
Leukocyte Tyrosine Kinase (Ltk) Is the Mendelian Determinant of the Axolotl Melanoid Color Variant

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
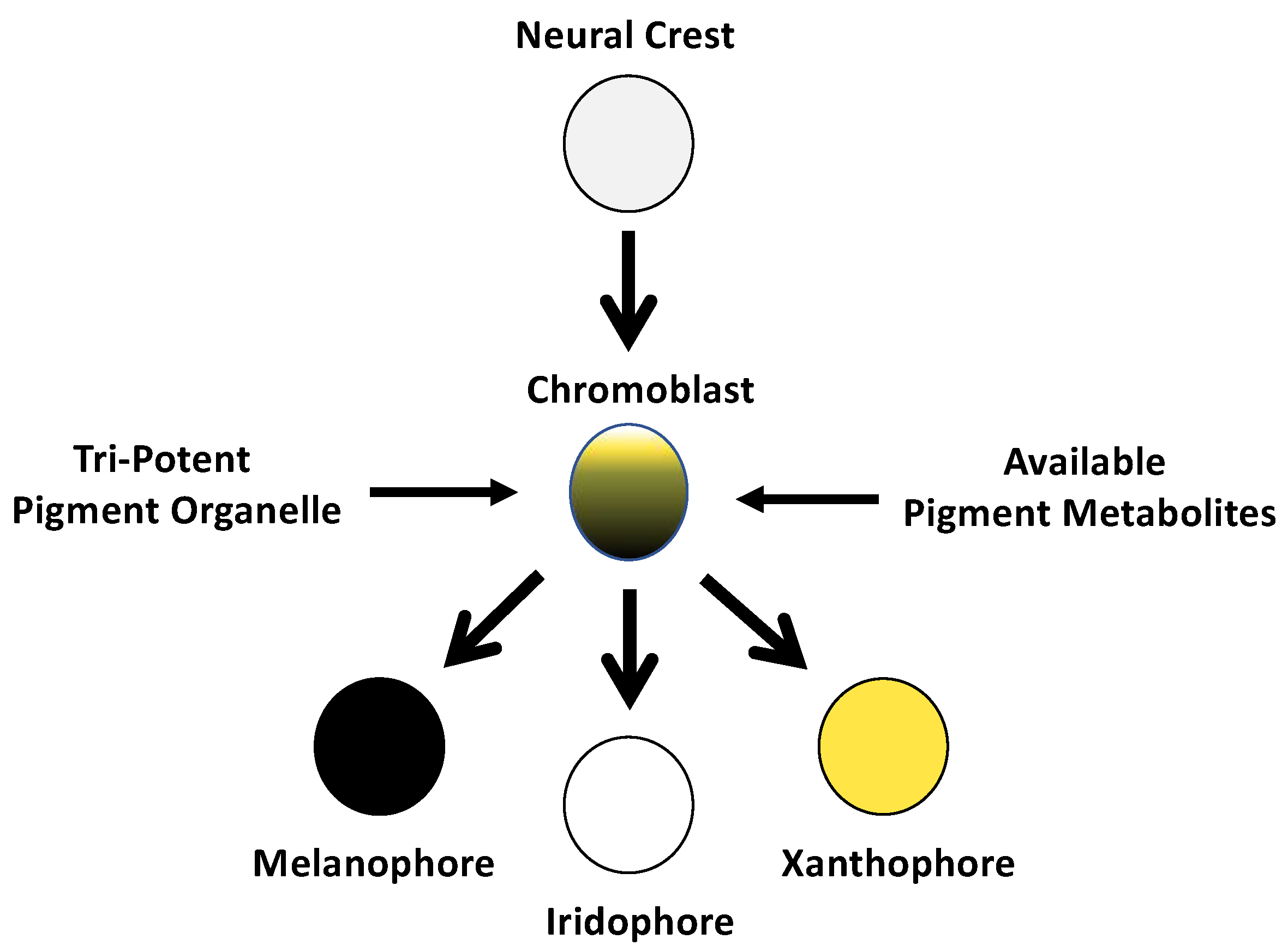
:1. Introduction
2. Materials and Methods
2.1. Animal Procedures
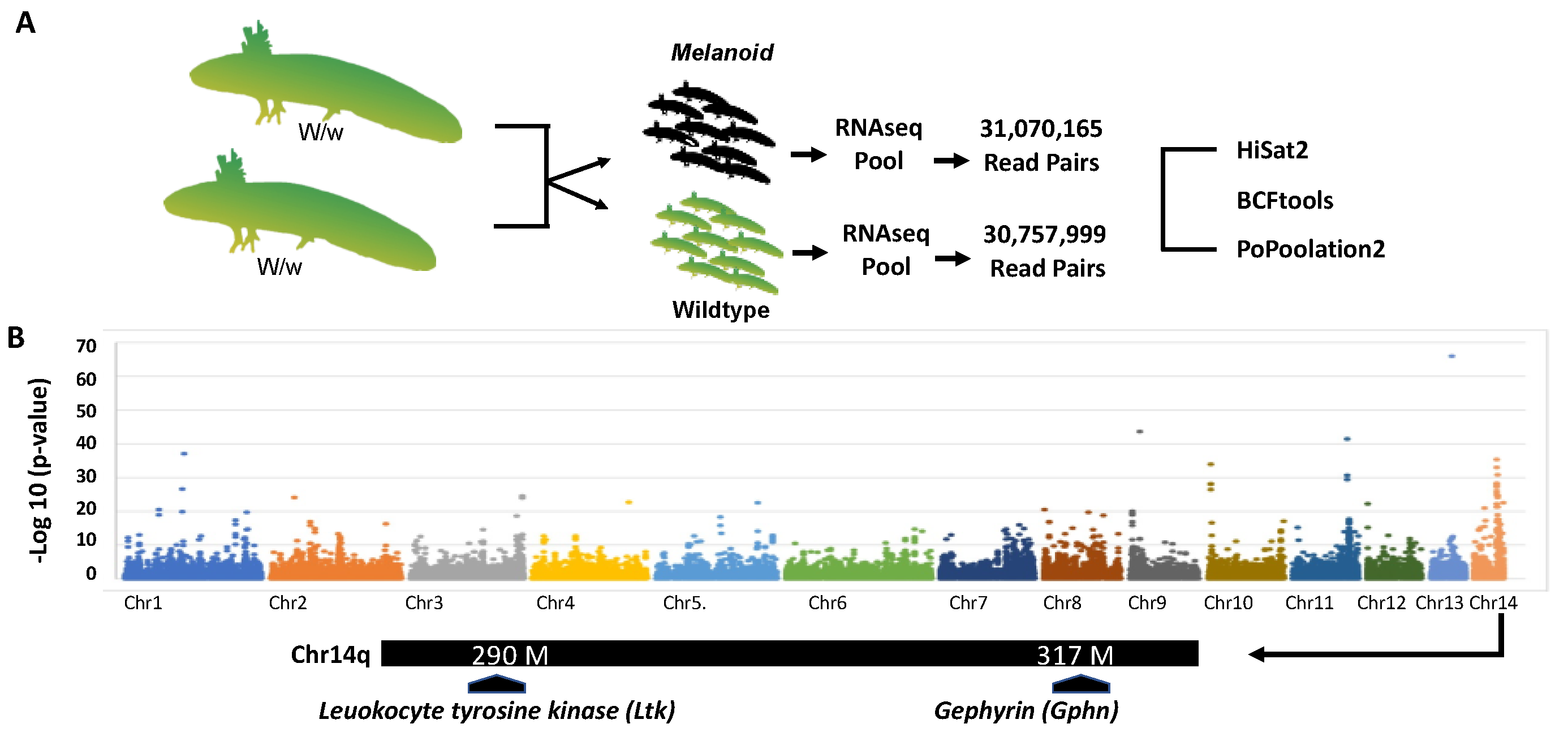
2.2. Bulked Segregant RNA-Seq
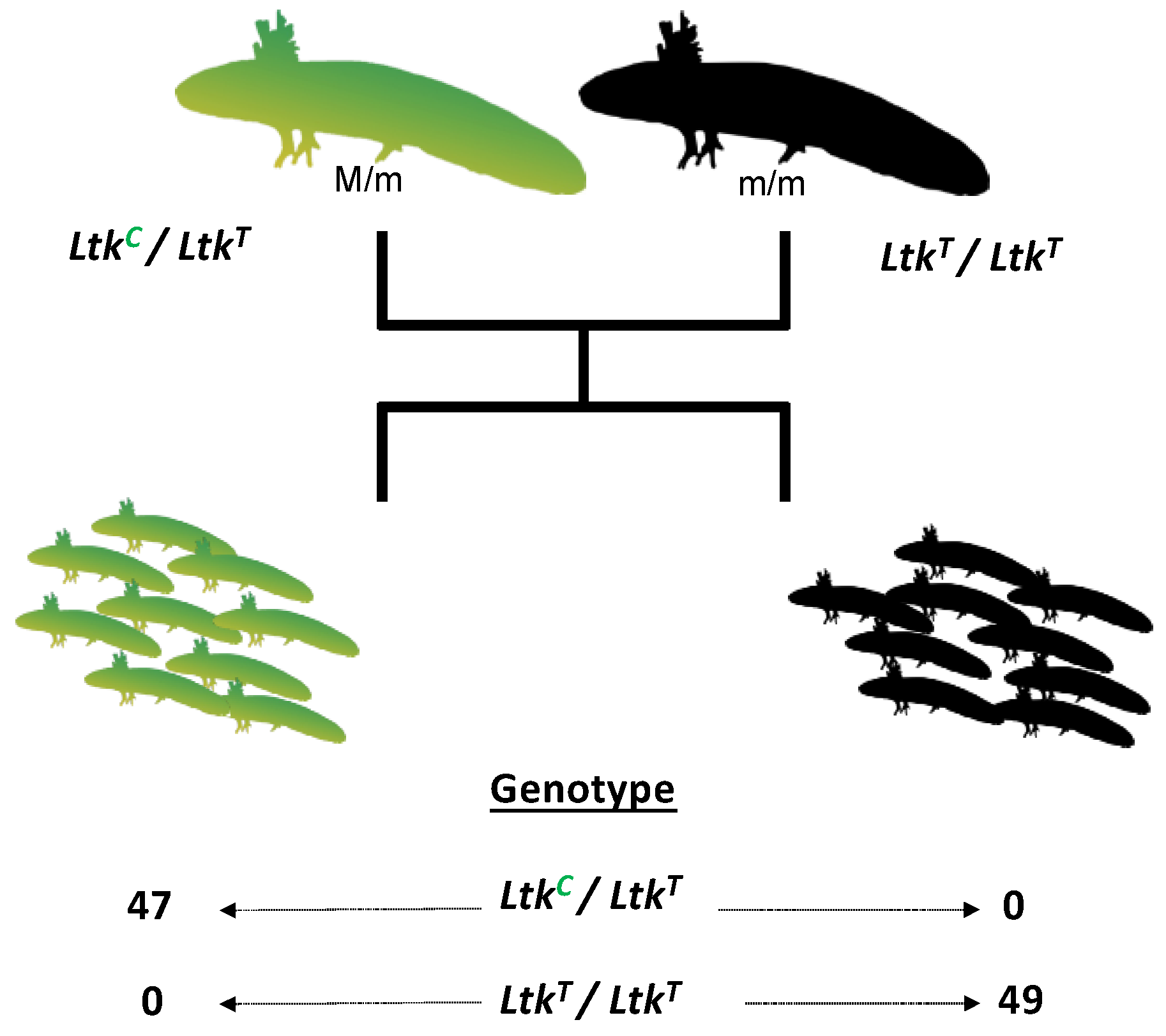
2.3. Genetic Linkage Analysis
2.4. CRISPR-CAS9 Disruption of Ltk and Gphn
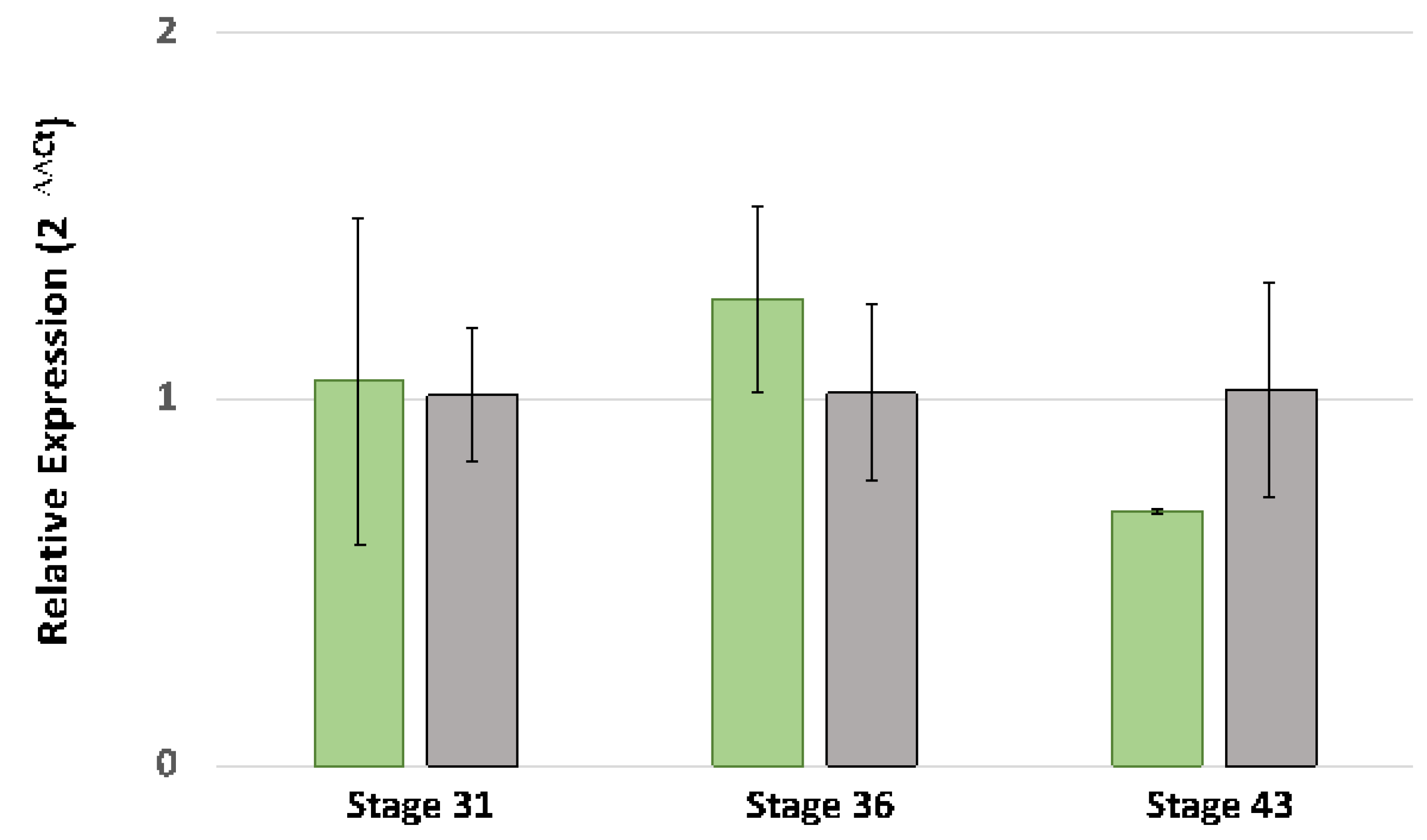
2.5. Ltk Quantitative PCR
2.6. Ltk Genomic Sequencing
3. Results
3.1. Bulk Segregant RNA-Seq Identified Two Candidate Genes for Melanoid
3.2. CRISPR-Cas9 Disruption of Ltk Phenocopied Melanoid
3.3. DNA Sequencing of Ltk Exons and Quantitative PCR Analysis
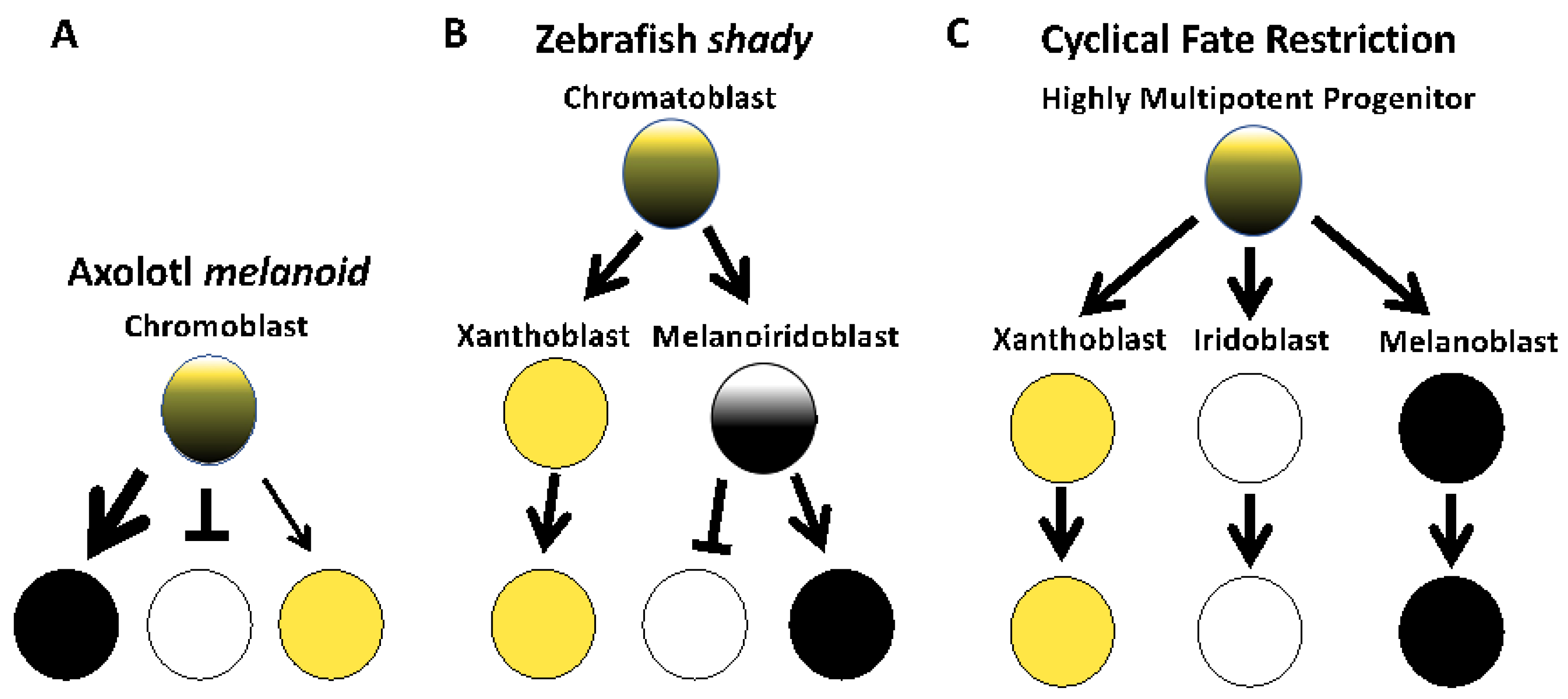
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reiß, C.; Olsson, L.; Hoßfeld, U. The history of the oldest self-sustaining laboratory animal: 150 years of axolotl research. J. Exp. Zool. Part B Mol. Dev. Evol. 2015, 324, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Häcker, V. Uber mendelschen vererbung bei axolotln. Zool. Anz. 1907, 31, 99–102. [Google Scholar]
- Woodcock, M.R.; Vaughn-Wolfe, J.; Elias, A.; Kump, D.K.; Kendall, K.D.; Timoshevskaya, N.; Timoshevskiy, V.; Perry, D.W.; Smith, J.J.; Spiewak, J.E.; et al. Identification of Mutant Genes and Introgressed Tiger Salamander DNA in the Laboratory Axolotl, Ambystoma mexicanum. Sci. Rep. 2017, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, R.R.; Bagnara, J.T. A Color Variant in the Mexican Axolotl. J. Hered. 1967, 58, 251–256. [Google Scholar] [CrossRef]
- Rivera, S.C. Embryological Analysis of Gene Action in the Melanoid Mutant of the Mexican Axolotl, Ambystoma mexicanum, Shaw. Ph.D. Thesis, Pennsylvania State University, State College, PA, USA, 1971. [Google Scholar]
- Bagnara, J.T.; Frost, S.K.; Matsumoto, J. On the Development of Pigment Patterns in Amphibians. Am. Zool. 1978, 18, 301–312. [Google Scholar] [CrossRef]
- Thorsteinsdottir, S.; Frost, S.K. Pigment cell differentiation: The relationship between pterin content, allopurinol treatment, and the melanoid gene in axolotls. Cell Diff. 1986, 19, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Bagnara, J.T.; Matsumoto, J.; Ferris, W.; Frost, S.K.; Turner, W.A., Jr.; Tchen, T.T.; Taylor, J.D. Common origin of pigment cells. Science 1979, 203, 410–415. [Google Scholar] [CrossRef]
- Lister, J.A.; Robertson, C.P.; Lepage, T.; Johnson, S.L.; Raible, D.W. nacre encodes a zebrafish microphthalmia-related protein that regulates neural-crest-derived pigment cell fate. Development 1999, 126, 3757–3767. [Google Scholar] [CrossRef]
- Lopes, S.S.; Yang, X.; Müller, J.; Carney, T.J.; McAdow, A.R.; Rauch, G.-J.; Jacoby, A.S.; Hurst, L.D.; Delfino-Machín, M.; Haffter, P.; et al. Leukocyte Tyrosine Kinase Functions in Pigment Cell Development. PLoS Genet. 2008, 4, e1000026. [Google Scholar] [CrossRef] [Green Version]
- Maderspacher, F.; Nüsslein-Volhard, C. Formation of the adult pigment pattern in zebrafish requires leopard and obelix dependent cell interactions. Development 2003, 130, 3447–3457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parichy, D.M.; Turner, J.M. Temporal and cellular requirements for Fms signaling during zebrafish adult pigment pattern development. Development 2003, 130, 817–833. [Google Scholar] [CrossRef] [Green Version]
- Frohnhöfer, H.G.; Krauss, J.; Maischein, H.-M.; Nüsslein-Volhard, C. Iridophores and their interactions with other chromatophores are required for stripe formation in zebrafish. Development 2013, 140, 2997–3007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, L.B.; Parichy, D.M. Interactions with Iridophores and the Tissue Environment Required for Patterning Melanophores and Xanthophores during Zebrafish Adult Pigment Stripe Formation. PLoS Genet. 2013, 9, e1003561. [Google Scholar] [CrossRef] [Green Version]
- Kelsh, R.N.; Sosa, K.C.; Farjami, S.; Makeev, V.; Dawes, J.H.P.; Rocco, A. Cyclical fate restriction: A new view of neural crest cell fate specification. Development 2021, 148, dev176057. [Google Scholar] [CrossRef] [PubMed]
- Subkhankulova, T.; Sosa, K.C.; Uroshlev, L.A.; Nikaido, M.; Shriever, N.; Kasianov, A.S.; Yang, X.; Rodrigues, F.S.L.M.; Carney, T.J.; Bavister, G.; et al. Zebrafish pigment cells develop directly from persistent highly multipotent progenitors. Nat. Commun. 2023, 14, 1258. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.S.; Voss, G.; Miesfeld, J.B.; Zarate-Sanchez, J.; Voss, S.R.; Glaser, T. The rax homeobox gene is mutated in the eyeless axolotl, Ambystoma mexicanum. Dev. Dyn. 2020, 250, 807–821. [Google Scholar] [CrossRef]
- Smith, J.J.; Timoshevskaya, N.; Timoshevskiy, V.A.; Keinath, M.C.; Hardy, D.; Voss, S.R. A chromosome-scale assembly of the axolotl genome. Genome Res. 2019, 29, 317–324. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Yeh, C.-T.; Tang, H.M.; Nettleton, D.; Schnable, P.S. Gene Mapping via Bulked Segregant RNA-Seq (BSR-Seq). PLoS ONE 2012, 7, e36406. [Google Scholar] [CrossRef] [Green Version]
- Schloissnig, S.; Kawaguchi, A.; Nowoshilow, S.; Falcon, F.; Otsuki, L.; Tardivo, P.; Timoshevskaya, N.; Keinath, M.C.; Smith, J.J.; Voss, S.R.; et al. The giant axolotl genome uncovers the evolution, scaling and transcriptional control of complex gene loci. Proc. Natl. Acad. Sci. USA 2021, 118, e2017176118. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H. Improving SNP discovery by base alignment quality. Bioinformatics 2011, 27, 1157–1158. [Google Scholar] [CrossRef] [PubMed]
- Kofler, R.; Pandey, R.V.; Schlötterer, C. PoPoolation2: Identifying differentiation between populations using sequencing of pooled DNA samples (Pool-Seq). Bioinformatics 2011, 27, 3435–3436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trofka, A.; Huang, B.-L.; Zhu, J.; Heinz, W.F.; Magidson, V.; Shibata, Y.; Shi, Y.-B.; Tarchini, B.; Stadler, H.S.; Kabangu, M.; et al. Genetic basis for an evolutionary shift from ancestral preaxial to postaxial limb polarity in non-urodele vertebrates. Curr. Biol. 2021, 31, 4923–4934.e5. [Google Scholar] [CrossRef] [PubMed]
- Bordzilovskaya, N.P.; Dettlaff, T.A.; Duhon, S.T.; Malacinski, G.M. Developmental stage series of axolotl embryos. In Developmental Biology of the Axolotl; Armstrong, J.B., Malacinski, G.M., Eds.; Oxford University Press: New York, NY, USA, 1989; pp. 201–219. [Google Scholar]
- Livak, K.J.; Thomas, D.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Stallmeyer, B.; Schwarz, G.; Schulze, J.; Nerlich, A.; Reiss, J.; Kirsch, J.; Mendel, R.R. The neurotransmitter receptor-anchoring protein gephyrin reconstitutes molybdenum cofactor biosynthesis in bacteria, plants, and mammalian cells. Proc. Natl. Acad. Sci. USA 1999, 96, 1333–1338. [Google Scholar] [CrossRef] [Green Version]
- Reiss, J.; Gross-Hardt, S.; Christensen, E.; Schmidt, P.; Mendel, R.R.; Schwarz, G. A Mutation in the Gene for the Neurotransmitter Receptor–Clustering Protein Gephyrin Causes a Novel Form of Molybdenum Cofactor Deficiency. Am. J. Hum. Genet. 2001, 68, 208–213. [Google Scholar] [CrossRef] [Green Version]
- Beckmann, G.; Bork, P. An adhesive domain detected in functionally diverse receptors. Trends Biochem. Sci. 1993, 18, 40–41. [Google Scholar] [CrossRef]
- Malacinski, G.M.; Brothers, J. Mutant genes in the Mexican axolotl. Science 1974, 184, 1142–1147. [Google Scholar] [CrossRef]
- Fadeev, A.; Krauss, J.; Singh, A.P.; Nüsslein-Volhard, C. Zebrafish Leucocyte tyrosine kinase controls iridophore establishment, proliferation and survival. Pigment. Cell Melanoma Res. 2016, 29, 284–296. [Google Scholar] [CrossRef]
- Mo, E.S.; Cheng, Q.; Reshetnyak, A.V.; Schlessinger, J.; Nicoli, S. Alk and Ltk ligands are essential for iridophore development in zebrafish mediated by the receptor tyrosine kinase Ltk. Proc. Natl. Acad. Sci. USA 2017, 114, 12027–12032. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kabangu, M.; Cecil, R.; Strohl, L., II; Timoshevskaya, N.; Smith, J.J.; Voss, S.R. Leukocyte Tyrosine Kinase (Ltk) Is the Mendelian Determinant of the Axolotl Melanoid Color Variant. Genes 2023, 14, 904. https://doi.org/10.3390/genes14040904
Kabangu M, Cecil R, Strohl L II, Timoshevskaya N, Smith JJ, Voss SR. Leukocyte Tyrosine Kinase (Ltk) Is the Mendelian Determinant of the Axolotl Melanoid Color Variant. Genes. 2023; 14(4):904. https://doi.org/10.3390/genes14040904
Chicago/Turabian StyleKabangu, Mirindi, Raissa Cecil, Lloyd Strohl, II, Nataliya Timoshevskaya, Jeramiah J. Smith, and Stephen R. Voss. 2023. "Leukocyte Tyrosine Kinase (Ltk) Is the Mendelian Determinant of the Axolotl Melanoid Color Variant" Genes 14, no. 4: 904. https://doi.org/10.3390/genes14040904