
The GJB2 (Cx26) Gene Variants in Patients with Hearing Impairment in the Baikal Lake Region (Russia)

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Clinical and Audiological Examinations
2.3. GJB2 Gene Sequence Analysis
2.4. Screening of Large DFNB1 Deletions
2.5. In Silico Pathogenicity Analysis
2.6. Haplotypes Analysis
2.7. Brief Information about Studied Region
2.8. Ethical Control
3. Results
3.1. Identified Variants in the GJB2 Gene
3.2. GJB2 Genotypes in Patients with HI
3.3. Contribution of the GJB2 Variants to the Etiology of HI in Buryat and Russian Patients
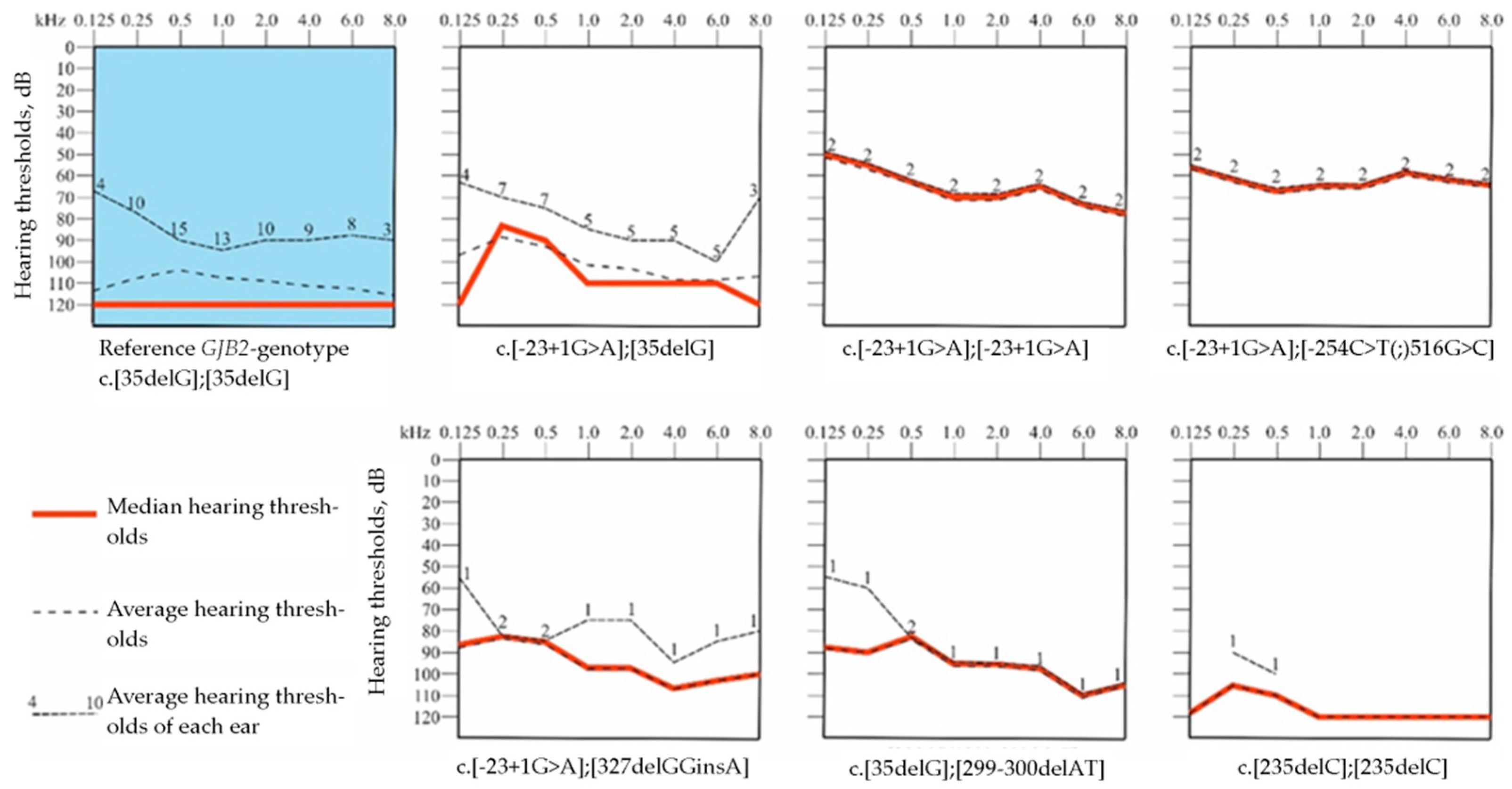
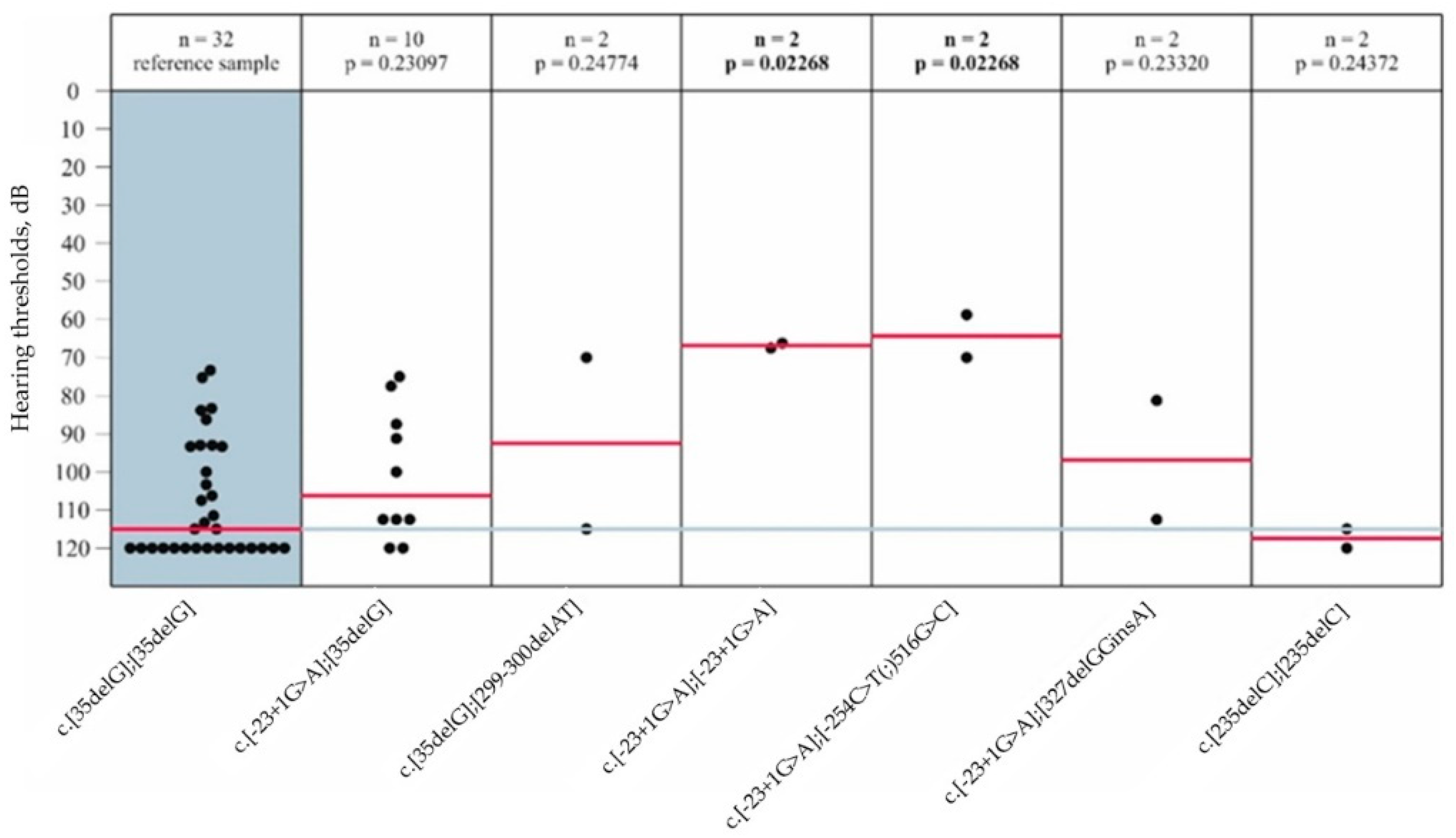
3.4. Audiological Analysis in Patients with GJB2 Gene Variants
3.5. Genotypes of 12 SNP Markers Flanking the GJB2 Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chan, D.K.; Chang, K.W. GJB2-associated hearing loss: Systematic review of worldwide prevalence, genotype, and auditory phenotype. Laryngoscope 2014, 124, E34–E53. [Google Scholar] [CrossRef]
- Tsukada, K.; Nishio, S.Y.; Hattori, M.; Usami, S. Ethnic-specific spectrum of GJB2 and SLC26A4 mutations: Their origin and a literature review. Ann. Otol. Rhinol. Laryngol. 2015, 124, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Adadey, S.M.; Wonkam-Tingang, E.; Twumasi Aboagye, E.; Nayo-Gyan, D.W.; Boatemaa Ansong, M.; Quaye, O.; Awandare, G.A.; Wonkam, A. Connexin Genes Variants Associated with Non-Syndromic Hearing Impairment: A Systematic Review of the Global Burden. Life 2020, 10, 258. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.A.; Carmi, R.; Elbedour, K.; Yosefsberg, S.; Stone, E.M.; Sheffield, V.C. An autosomal recessive nonsyndromic-hearing-loss locus identified by DNA pooling using two inbred Bedouin kindreds. Am. J. Hum. Genet. 1996, 59, 385–391. [Google Scholar]
- Morell, R.J.; Kim, H.J.; Hood, L.J.; Goforth, L.; Friderici, K.; Fisher, R.; Van Camp, G.; Berlin, C.I.; Oddoux, C.; Ostrer, H.; et al. Mutations in the connexin 26 gene (GJB2) among Ashkenazi Jews with nonsyndromic recessive deafness. N. Engl. J. Med. 1998, 339, 1500–1505. [Google Scholar] [CrossRef]
- Van Laer, L.; Coucke, P.; Mueller, R.F.; Caethoven, G.; Flothmann, K.; Prasad, S.D.; Chamberlin, G.P.; Houseman, M.; Taylor, G.R.; Van de Heyning, C.M.; et al. A common founder for the 35delG GJB2 gene mutation in connexin 26 hearing impairment. J. Med. Genet. 2001, 38, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Park, H.J.; Ouyang, X.M.; Pandya, A.; Doi, K.; Erdenetungalag, R.; Du, L.L.; Matsushiro, N.; Nance, W.E.; Griffith, A.J.; et al. Evidence of a founder effect for the 235delC mutation of GJB2 (connexin 26) in east Asians. Hum. Genet. 2003, 114, 44–50. [Google Scholar] [CrossRef]
- RamShankar, M.; Girirajan, S.; Dagan, O.; Ravi Shankar, H.M.; Jalvi, R.; Rangasayee, R.; Avraham, K.B.; Anand, A. Contribution of connexin26 (GJB2) mutations and founder effect to non-syndromic hearing loss in India. J. Med. Genet. 2003, 40, e68. [Google Scholar] [CrossRef]
- Tekin, M.; Duman, T.; Boğoçlu, G.; İncesulu, A.; Çomak, E.; Ilhan, I.; Akar, N. Spectrum of GJB2 Mutations in Turkey Comprises Both Caucasian and Oriental Variants: Roles of Parental Consanguinity and Assortative Mating. Hum. Mutat. 2003, 21, 552–553. [Google Scholar] [CrossRef]
- Nance, W.E.; Kearsey, M.J. Relevance of connexin deafness (DFNB1) to human evolution. Am. J. Hum. Genet. 2004, 74, 1081–1087. [Google Scholar] [CrossRef]
- Common, J.E.; Di, W.L.; Davies, D.; Kelsell, D.P. Further evidence for heterozygote advantage of GJB2 deafness mutations: A link with cell survival. J. Med. Genet. 2004, 41, 573–575. [Google Scholar] [CrossRef] [PubMed]
- D’Adamo, P.; Guerci, V.I.; Fabretto, A.; Faletra, F.; Grasso, D.L.; Ronfani, L.; Montico, M.; Morgutti, M.; Guastalla, P.; Gasparini, P. Does epidermal thickening explain GJB2 high carrier frequency and heterozygote advantage? Eur. J. Hum. Genet. 2009, 17, 284–286. [Google Scholar] [CrossRef] [PubMed]
- Blanton, S.H.; Nance, W.E.; Norris, V.W.; Welch, K.O.; Burt, A.; Pandya, A.; Arnos, K.S. Fitness Among Individuals with Early Childhood Deafness: Studies in Alumni Families from Gallaudet University. Ann. Hum. Genet. 2010, 74, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Tekin, M.; Xia, X.J.; Erdenetungalag, R.; Cengiz, F.B.; White, T.W.; Radnaabazar, J.; Dangaasuren, B.; Tastan, H.; Nance, W.E.; Pandya, A. GJB2 mutations in Mongolia: Complex alleles, low frequency, and reduced fitness of the deaf. Ann. Hum. Genet. 2010, 74, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Gallant, E.; Francey, L.; Tsai, E.A.; Berman, M.; Zhao, Y.; Fetting, H.; Kaur, M.; Matthew, A.D.; Wilkens, A.; Clark, D.; et al. Homozygosity for the V37I GJB2 mutation in fifteen probands with mild to moderate sensorineural hearing impairment: Further confirmation of pathogenicity and haplotype analysis in Asian populations. Am. J. Med. Genet. A. 2013, 161, 2148–2157. [Google Scholar] [CrossRef]
- Vuckovic, D.; Dallapiccola, B.; Franzè, A.; Mauri, L.; Perrone, M.D.; Gasparini, P. Connexin 26 variant carriers have a better gastrointestinal health: Is this the heterozygote advantage? Eur. J. Hum. Genet. 2015, 23, 563–564. [Google Scholar] [CrossRef]
- Carranza, C.; Menendez, I.; Herrera, M.; Castellanos, P.; Amado, C.; Maldonado, F.; Rosales, L.; Escobar, N.; Guerra, M.; Alvarez, D.; et al. A Mayan founder mutation is a common cause of deafness in Guatemala. Clin. Genet. 2016, 89, 461–465. [Google Scholar] [CrossRef]
- Solovyev, A.V.; Barashkov, N.A.; Teryutin, F.M.; Pshennikova, V.G.; Romanov, G.P.; Rafailov, A.M.; Sazonov, N.N.; Dzhemileva, L.U.; Tomsky, M.I.; Posukh, O.L.; et al. Selective Heterozygous Advantage of Carriers of c.-23+1G>A Mutation in GJB2 Gene Causing Autosomal Recessive Deafness 1A. Bull. Exp. Biol. Med. 2019, 167, 380–383. [Google Scholar] [CrossRef]
- Braun, D.C.; Jain, S.; Epstein, E.; Greenwald, B.H.; Herold, B.; Gray, M. Deaf intermarriage has limited effect on the prevalence of recessive deafness and no effect on underlying allelic frequency. PLoS ONE 2020, 15, e90972. [Google Scholar] [CrossRef]
- Shinagawa, J.; Moteki, H.; Nishio, S.; Noguchi, Y.; Usami, S. Haplotype Analysis of GJB2 Mutations: Founder Effect or Mutational Hot Spot? Genes 2020, 11, 250. [Google Scholar] [CrossRef]
- Zytsar, M.V.; Bady-Khoo, M.S.; Danilchenko, V.Y.; Maslova, E.A.; Barashkov, N.A.; Morozov, I.V.; Bondar, A.A.; Posukh, O.L. High Rates of Three Common GJB2 Mutations c.516G>C, c.-23+1G>A, c.235delC in Deaf Patients from Southern Siberia Are Due to the Founder Effect. Genes 2020, 11, 833. [Google Scholar] [CrossRef]
- Garcia-Vega, L.; O’Shaughnessy, E.M.; Albuloushi, A.; Martin, P.E. Connexins and the Epithelial Tissue Barrier: A Focus on Connexin 26. Biology 2021, 10, 59. [Google Scholar] [CrossRef]
- Jiang, Y.; Huang, S.; Zhang, Y.; Fang, N.; Liu, Q.; Liu, Y.; Bai, L.; Han, D.; Dai, P. Evolutionary origin of pathogenic GJB2 alleles in China. Clin. Genet. 2022, 102, 305–313. [Google Scholar] [CrossRef]
- Aboagye, E.T.; Adadey, S.M.; Esoh, K.; Jonas, M.; de Kock, C.; Amenga-Etego, L.; Awandare, G.A.; Wonkam, A. Age Estimate of GJB2-p.(Arg143Trp) Founder Variant in Hearing Impairment in Ghana, Suggests Multiple Independent Origins across Populations. Biology 2022, 11, 476. [Google Scholar] [CrossRef] [PubMed]
- Dia, Y.; Adadey, S.M.; Diop, J.P.D.; Aboagye, E.T.; Ba, S.A.; De Kock, C.; Ly, C.A.T.; Oluwale, O.G.; Sène, A.R.G.; Sarr, P.D.; et al. GJB2 Is a Major Cause of Non-Syndromic Hearing Impairment in Senegal. Biology 2022, 11, 795. [Google Scholar] [CrossRef]
- Aboagye, E.T.; Adadey, S.M.; Wonkam-Tingang, E.; Amenga-Etego, L.; Awandare, G.A.; Wonkam, A. Global Distribution of Founder Variants Associated with Non-Syndromic Hearing Impairment. Genes 2023, 14, 399. [Google Scholar] [CrossRef] [PubMed]
- Solovyev, A.V.; Kushniarevich, A.; Bliznetz, E.; Bady-Khoo, M.; Lalayants, M.R.; Markova, T.G.; Minárik, G.; Kádasi, L.; Metspalu, E.; Pshennikova, V.G.; et al. A common founder effect of the splice site variant c.-23+1G >A in GJB2 gene causing autosomal recessive deafness 1A (DFNB1A) in Eurasia. Hum. Genet. 2022, 141, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Romanov, G.P.; Smirnova, A.A.; Zamyatin, V.I.; Mukhin, A.M.; Kazantsev, F.V.; Pshennikova, V.G.; Teryutin, F.M.; Solovyev, A.V.; Fedorova, S.A.; Posukh, O.L.; et al. Agent-Based Modeling of Autosomal Recessive Deafness 1A (DFNB1A) Prevalence with Regard to Intensity of Selection Pressure in Isolated Human Population. Biology 2022, 11, 257. [Google Scholar] [CrossRef]
- Bliznetz, E.A.; Lalayants, M.R.; Markova, T.G.; Balanovsky, O.P.; Balanovska, E.V.; Skhalyakho, R.A.; Pocheshkhova, E.A.; Nikitina, N.V.; Voronin, S.V.; Kudryashova, E.K.; et al. Update of the GJB2/DFNB1 mutation spectrum in Russia: A founder Ingush mutation del(GJB2-D13S175) is the most frequent among other large deletions. J. Hum. Genet. 2017, 62, 789–795. [Google Scholar] [CrossRef]
- Shatokhina, O.; Galeeva, N.; Stepanova, A.; Markova, T.; Lalayants, M.; Alekseeva, N.; Tavarkiladze, G.; Markova, T.; Bessonova, L.; Petukhova, M.; et al. Spectrum of Genes for Non-GJB2-Related Non-Syndromic Hearing Loss in the Russian Population Revealed by a Targeted Deafness Gene Panel. Int. J. Mol. Sci. 2022, 23, 15748. [Google Scholar] [CrossRef]
- Posukh, O.; Pallares-Ruiz, N.; Tadinova, V.; Osipova, L.; Claustres, M.; Roux, A.F. First molecular screening of deafness in the Altai Republic population. BMC Med. Genet. 2005, 6, 12. [Google Scholar] [CrossRef] [PubMed]
- Barashkov, N.A.; Pshennikova, V.G.; Posukh, O.L.; Teryutin, F.M.; Solovyev, A.V.; Klarov, L.A.; Romanov, G.P.; Gotovtsev, N.N.; Kozhevnikov, A.A.; Kirillina, E.V.; et al. Spectrum and Frequency of the GJB2 Gene Pathogenic Variants in a Large Cohort of Patients with Hearing Impairment Living in a Subarctic Region of Russia (the Sakha Republic). PLoS ONE 2016, 11, e0156300. [Google Scholar] [CrossRef] [PubMed]
- Posukh, O.L.; Zytsar, M.V.; Bady-Khoo, M.S.; Danilchenko, V.Y.; Maslova, E.A.; Barashkov, N.A.; Bondar, A.A.; Morozov, I.V.; Maximov, V.N.; Voevoda, M.I. Unique Mutational Spectrum of the GJB2 Gene and its Pathogenic Contribution to Deafness in Tuvinians (Southern Siberia, Russia): A High Prevalence of Rare Variant c.516G>C (p.Trp172Cys). Genes 2019, 10, 429. [Google Scholar] [CrossRef] [PubMed]
- Sirmaci, A.; Akcayoz-Duman, D.; Tekin, M. The c.IVS1+1G>A mutation in the GJB2 gene is prevalent and large deletions involving the GJB6 gene are not present in the Turkish population. J. Genet. 2006, 85, 213–216. [Google Scholar] [CrossRef]
- Kelley, P.M.; Harris, D.J.; Comer, B.C.; Askew, J.W.; Fowler, T.; Smith, S.D.; Kimberling, W.J. Novel mutations in the connexin 26 gene (GJB2) that cause autosomal recessive (DFNB1) hearing loss. Am. J. Hum. Genet. 1998, 62, 792–799. [Google Scholar] [CrossRef]
- Kelsell, D.P.; Dunlop, J.; Stevens, H.P.; Lench, N.J.; Liang, J.N.; Parry, G.; Mueller, R.F.; Leigh, I.M. Connexin 26 mutations in hereditary non-syndromic sensorineural deafness. Nature 1997, 387, 80–83. [Google Scholar] [CrossRef]
- Zelante, L.; Gasparini, P.; Estivill, X.; Melchionda, S.; D’Agruma, L.; Govea, N.; Milá, M.; Monica, M.D.; Lutfi, J.; Shohat, M.P.; et al. Connexin26 mutations associated with the most common form of non-syndromic neurosensory autosomal recessive deafness (DFNB1) in Mediterraneans. Hum. Mol. Genet. 1997, 6, 1605–1609. [Google Scholar] [CrossRef]
- del Castillo, F.J.; Rodríguez-Ballesteros, M.; Alvarez, A.; Hutchin, T.; Leonardi, E.; de Oliveira, C.A.; Azaiez, H.; Brownstein, Z.; Avenarius, M.R.; Marlin, S.; et al. A novel deletion involving the connexin-30 gene, del(GJB6-d13s1854), found in trans with mutations in the GJB2 gene (connexin-26) in subjects with DFNB1 non-syndromic hearing impairment. J. Med. Genet. 2005, 42, 588–594. [Google Scholar] [CrossRef]
- Solovyev, A.V.; Barashkov, N.A.; Bady-Khoo, M.S.; Zytsar, M.V.; Posukh, O.L.; Romanov, G.P.; Rafailov, A.M.; Sazonov, N.N.; Alexeev, A.N.; Dzhemileva, L.U.; et al. Reconstruction of SNP haplotypes with mutation c.-23+1G>A in human gene GJB2 (Chromosome 13) in some populations of Eurasia. Russ. J. Genet. 2017, 53, 936. [Google Scholar] [CrossRef]
- Tang, H.Y.; Fang, P.; Ward, P.A.; Schmitt, E.; Darilek, S.; Manolidis, S.; Oghalai, J.S.; Roa, B.B.; Alford, R.L. DNA sequence analysis of GJB2, encoding connexin 26: Observations from a population of hearing impaired cases and variable carrier rates, complex genotypes, and ethnic stratification of alleles among controls. Am. J. Med. Genet. A 2006, 140, 2401–2415. [Google Scholar] [CrossRef]
- Choi, S.Y.; Lee, K.Y.; Kim, H.J.; Kim, H.K.; Chang, Q.; Park, H.J.; Jeon, C.J.; Lin, X.; Bok, J.; Kim, U.K. Functional evaluation of GJB2 variants in nonsyndromic hearing loss. Mol. Med. 2011, 17, 550–556. [Google Scholar] [CrossRef]
- Barashkov, N.A.; Dzhemileva, L.U.; Fedorova, S.A.; Teryutin, F.M.; Posukh, O.L.; Fedotova, E.E.; Lobov, S.L.; Khusnutdinova, E.K. Autosomal recessive deafness 1A (DFNB1A) in Yakut population isolate in Eastern Siberia: Extensive accumulation of the splice site mutation IVS1+1G>A in GJB2 gene as a result of founder effect. J. Hum. Genet. 2011, 56, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Erdenechuluun, J.; Lin, Y.H.; Ganbat, K.; Bataakhuu, D.; Makhbal, Z.; Tsai, C.Y.; Lin, Y.H.; Chan, Y.H.; Hsu, C.J.; Hsu, W.C. Unique spectra of deafness-associated mutations in Mongolians provide insights into the genetic relationships among Eurasian populations. PLoS ONE 2018, 13, e0209797. [Google Scholar] [CrossRef] [PubMed]
- Cryns, K.; Orzan, E.; Murgia, A.; Huygen, P.L.; Moreno, F.; del Castillo, I.; Chamberlin, G.P.; Azaiez, H.; Prasad, S.; Cucci, R.A.; et al. Genotype-phenotype correlation for GJB2 (connexin 26) deafness. J. Med. Genet. 2004, 41, 147–514. [Google Scholar] [CrossRef] [PubMed]
- Snoeckx, R.L.; Huygen, P.L.; Feldmann, D.; Marlin, S.; Denoyelle, F.; Waligora, J.; Mueller-Malesinska, M.; Pollak, A.; Ploski, R.; Murgia, A.; et al. GJB2 mutations and degree of hearing loss: A multicenter study. Am. J. Hum. Genet. 2005, 77, 945–957. [Google Scholar] [CrossRef] [PubMed]
- Abidi, O.; Boulouiz, R.; Nahili, H.; Imken, L.; Rouba, H.; Chafik, A.; Barakat, A. The Analysis of Three Markers Flanking GJB2 Gene Suggests a Single Origin of the Most Common 35delG Mutation in the Moroccan Population. Biochem. Biophys. Res. Commun. 2008, 377, 971–974. [Google Scholar] [CrossRef]
- Norouzi, V.; Azizi, H.; Fattahi, Z.; Esteghamat, F.; Bazazzadegan, N.; Nishimura, C.; Nikzat, N.; Jalalvand, K.; Kahrizi, K.; Smith, R.J. Did the GJB2 35delG Mutation Originate in Iran? Am. J. Med. Genet. Part A 2011, 155, 2453–2458. [Google Scholar] [CrossRef]
- Tekin, M.; Boğoclu, G.; Arican, S.T.; Orman, M.N.; Tastan, H.; Elsayed, S.; Akar, N. Evidence for Single Origins of 35delG and DelE120 Mutations in the GJB2 Gene in Anatolia. Clin. Genet. 2005, 67, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Kokotas, H.; Van Laer, L.; Grigoriadou, M.; Iliadou, V.; Economides, J.; Pomoni, S.; Pampanos, A.; Eleftheriades, N.; Ferekidou, E.; Korres, S. Strong Linkage Disequilibrium for the Frequent GJB2 35delG Mutation in the Greek Population. Am. J. Med. Genet. Part A 2008, 146, 2879–2884. [Google Scholar] [CrossRef]
- Kokotas, H.; Grigoriadou, M.; Villamar, M.; Giannoulia-Karantana, A.; Del Castillo, I.; Petersen, M.B. Hypothesizing an ancient greek origin of the GJB2 35delG Mutation: Can science meet history? Genet. Test. Mol. Biomark. 2010, 14, 183–187. [Google Scholar] [CrossRef]
- Zytsar, M.V.; Barashkov, N.A.; Bady-Khoo, M.S.; Shubina-Olejnik, O.A.; Danilenko, N.G.; Bondar, A.A.; Morozov, I.V.; Solovyev, A.V.; Danilchenko, V.Y.; Maximov, V.N.; et al. Updated Carrier Rates for c.35delG (GJB2) Associated with Hearing Loss in Russia and Common c.35delG Haplotypes in Siberia. BMC Med. Genet. 2018, 19, 138. [Google Scholar] [CrossRef] [PubMed]
- Tekin, M.; Akar, N.; Cin, Ş.; Blanton, S.; Xia, X.; Liu, X.; Nance, W.; Pandya, A. Connexin 26 (GJB2) mutations in the Turkish population: Implications for the origin and high frequency of the 35delG mutation in Caucasians. Qual. Life Res. 2001, 108, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Rothrock, C.R.; Murgia, A.; Sartorato, E.L.; Leonardi, E.; Wei, S.; Lebeis, S.L.; Yu, L.E.; Elfenbein, J.L.; Fisher, R.A.; Friderici, K.H. Connexin 26 35delG does not represent a mutational hotspot. Qual. Life Res. 2003, 113, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Balci, B.; Gerçeker, F.O.; Aksoy, S.; Sennaroğlu, G.; Kalay, E.; Sennaroğlu, L.; Dinçer, P. Identification of an ancestral haplotype of the 35delG mutation in the GJB2 (connexin 26) gene responsible for autosomal recessive non-syndromic hearing loss in families from the Eastern Black Sea Region in Turkey. Turk. J. Pediatr. 2005, 47, 213–221. [Google Scholar] [PubMed]
- Shahin, H.; Walsh, T.; Sobe, T.; Lynch, E.; King, M.-C.; Avraham, K.B.; Kanaan, M. Genetics of congenital deafness in the Palestinian population: Multiple connexin 26 alleles with shared origins in the Middle East. Qual. Life Res. 2002, 110, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Belguith, H.; Hajji, S.; Salem, N.; Charfeddine, I.; Lahmar, I.; Amor, M.B.; Ouldim, K.; Chouery, E.; Driss, N.; Drira, M.; et al. Analysis of GJB2 mutation: Evidence for a Mediterranean ancestor for the 35delG mutation. Clin. Genet. 2005, 68, 188–189. [Google Scholar] [PubMed]
- Dzhemileva, L.U.; Posukh, O.L.; Barashkov, N.A.; Fedorova, S.A.; Teryutin, F.M.; Akhmetova, V.L.; Khidiyatova, I.M.; Khusainova, R.I.; Lobov, S.L.; Khusnutdinova, E.K. Haplotype diversity and reconstruction of ancestral haplotype associated with the c.35delG Mutation in the GJB2 (Cx26) gene among the Volgo-Ural populations of Russia. Acta Naturae 2011, 3, 52–63. [Google Scholar] [CrossRef]
- Kudo, T.; Ikeda, K.; Kure, S.; Matsubara, Y.; Oshima, T.; Watanabe, K.-I.; Kawase, T.; Narisawa, K.; Takasaka, T. Novel mutations in the connexin 26 gene (GJB2) responsible for childhood deafness in the Japanese population. Am. J. Med. Genet. 2000, 90, 141–145. [Google Scholar] [CrossRef]
- Liu, X.; Xia, X.; Ke, X.; Ouyang, X.; Du, L.; Liu, Y.; Angeli, S.; Telischi, F.F.; Nance, W.E.; Balkany, T.; et al. The prevalence of connexin 26 (GJB2) mutations in the Chinese population. Qual. Life Res. 2002, 111, 394–397. [Google Scholar] [CrossRef]
- Ohtsuka, A.; Yuge, I.; Kimura, S.; Namba, A.; Abe, S.; Van Laer, L.; Van Camp, G.; Usami, S. GJB2 deafness gene shows a specific spectrum of mutations in Japan, including a frequent founder mutation. Qual. Life Res. 2003, 112, 329–333. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | n (%) | Males | Females | Age |
|---|---|---|---|---|
| Buryats | 79 (47.8%) | 31 (39.2%) | 48 (60.8%) | 46.9 ± 14.3 |
| Russians | 76 (46.1%) | 32 (42.1%) | 44 (57.9%) | 54.8 ± 15.7 |
| Others | 10 (6.1%) | 5 (50.0%) | 5 (50.0%) | 49.2 ± 15.6 |
| Total | 165 (100%) | 68 (41.2%) | 97 (58.8%) | 50.7 ± 15.5 |
| # | GJB2-Genotypes | Buryats | Russians | Others | Total | ||||
|---|---|---|---|---|---|---|---|---|---|
| n = 79 | F (%) | n = 76 | F (%) | n = 10 | F (%) | n = 165 | F (%) | ||
| GJB2 genotypes with biallelic PLP variants | |||||||||
| 1 | c.[-23+1G>A];[-23+1G>A] | 1 | 1.2 | - | - | - | - | 1 | 0.6 |
| 2 | c.[-23+1G>A];[35delG] | - | - | 5 | 6.6 | - | - | 5 | 3.0 |
| 3 | c.[-23+1G>A];[327_328delGGinsA] | 1 | 1.2 | - | - | - | - | 1 | 0.6 |
| 4 | c.[-23+1G>A];[-254C>T(;)516G>C] | 1 | 1.2 | - | - | - | - | 1 | 0.6 |
| 5 | c.[35delG];[35delG] | - | - | 16 | 21.1 | - | - | 16 | 9.7 |
| 6 | c.[35delG];[299_300delAT] | - | - | 1 | 1.3 | - | - | 1 | 0.6 |
| 7 | c.[235delC];[235delC] | 1 | 1.2 | - | - | - | - | 1 | 0.6 |
| Total | 4 | 5.1 | 22 | 28.9 | - | - | 26 | 15.8 | |
| GJB2 genotypes with monoallelic PLP variants | |||||||||
| 8 | c.[-23+1G>A];[wt] | 1 | 1.2 | 1 | 1.3 | - | - | 2 | 1.2 |
| 9 | c.[23C>T(;)457G>A];[457G>A] | - | - | 1 | 1.3 | - | - | 1 | 0.6 |
| 10 | c.[35delG];[wt] | - | - | 1 | 1.3 | - | - | 1 | 0.6 |
| 11 | c.[101T>C];[wt] | - | - | 1 | 1.3 | - | - | 1 | 0.6 |
| 12 | c.[109G>A];[wt] | - | - | 2 | 2.6 | - | - | 2 | 1.2 |
| 13 | c.[109G>A];[79G>A] | 1 | 1.2 | 1 | 1.3 | - | - | 2 | 1.2 |
| Total | 2 | 2.5 | 7 | 9.2 | - | - | 9 | 5.5 | |
| GJB2 genotypes with benign variants | |||||||||
| 14 | c.[-49G>A];[wt] | - | - | 1 | 1.3 | - | - | 1 | 0.6 |
| 15 | c.[79G>A];[wt] | 18 | 22.8 | 1 | 1.3 | 1 | 10 | 20 | 12.1 |
| 16 | c.[79G>A(;)341A>G];[wt] | 5 | 6.3 | 2 | 2.6 | 2 | 20 | 9 | 5.5 |
| 17 | c.[79G>A(;)341A>G];[79G>A] | 1 | 1.2 | - | - | - | - | 1 | 0.6 |
| 18 | c.[457G>A];[wt] | - | - | 1 | 1.3 | - | - | 1 | 0.6 |
| Total | 24 | 30.4 | 5 | 6.6 | 3 | 30 | 32 | 19.4 | |
| GJB2 genotype without sequence changes (wild type) | |||||||||
| c.[wt];[wt] | 49 | 62.0 | 42 | 55.3 | 7 | 70 | 98 | 59.4 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pshennikova, V.G.; Teryutin, F.M.; Cherdonova, A.M.; Borisova, T.V.; Solovyev, A.V.; Romanov, G.P.; Morozov, I.V.; Bondar, A.A.; Posukh, O.L.; Fedorova, S.A.; et al. The GJB2 (Cx26) Gene Variants in Patients with Hearing Impairment in the Baikal Lake Region (Russia). Genes 2023, 14, 1001. https://doi.org/10.3390/genes14051001
Pshennikova VG, Teryutin FM, Cherdonova AM, Borisova TV, Solovyev AV, Romanov GP, Morozov IV, Bondar AA, Posukh OL, Fedorova SA, et al. The GJB2 (Cx26) Gene Variants in Patients with Hearing Impairment in the Baikal Lake Region (Russia). Genes. 2023; 14(5):1001. https://doi.org/10.3390/genes14051001
Chicago/Turabian StylePshennikova, Vera G., Fedor M. Teryutin, Alexandra M. Cherdonova, Tuyara V. Borisova, Aisen V. Solovyev, Georgii P. Romanov, Igor V. Morozov, Alexander A. Bondar, Olga L. Posukh, Sardana A. Fedorova, and et al. 2023. "The GJB2 (Cx26) Gene Variants in Patients with Hearing Impairment in the Baikal Lake Region (Russia)" Genes 14, no. 5: 1001. https://doi.org/10.3390/genes14051001