
A Comprehensive Pan-Cancer Analysis of the Regulation and Prognostic Effect of Coat Complex Subunit Zeta 1
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Downloading Data, Analyzing Differential Expression and Exploring the Significance of COPZ1 in Tumor
2.2. Exploring the Association of COPZ1 Expression and Special Signature
2.3. Identifying Differential Expressed Genes and Conducting Functional Enrichment Analysis
2.4. Evaluating the Association between COPZ1 Expression and Tumor Immune Microenvironment
2.5. Defining the Regulation Mechanism of COPZ1 Expression in Tumor
2.6. Cell Lines and Reagents
2.7. RNA Extraction and qRT-PCR Analysis
- COPZ1-F: 5′-GATGGAGATCGACTTTTTGCCA-3′
- COPZ1-R: 5′-TCAGTCCGATGGGTCTTGTTG-3′
- GAPDH-F: 5′-GGAGCGAGATCCCTCCAAAAT-3′
- GAPDH-R: 5′-GGCTGTTGTCATACTTCTCATGG-3′
2.8. Western Blotting
2.9. siRNA Transfection
2.10. Cell Proliferation Detection
2.11. Cell Migration and Invasion Assay
2.12. Statistical Analysis
3. Results
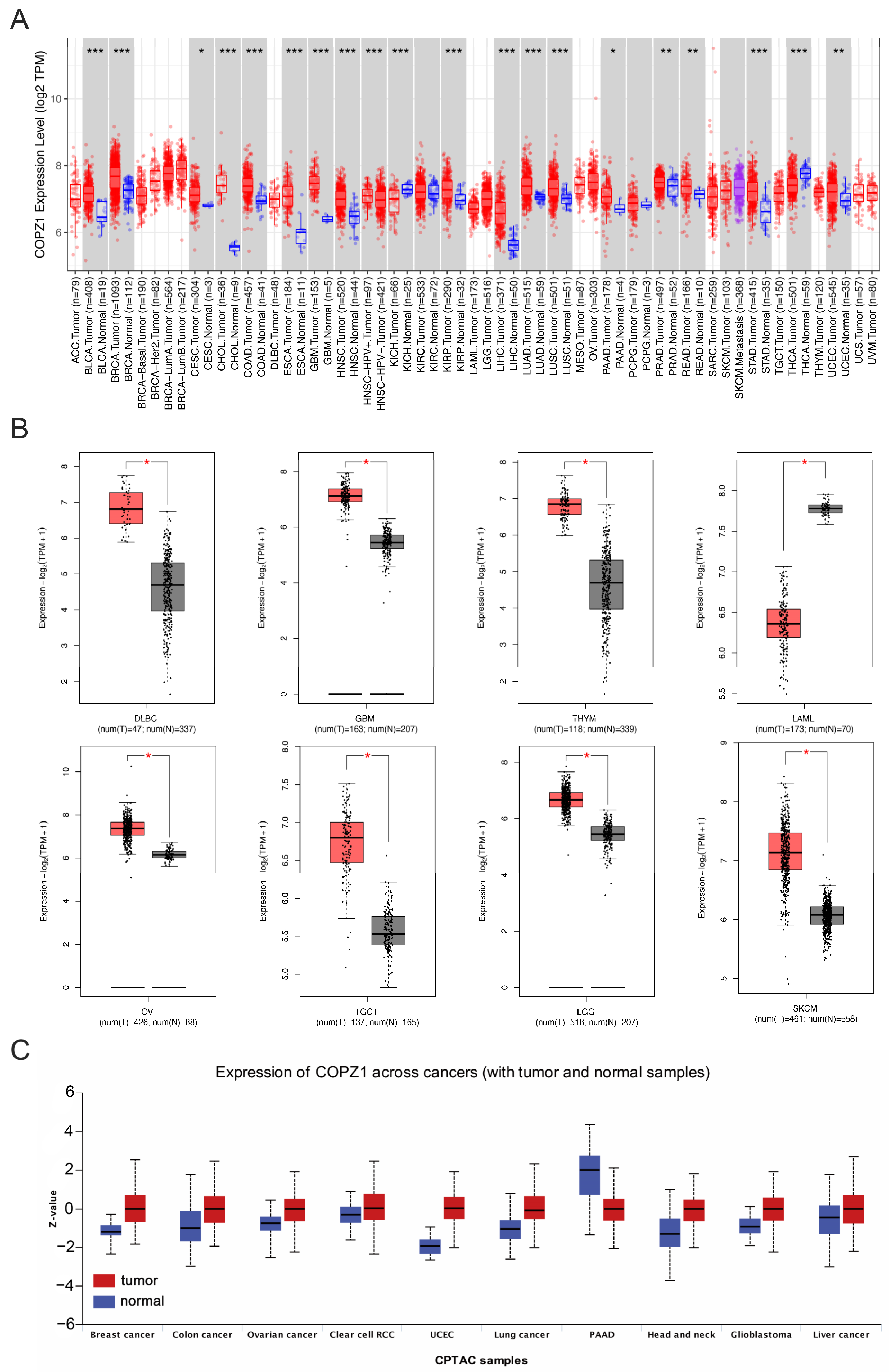
3.1. COPZ1 Expression Profile in Pan-Cancers and Corresponding Normal Tissues
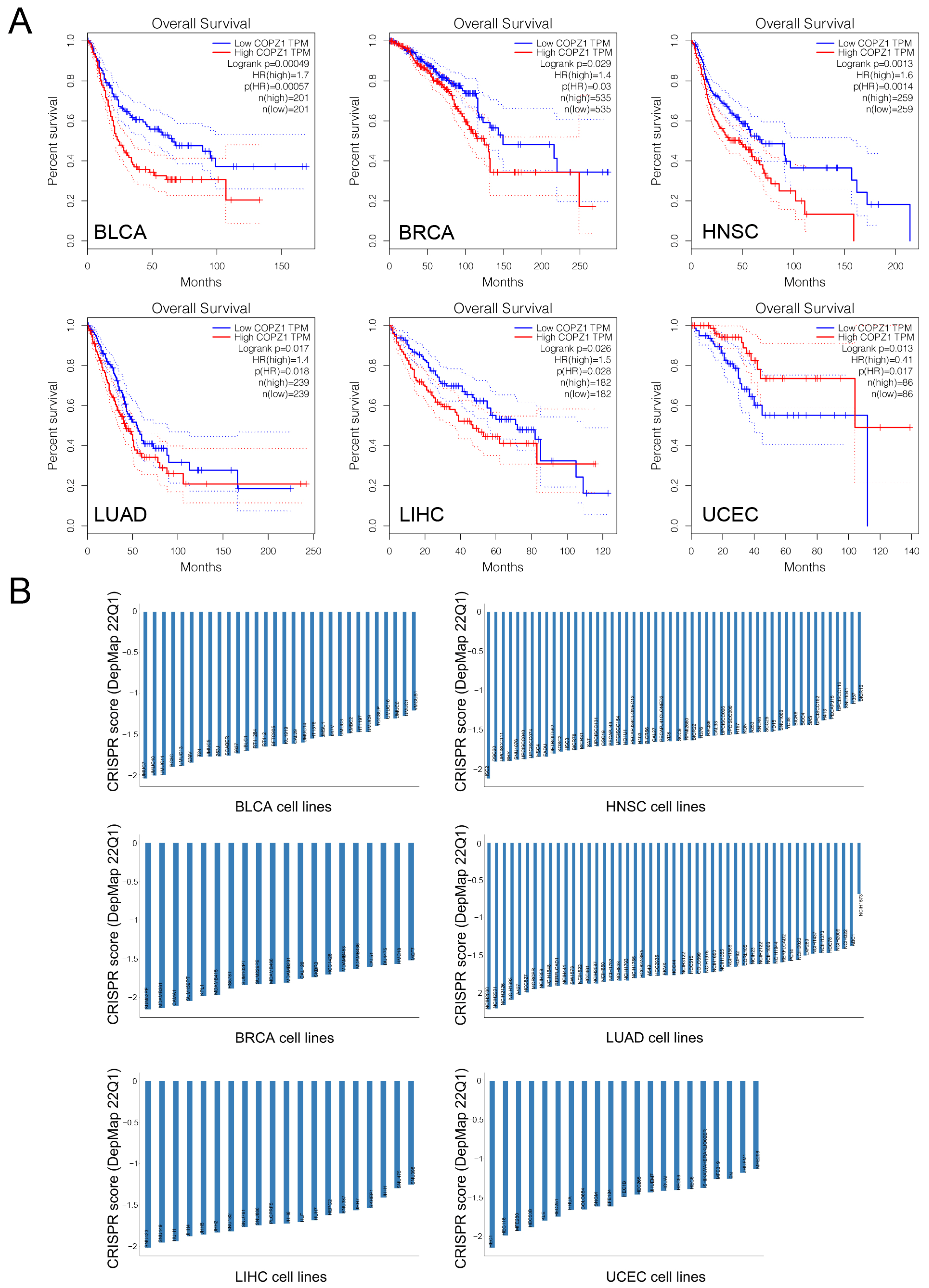
3.2. High COPZ1 Expression Predicts Poor Prognosis in Multiple Cancers
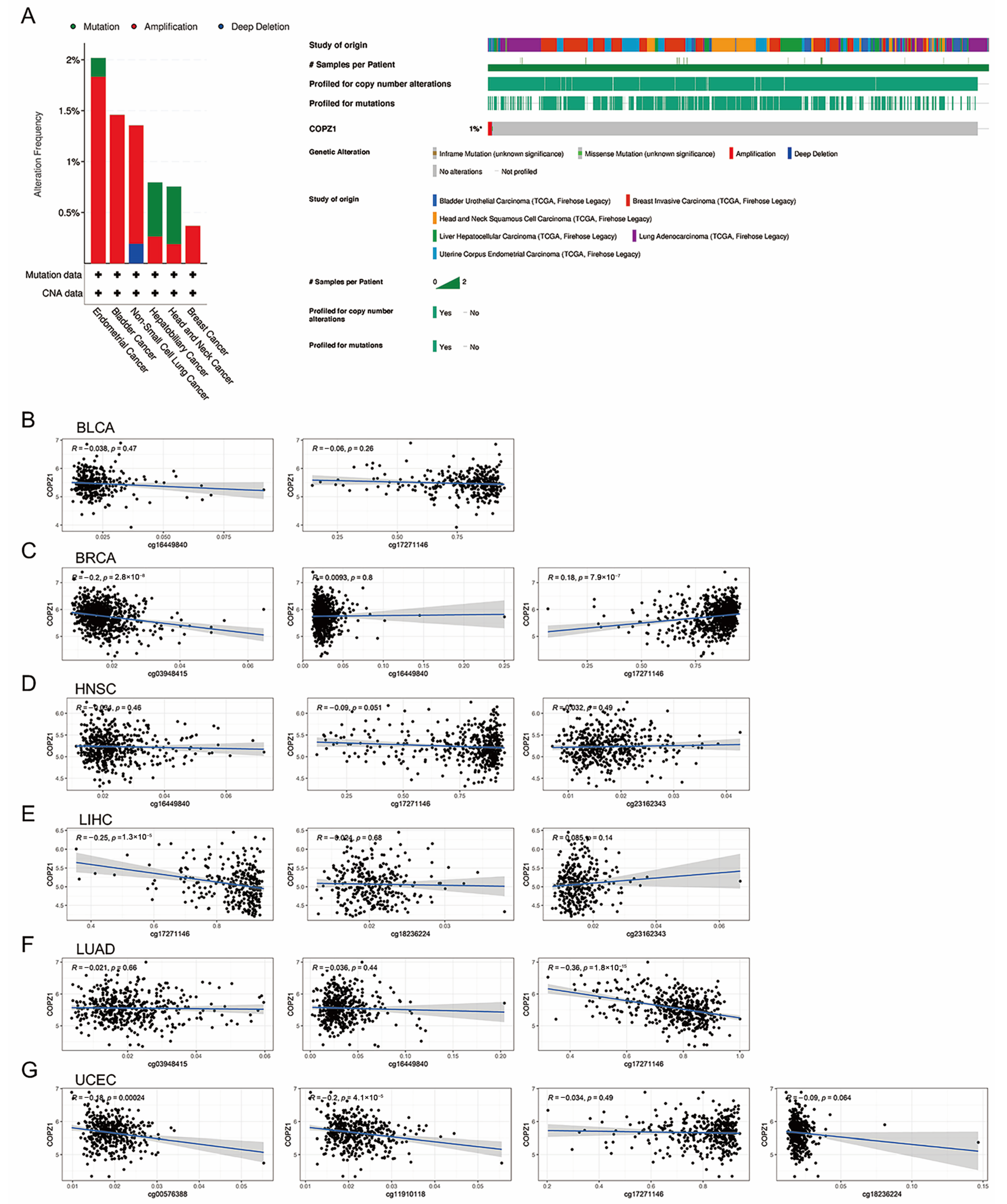
3.3. Investigation of the Regulation Mechanism of the COPZ1 Expression
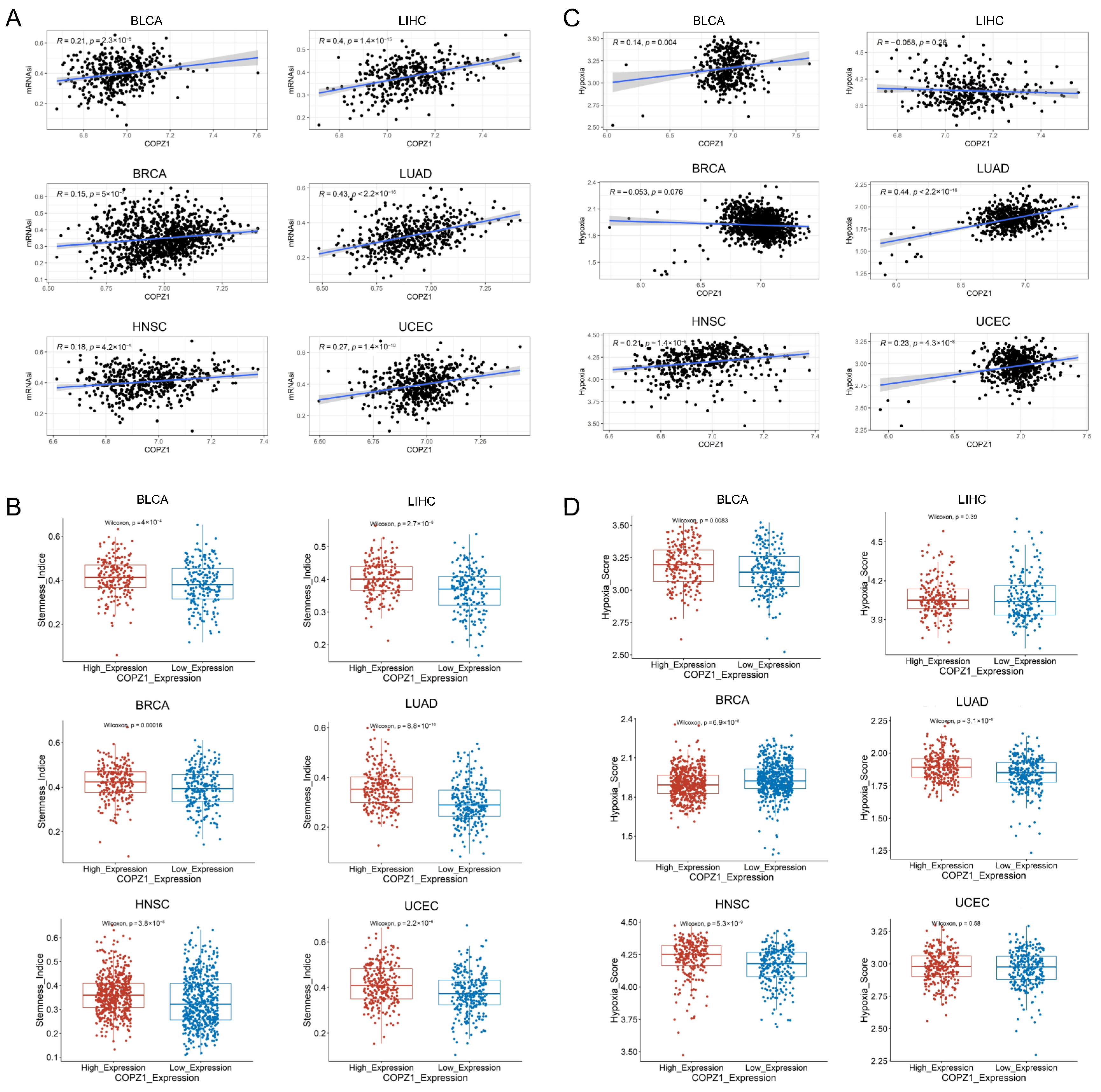
3.4. COPZ1 Positively Correlated with Stemness Score and Hypoxia Score
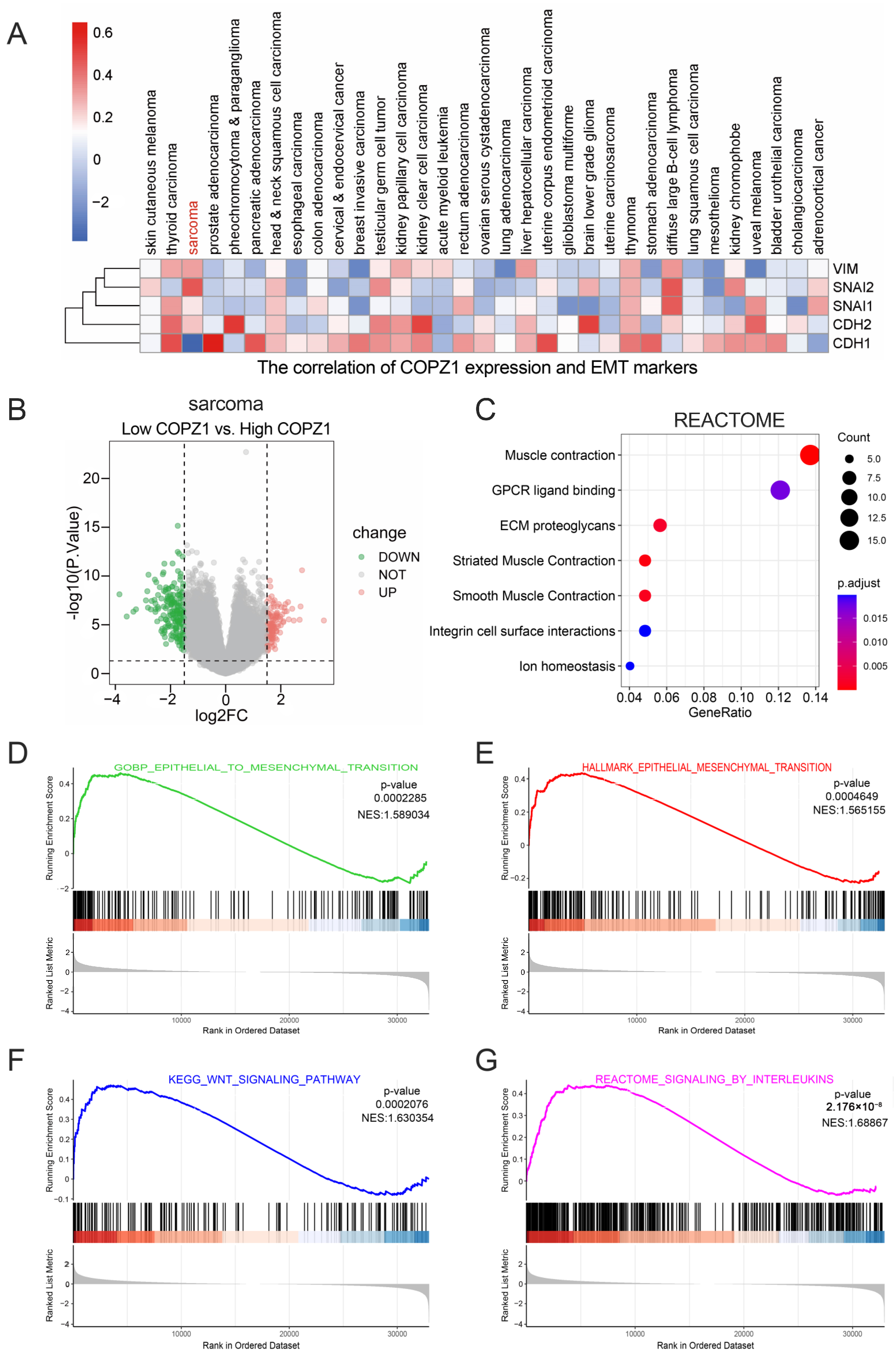
3.5. The Relationship between COPZ1 Expression and Tumor Metastasis
3.6. Correlation Analysis of COPZ1 Expression with Tumor Mutation Burden and Microsatellite Instability
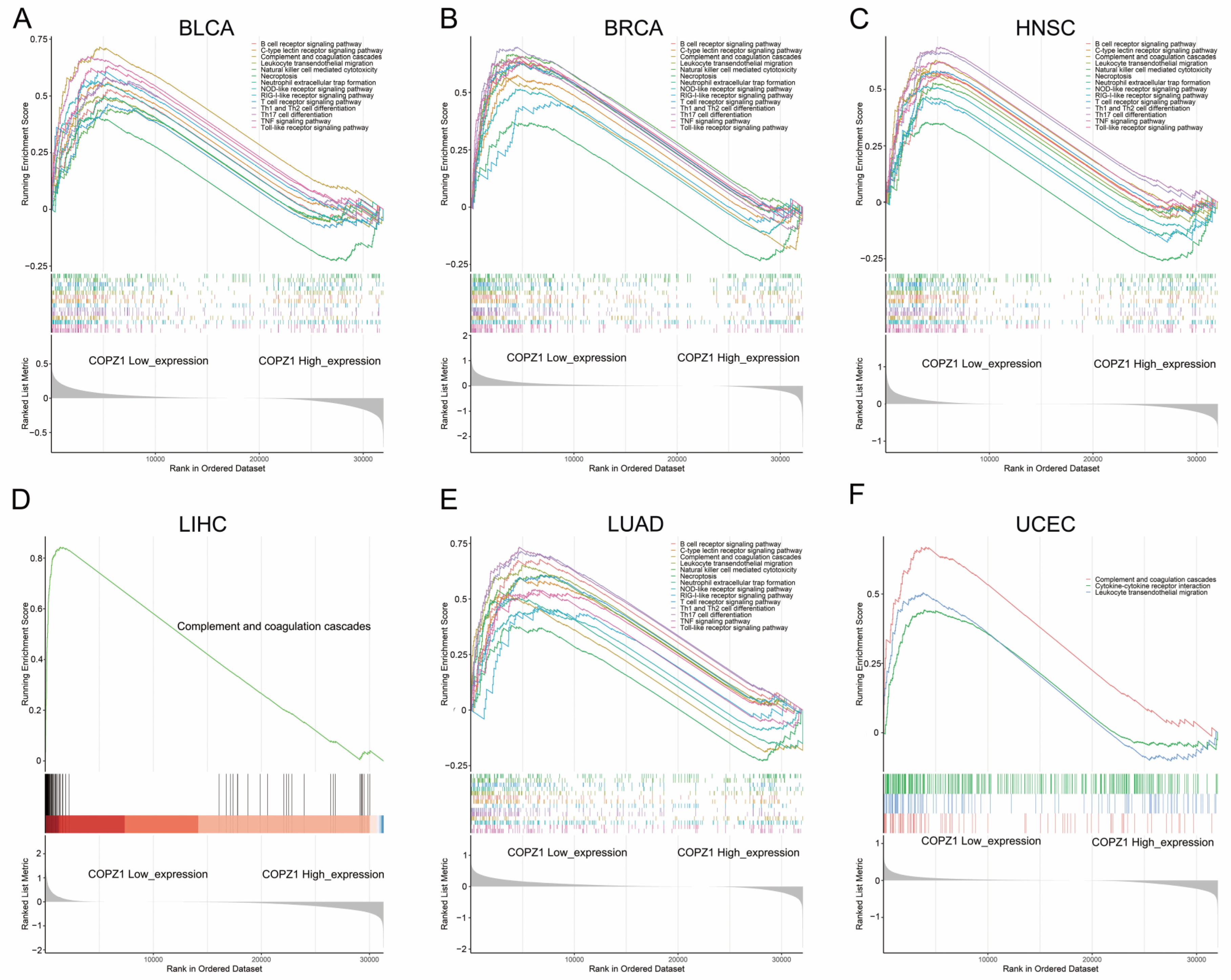
3.7. Pathway Enrichment Based on the Expression of COPZ1
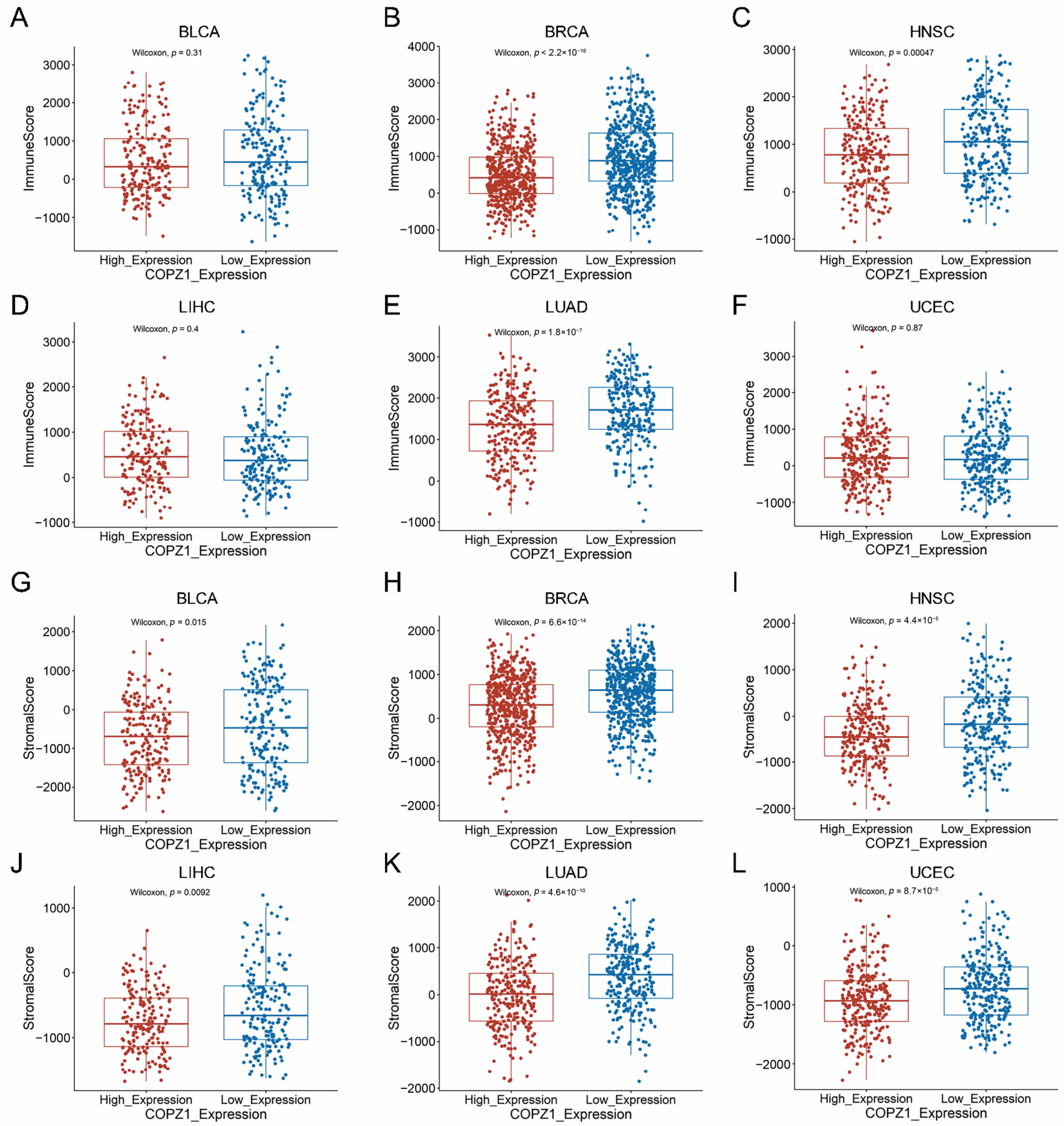
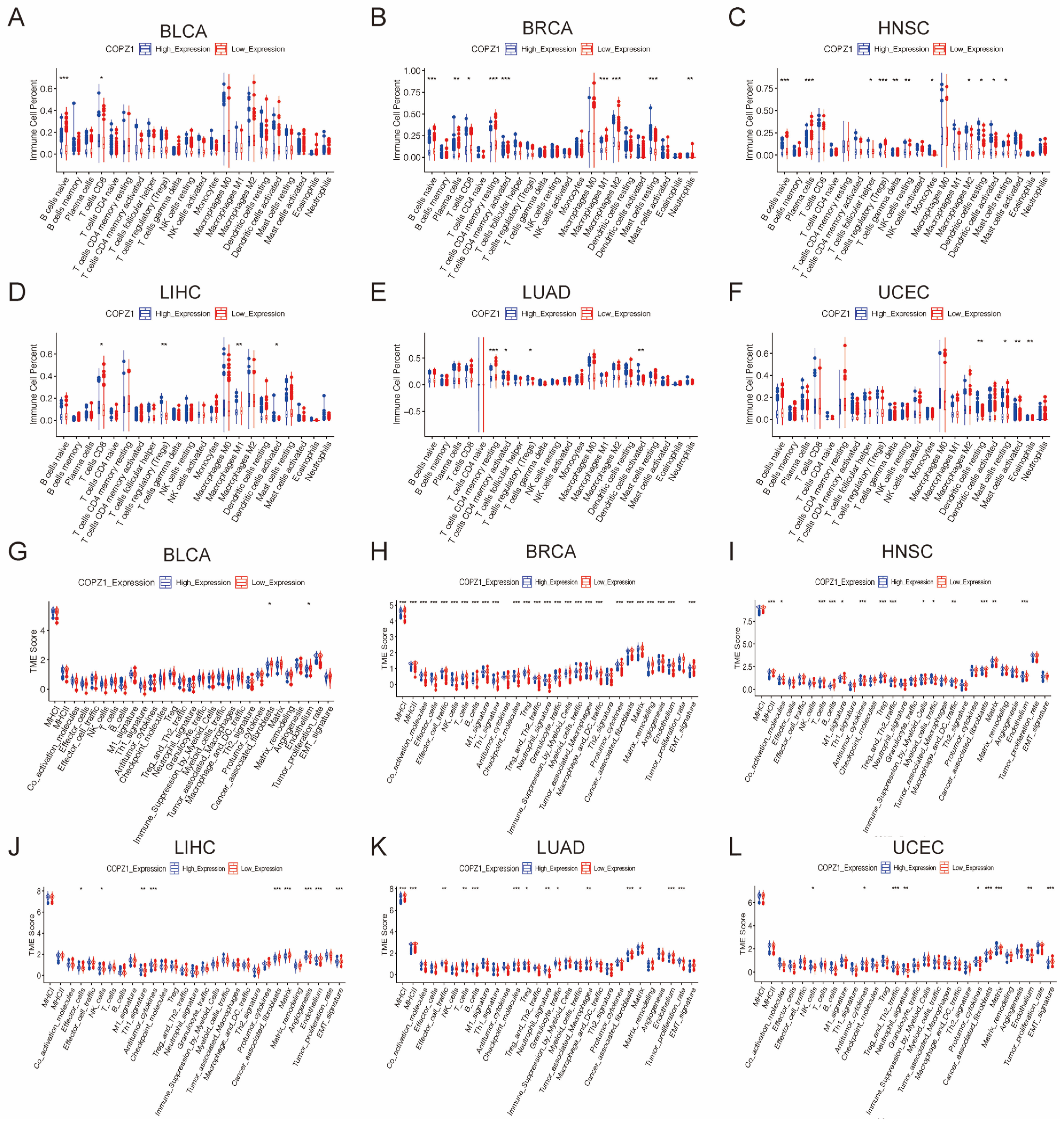
3.8. Correlation Analysis of COPZ1 Expression with Immune Cells and Tumor Microenvironment
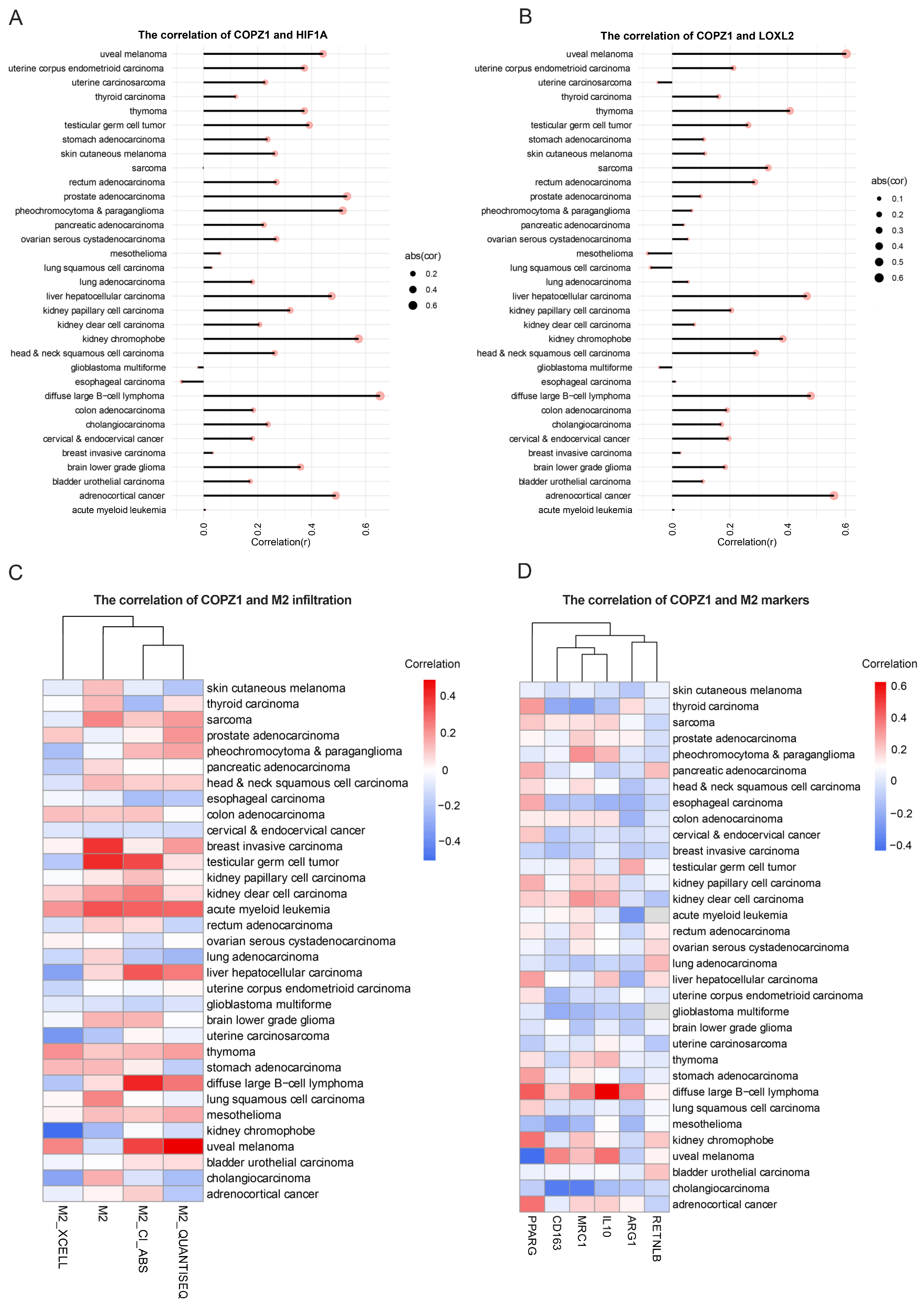
3.9. Correlation Analysis of COPZ1 Expression with Hypoxia and M2 Infiltration Level
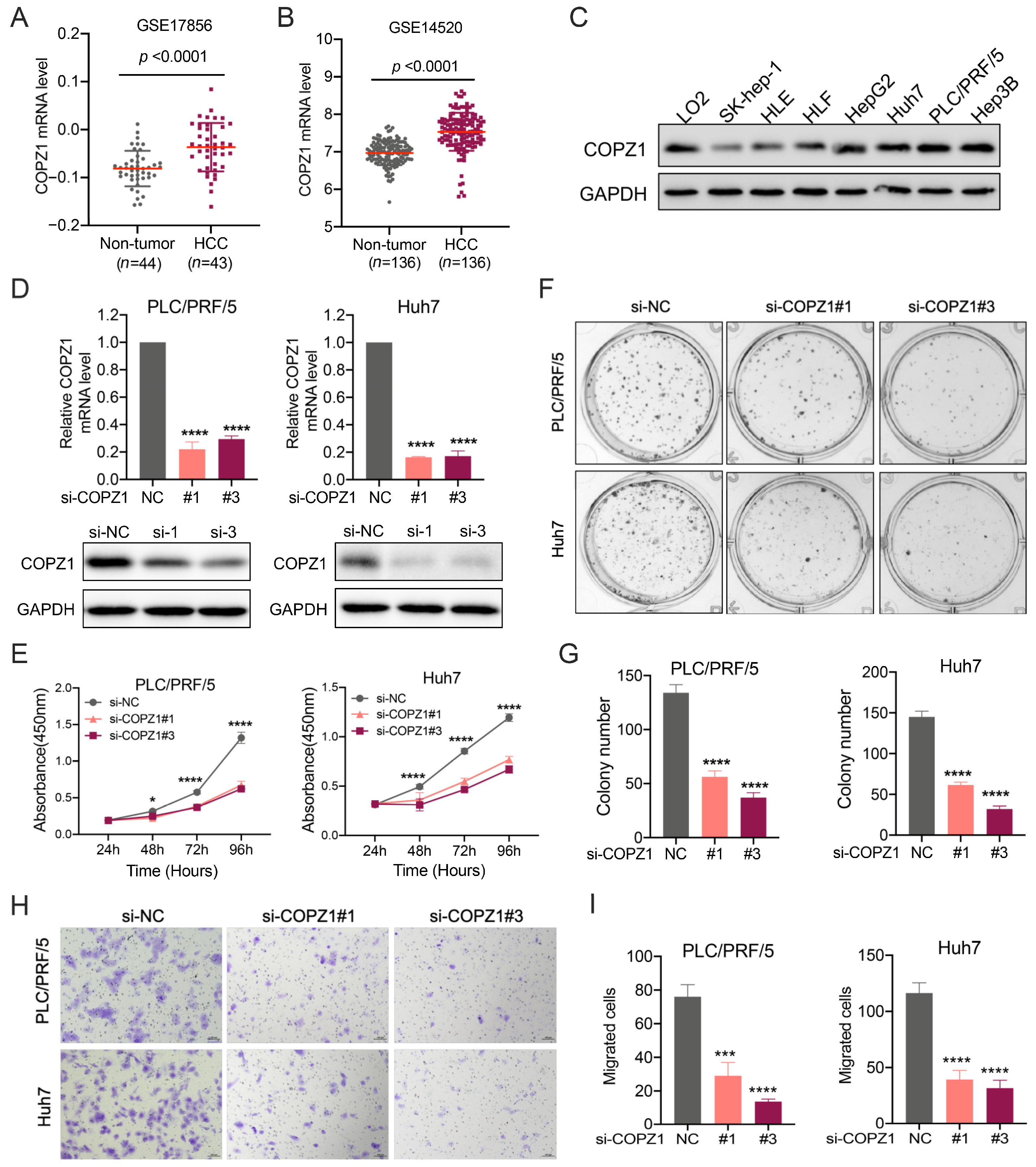
3.10. High COPZ1 Levels Contribute to the Proliferation and Migration of HCC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Torry, D.S.; Cooper, G.M. Proto-Oncogenes in Development and Cancer. Am. J. Reprod. Immunol. 1991, 25, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-H.; Wu, C.-F.; Rajasekaran, N.; Shin, Y.K. Loss of Tumor Suppressor Gene Function in Human Cancer: An Overview. Cell. Physiol. Biochem. 2018, 51, 2647–2693. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.-Y.; Bang, Y.-J. HER2-targeted therapies—A role beyond breast cancer. Nat. Rev. Clin. Oncol. 2019, 17, 33–48. [Google Scholar] [CrossRef]
- Roskoski, R., Jr. Small molecule inhibitors targeting the EGFR/ErbB family of protein-tyrosine kinases in human cancers. Pharmacol. Res. 2019, 139, 395–411. [Google Scholar] [CrossRef]
- Davies, H.; Bignell, G.R.; Cox, C.; Stephens, P.; Edkins, S.; Clegg, S.; Teague, J.; Woffendin, H.; Garnett, M.J.; Bottomley, W.; et al. Mutations of the BRAF gene in human cancer. Nature 2002, 417, 949–954. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Liu, D.; Li, L. PD-1/PD-L1 pathway: Current researches in cancer. Am. J. Cancer Res. 2020, 10, 727–742. [Google Scholar]
- Beck, R.; Rawet, M.; Wieland, F.T.; Cassel, D. The COPI system: Molecular mechanisms and function. FEBS Lett. 2009, 583, 2701–2709. [Google Scholar] [CrossRef] [Green Version]
- Beller, M.; Sztalryd, C.; Southall, N.; Bell, M.; Jäckle, H.; Auld, D.S.; Oliver, B. COPI Complex Is a Regulator of Lipid Homeostasis. PLoS Biol. 2008, 6, e292. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Luo, X.; Zhao, S.; Siu, G.K.; Liang, Y.; Chan, H.C.; Satoh, A.; Yu, S.S. COPI-TRAPPII activates Rab18 and regulates its lipid droplet association. EMBO J. 2017, 36, 441–457. [Google Scholar] [CrossRef] [Green Version]
- Razi, M.; Chan, E.Y.; Tooze, S.A. Early endosomes and endosomal coatomer are required for autophagy. J. Cell Biol. 2009, 185, 305–321. [Google Scholar] [CrossRef] [Green Version]
- Shtutman, M.; Baig, M.; Levina, E.; Roninson, I.B. Tumor-specific silencing of COPZ2 gene encoding coatomer protein complex subunit zeta 2 renders tumor cells dependent on its paralogous gene COPZ1. Proc. Natl. Acad. Sci. USA 2011, 108, 12449–12454. [Google Scholar] [CrossRef] [Green Version]
- Anania, M.C.; Cetti, E.; Lecis, D.; Todoerti, K.; Gulino, A.; Mauro, G.; DI Marco, T.; Cleris, L.; Pagliardini, S.; Manenti, G.; et al. Targeting COPZ1 non-oncogene addiction counteracts the viability of thyroid tumor cells. Cancer Lett. 2017, 410, 201–211. [Google Scholar] [CrossRef]
- Di Marco, T.; Bianchi, F.; Sfondrini, L.; Todoerti, K.; Bongarzone, I.; Maffioli, E.M.; Tedeschi, G.; Mazzoni, M.; Pagliardini, S.; Pellegrini, S.; et al. COPZ1 depletion in thyroid tumor cells triggers type I IFN response and immunogenic cell death. Cancer Lett. 2020, 476, 106–119. [Google Scholar] [CrossRef]
- Zhang, Y.; Kong, Y.; Ma, Y.; Ni, S.; Wikerholmen, T.; Xi, K.; Zhao, F.; Zhao, Z.; Wang, J.; Huang, B.; et al. Loss of COPZ1 induces NCOA4 mediated autophagy and ferroptosis in glioblastoma cell lines. Oncogene 2021, 40, 1425–1439. [Google Scholar] [CrossRef]
- Li, T.; Fu, J.; Zeng, Z.; Cohen, D.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids Res. 2020, 48, W509–W514. [Google Scholar] [CrossRef]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [Green Version]
- Mohr, A.M.; Mott, J.L. Overview of MicroRNA Biology. Semin. Liver Dis. 2015, 35, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Malta, T.M.; Sokolov, A.; Gentles, A.J.; Burzykowski, T.; Poisson, L.; Weinstein, J.N.; Kamińska, B.; Huelsken, J.; Omberg, L.; Gevaert, O.; et al. Machine Learning Identifies Stemness Features Associated with Oncogenic Dedifferentiation. Cell 2018, 173, 338–354.e15. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Li, H.; Chen, S.; Wang, B.; Zhang, K. BMI1 promotes the proliferation and inhibits autophagy of breast cancer cells by activating COPZ1. Clin. Transl. Oncol. 2022, 24, 2166–2174. [Google Scholar] [CrossRef]
- Sui, B.; Wang, M.; Cheng, C.; Zhang, Q.; Zhang, J.; Fan, D.; Xu, P. Nanogel-Facilitated Protein Intracellular Specific Degradation through Trim-Away. Adv. Funct. Mater. 2021, 31, 556. [Google Scholar] [CrossRef]
- Islam, F.; Gopalan, V.; Smith, R.A.; Lam, A.K. Translational potential of cancer stem cells: A review of the detection of cancer stem cells and their roles in cancer re-currence and cancer treatment. Exp. Cell Res. 2015, 335, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Suresh, R.; Ali, S.; Ahmad, A.; Philip, P.A.; Sarkar, F.H. The Role of Cancer Stem Cells in Recurrent and Drug-Resistant Lung Cancer. In Lung Cancer and Personalized Medicine: Novel Therapies and Clinical Management; Springer: Cham, Switzerland, 2015; Volume 890, pp. 57–74. [Google Scholar] [CrossRef]
- Sokolov, A.; Paull, E.O.; Stuart, J.M. One-Class Detection of Cell States in Tumor Subtypes. Biocomputing 2015, 21, 405–416. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, Y.; Yang, T.; Xing, H.; Wang, Y.; Gao, L.; Guo, X.; Xing, B.; Wang, Y.; Ma, W. Machine learning revealed stemness features and a novel stemness-based classification with appealing implications in discriminating the prognosis, immunotherapy and temozolomide responses of 906 glioblastoma patients. Briefings Bioinform. 2021, 22, bbab032. [Google Scholar] [CrossRef] [PubMed]
- Ben-Porath, I.; Thomson, M.W.; Carey, V.J.; Ge, R.; Bell, G.W.; Regev, A.; Weinberg, R.A. An embryonic stem cell–like gene expression signature in poorly differentiated aggressive human tumors. Nat. Genet. 2008, 40, 499–507. [Google Scholar] [CrossRef]
- Shi, X.; Liu, Y.; Cheng, S.; Hu, H.; Zhang, J.; Wei, M.; Zhao, L.; Xin, S. Cancer Stemness Associated with Prognosis and the Efficacy of Immunotherapy in Adrenocortical Carcinoma. Front. Oncol. 2021, 11, 651622. [Google Scholar] [CrossRef]
- Li, N.; Li, Y.; Zheng, P.; Zhan, X. Cancer Stemness-Based Prognostic Immune-Related Gene Signatures in Lung Adenocarcinoma and Lung Squamous Cell Carcinoma. Front. Endocrinol. 2021, 12, 755805. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, X.; Chen, X.; Guo, F.; Hong, J. Prognostic Value of a Stemness Index-Associated Signature in Primary Lower-Grade Glioma. Front. Genet. 2020, 11, 441. [Google Scholar] [CrossRef]
- Jain, R.K. Antiangiogenesis Strategies Revisited: From Starving Tumors to Alleviating Hypoxia. Cancer Cell 2014, 26, 605–622. [Google Scholar] [CrossRef] [Green Version]
- Shao, C.; Yang, F.; Miao, S.; Liu, W.; Wang, C.; Shu, Y.; Shen, H. Role of hypoxia-induced exosomes in tumor biology. Mol. Cancer 2018, 17, 120. [Google Scholar] [CrossRef]
- Nelson, D.A.; Tan, T.-T.; Rabson, A.B.; Anderson, D.; Degenhardt, K.; White, E. Hypoxia and defective apoptosis drive genomic instability and tumorigenesis. Genes Dev. 2004, 18, 2095–2107. [Google Scholar] [CrossRef] [Green Version]
- Gilkes, D.M.; Semenza, G.L.; Wirtz, D. Hypoxia and the extracellular matrix: Drivers of tumour metastasis. Nat. Rev. Cancer 2014, 14, 430–439. [Google Scholar] [CrossRef] [Green Version]
- Barsoum, I.B.; Smallwood, C.A.; Siemens, D.R.; Graham, C.H. A Mechanism of Hypoxia-Mediated Escape from Adaptive Immunity in Cancer Cells. Cancer Res. 2014, 74, 665–674. [Google Scholar] [CrossRef] [Green Version]
- Mo, Z.; Liu, D.; Rong, D.; Zhang, S. Hypoxic Characteristic in the Immunosuppressive Microenvironment of Hepatocellular Carcinoma. Front. Immunol. 2021, 12, 611058. [Google Scholar] [CrossRef]
- Liu, Z.; Tang, Q.; Qi, T.; Othmane, B.; Yang, Z.; Chen, J.; Hu, J.; Zu, X. A Robust Hypoxia Risk Score Predicts the Clinical Outcomes and Tumor Microenvironment Immune Characters in Bladder Cancer. Front. Immunol. 2021, 12, 725223. [Google Scholar] [CrossRef]
- Luo, F.; Lu, F.-T.; Cao, J.-X.; Ma, W.-J.; Xia, Z.-F.; Zhan, J.-H.; Zeng, K.-M.; Huang, Y.; Zhao, H.-Y.; Zhang, L. HIF-1α inhibition promotes the efficacy of immune checkpoint blockade in the treatment of non-small cell lung cancer. Cancer Lett. 2022, 531, 39–56. [Google Scholar] [CrossRef]
- Park, J.E.; Dutta, B.; Tse, S.W.; Gupta, N.; Tan, C.F.; Low, J.K.; Yeoh, K.W.; Kon, O.L.; Tam, J.P.; Sze, S.K. Hypoxia-induced tumor exosomes promote M2-like macrophage polarization of infiltrating myeloid cells and mi-croRNA-mediated metabolic shift. Oncogene 2019, 38, 5158–5173. [Google Scholar] [CrossRef]
- Corzo, C.A.; Condamine, T.; Lu, L.; Cotter, M.J.; Youn, J.-I.; Cheng, P.; Cho, H.-I.; Celis, E.; Quiceno, D.G.; Padhya, T.; et al. HIF-1α regulates function and differentiation of myeloid-derived suppressor cells in the tumor microenvironment. J. Exp. Med. 2010, 207, 2439–2453. [Google Scholar] [CrossRef]
- Noman, M.Z.; Desantis, G.; Janji, B.; Hasmim, M.; Karray, S.; Dessen, P.; Bronte, V.; Chouaib, S. PD-L1 is a novel direct target of HIF-1α, and its blockade under hypoxia enhanced MDSC-mediated T cell activation. J. Exp. Med. 2014, 211, 781–790. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Bagchi, S.; Yuan, R.; Engleman, E.G. Immune Checkpoint Inhibitors for the Treatment of Cancer: Clinical Impact and Mechanisms of Response and Resistance. Annu. Rev. Pathol. Mech. Dis. 2021, 16, 223–249. [Google Scholar] [CrossRef] [PubMed]
- Passaro, A.; Brahmer, J.; Antonia, S.; Mok, T.; Peters, S. Managing Resistance to Immune Checkpoint Inhibitors in Lung Cancer: Treatment and Novel Strategies. J. Clin. Oncol. 2022, 40, 598–610. [Google Scholar] [CrossRef] [PubMed]
- Petitprez, F.; Meylan, M.; De Reyniès, A.; Sautes-Fridman, C.; Fridman, W.H. The Tumor Microenvironment in the Response to Immune Checkpoint Blockade Therapies. Front. Immunol. 2020, 11, 784. [Google Scholar] [CrossRef]
- Tong, Y.; Peng, M.; Li, J.; Niu, Y. Comprehensive Analyses of Stromal-Immune Score-Related Competing Endogenous RNA Networks in Colon Adeno-carcinoma. Dis. Markers 2022, 2022, 4235305. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Lu, L.; Liang, X.; Chen, Y. Identification of a pyroptosis-related lncRNA risk model for predicting prognosis and immune response in colon adeno-carcinoma. World J. Surg Oncol. 2022, 20, 118. [Google Scholar] [CrossRef]
- Li, Y.; Patel, S.P.; Roszik, J.; Qin, Y. Hypoxia-Driven Immunosuppressive Metabolites in the Tumor Microenvironment: New Approaches for Combinational Immunotherapy. Front. Immunol. 2018, 9, 1591. [Google Scholar] [CrossRef] [Green Version]
- Multhoff, G.; Vaupel, P. Hypoxia Compromises Anti-Cancer Immune Responses. In Oxygen Transport to Tissue XLI; Springer: Cham, Switzerland, 2020; Volume 1232, pp. 131–143. [Google Scholar] [CrossRef]
- Quezada, S.; Peggs, K.S.; Simpson, T.R.; Shen, Y.; Littman, D.R.; Allison, J.P. Limited tumor infiltration by activated T effector cells restricts the therapeutic activity of regulatory T cell depletion against established melanoma. J. Exp. Med. 2008, 205, 2125–2138. [Google Scholar] [CrossRef]
- Castagnoli, L.; Cancila, V.; Cordoba-Romero, S.L.; Faraci, S.; Talarico, G.; Belmonte, B.; Iorio, M.V.; Milani, M.; Volpari, T.; Chiodoni, C.; et al. WNT signaling modulates PD-L1 expression in the stem cell compartment of triple-negative breast cancer. Oncogene 2019, 38, 4047–4060. [Google Scholar] [CrossRef] [Green Version]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, Y.; Xia, Z.; Sun, Y.; Lan, Y.; Di, T.; Yang, J.; Sun, J.; Qiu, M.; Luo, Q.; Yang, D. A Comprehensive Pan-Cancer Analysis of the Regulation and Prognostic Effect of Coat Complex Subunit Zeta 1. Genes 2023, 14, 889. https://doi.org/10.3390/genes14040889
Hong Y, Xia Z, Sun Y, Lan Y, Di T, Yang J, Sun J, Qiu M, Luo Q, Yang D. A Comprehensive Pan-Cancer Analysis of the Regulation and Prognostic Effect of Coat Complex Subunit Zeta 1. Genes. 2023; 14(4):889. https://doi.org/10.3390/genes14040889
Chicago/Turabian StyleHong, Ye, Zengfei Xia, Yuting Sun, Yingxia Lan, Tian Di, Jing Yang, Jian Sun, Miaozhen Qiu, Qiuyun Luo, and Dajun Yang. 2023. "A Comprehensive Pan-Cancer Analysis of the Regulation and Prognostic Effect of Coat Complex Subunit Zeta 1" Genes 14, no. 4: 889. https://doi.org/10.3390/genes14040889