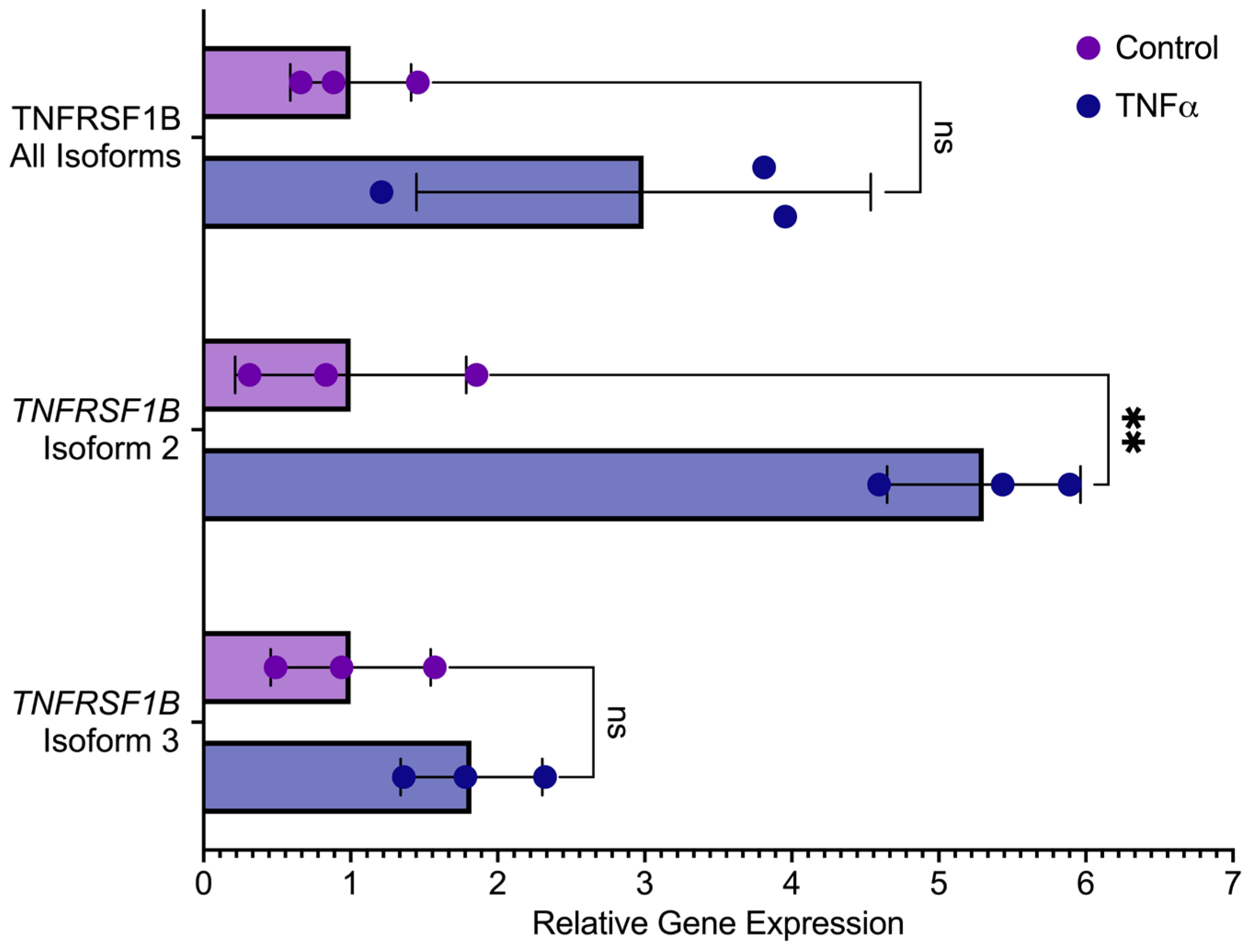
3.2. Study of TNFRSF1A and TNFRSF1B Gene Isoform Expression after TNFα Exposure
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pan, Q.; Shai, O.; Lee, L.J.; Frey, B.J.; Blencowe, B.J. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat. Genet. 2008, 40, 1413–1415. [Google Scholar] [CrossRef]
- Jiang, W.; Chen, L. Alternative splicing: Human disease and quantitative analysis from high-throughput sequencing. Comput. Struct. Biotechnol. J. 2021, 19, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Yatsenko, O.P.; Filipenko, M.L.; Khrapov, E.A.; Voronina, E.N.; Kozlov, V.A.; Sennikov, S.V. Alternative splicing of mRNA of mouse interleukin-4 and interleukin-6. Cytokine 2004, 28, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Rittore, C.; Méchin, D.; Sanchez, E.; Marinèche, L.; Ea, V.; Soler, S.; Vereecke, M.; Mallavialle, A.; Richard, E.; Duroux-Richard, I.; et al. TNFR1-d2 carrying the p.(Thr79Met) pathogenic variant is a potential novel actor of TNFα/TNFR1 signalling regulation in the pathophysiology of TRAPS. Sci. Rep. 2021, 11, 4172. [Google Scholar] [CrossRef] [PubMed]
- Alshevskaya, A.; Zhukova, J.; Kireev, F.; Lopatnikova, J.; Evsegneeva, I.; Demina, D.; Nepomniashchikch, V.; Gladkikh, V.; Karaulov, A.; Sennikov, S. Redistribution of TNF Receptor 1 and 2 Expression on Immune Cells in Patients with Bronchial Asthma. Cells 2022, 11, 1736. [Google Scholar] [CrossRef] [PubMed]
- Alshevskaya, A.; Lopatnikova, J.; Zhukova, J.; Chumasova, O.; Shkaruba, N.; Sizikov, A.; Evsegneeva, I.; Gladkikh, V.; Karaulov, A.; Sennikov, S.V. Co-Expression Profile of TNF Membrane-Bound Receptors Type 1 and 2 in Rheumatoid Arthritis on Immunocompetent Cells Subsets. Int. J. Mol. Sci. 2019, 21, 288. [Google Scholar] [CrossRef] [PubMed]
- Alshevskaya, A.; Lopatnikova, J.; Zhukova, J.; Chumasova, O.; Shkaruba, N.; Sizikov, A.; Evsegneeva, I.; Demina, D.; Nepomniashchikch, V.; Karaulov, A.; et al. The Influence of Severity and Disease Duration on TNF Receptors’ Redistribution in Asthma and Rheumatoid Arthritis. Cells 2022, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- Lainez, B.; Fernandez-Real, J.M.; Romero, X.; Esplugues, E.; Cañete, J.D.; Ricart, W.; Engel, P. Identification and characterization of a novel spliced variant that encodes human soluble tumor necrosis factor receptor 2. Int. Immunol. 2004, 16, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Cañete, J.D.; Albaladejo, C.; Hernández, M.V.; Laínez, B.; Pinto, J.A.; Ramírez, J.; López-Armada, M.J.; Rodríguez-Cros, J.R.; Engel, P.; Blanco, F.J.; et al. Clinical significance of high levels of soluble tumour necrosis factor-α receptor-2 produced by alternative splicing in rheumatoid arthritis: A longitudinal prospective cohort study. Rheumatology 2011, 50, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Alshevskaya, A.A.; Belomestnova, I.; Lopatnikova, J.A.; Zhukova, J.; Evsegneeva, I.; Koneva, O.; Karaulov, A.V.; Sennikov, S.V. Co-Expression of Membrane-Bound Tumor Necrosis Factor-Alpha Receptor Types 1 and 2 by Tumor Cell Lines. Int. Arch. Allergy Immunol. 2020, 181, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Quevillon, E.; Silventoinen, V.; Pillai, S.; Harte, N.; Mulder, N.; Apweiler, R.; Lopez, R. InterProScan: Protein domains identifier. Nucleic Acids Res. 2005, 33, W116–W120. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.C.; Nadeau, K.; Abbasi, M.; Lachance, C.; Nguyen, M.; Fenrich, J. The ultimate qPCR experiment: Producing publication quality, reproducible data the first time. Trends Biotechnol. 2019, 37, 761–774. [Google Scholar] [CrossRef] [PubMed]
- Franco, H.; Nagari, A.; Kraus, W.L. TNFα signaling exposes latent estrogen receptor binding sites to alter the breast cancer cell transcriptome. Mol. Cell 2015, 58, 21–34. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isoform | Primer | Sequence | Tm °C | CG% | nt | Melt C | NCBI Accession Number |
|---|---|---|---|---|---|---|---|
| 1 | TNFRSF1A1F | GGACAGGGAGAAGAGAGATAGTGTG | 65.6 | 52.0 | 25 | 84.6 | NM_001065.4 |
| TNFRSF1A1R | GGCTTTTCTTACAGTTACTACAGGAGACAC | 66.1 | 43.3 | 30 | NM_001065.4 | ||
| 3 | TNFRSF1A3F | CGATCTCGGCTCACTGCAAC | 67.2 | 60.0 | 20 | 77.4 | NM_001346092.2 |
| TNFRSF1A3R | CACTGTGGTGCCTGAGTCCT | 64.3 | 60.0 | 20 | NM_001346092.2 | ||
| 4 | TNFRSF1A4F | CCAGGAGAAACAGAACACCGTG | 66.7 | 54.5 | 22 | 74.7 | NR_144351.2 |
| TNFRSF1A4R | GGTGCTTACTACAGGAGACACACTCG | 66.8 | 53.8 | 26 | NR_144351.2 |
| Isoform | Primer | Sequence | Tm °C | CG% | nt | Melt C | NCBI Accession Number |
|---|---|---|---|---|---|---|---|
| 2 | TNFRSF1B2F | CTATGACCAGACAGCTCAGATGTGC | 67.0 | 52.0 | 25 | 84.3 | XM_047429422.1 |
| TNFRSF1B2R | GACATTGTCTTCTCCTGTCTCCCA | 66.5 | 50.0 | 24 | |||
| 3 | TNFRSF1B3F | GCATTTACACCCTACGCCCC | 66.2 | 60.0 | 20 | 84.4 | XM_047429423.1 |
| TNFRSF1B3R | CAGGGGAAGAATCTGCTGAGC | 66.2 | 57.1 | 21 | |||
| All Isoforms | TNFRSF1BCONSF | GGTTTCTGCCCACATTGGAC | 65.4 | 55.0 | 20 | 84.7 | NM_001066.3 |
| TNFRSF1BCONSR | CAGTGCAAACTTTCATTGTCTTGG | 65.5 | 41.7 | 24 |
| Gene | Primer | Sequence | Tm C | CG% | nt |
|---|---|---|---|---|---|
| GAPDH | F | GTCTCCTCTGACTTCAACAGCG | 65.7 | 54.5 | 22 |
| R | ACCACCCTGTTGCTGTAGCC | 66.7 | 60.0 | 20 | |
| HPRT | F | CATTATGCTGAGGATTTGGAAAGG | 66.5 | 41.7 | 24 |
| R | CTTGAGCACACAGAGGGCTACA | 67.0 | 54.5 | 22 | |
| TUBB5 | F | CTGGACCGCATCTCTGTGTACT | 65.8 | 54.5 | 22 |
| R | CAAAAGGACCTGAGCGAACA | 65.6 | 50.0 | 20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perik-Zavodskaia, O.; Zhukova, J.; Perik-Zavodskii, R.; Alshevskaya, A.; Lopatnikova, J.; Sennikov, S. TNFα Causes a Shift in Gene Expression of TNFRSF1A and TNFRSF1B Isoforms. Genes 2023, 14, 1074. https://doi.org/10.3390/genes14051074
Perik-Zavodskaia O, Zhukova J, Perik-Zavodskii R, Alshevskaya A, Lopatnikova J, Sennikov S. TNFα Causes a Shift in Gene Expression of TNFRSF1A and TNFRSF1B Isoforms. Genes. 2023; 14(5):1074. https://doi.org/10.3390/genes14051074
Chicago/Turabian StylePerik-Zavodskaia, Olga, Julia Zhukova, Roman Perik-Zavodskii, Alina Alshevskaya, Julia Lopatnikova, and Sergey Sennikov. 2023. "TNFα Causes a Shift in Gene Expression of TNFRSF1A and TNFRSF1B Isoforms" Genes 14, no. 5: 1074. https://doi.org/10.3390/genes14051074